Abstract
During the host location process, egg parasitoids can eavesdrop on chemical cues released from immature and adult hosts. These indirect host-related cues are highly detectable, but of low reliability because they lead egg parasitoid females to an area where oviposition is likely to occur rather then providing wasps with direct information on the presence of eggs and their location. In the host-parasitoid associations between true bugs and their scelionid egg parasitoids, female wasps perceive the chemical residues left by host adults walking on substrates as contact kairomones, displaying a characteristic arrestment posture. In this study, we demonstrated that epicuticular waxes of leaves of two vegetable crops, broad bean, Vicia faba, and collard greens, Brassica oleracea, mediate the foraging behavior of Trissolcus basalis (Wollaston) by adsorbing contact kairomones from adults of Nezara viridula (L.). Trissolcus basalis females showed no response when released on the adaxial leaf surface of broad bean or collard green plants with intact cuticular wax layers that had not been exposed to bugs, whereas wasps displayed the arrestment posture when intact leaves were contaminated by chemical residues from host females. Adaxial leaf surfaces that were dewaxed with an aqueous solution of gum arabic and afterwards contaminated by N. viridula females elicited no arrestment responses from wasp females. Similarly, leaves contaminated by host females and subsequently dewaxed did not elicit responses from female wasps. These findings reveal the important role of plant waxes in N. viridula - T. basalis semiochemical communication.
Insect parasitoids find their hosts via a series of behavioral steps which are mainly mediated by chemical cues directly or indirectly related to the host.Citation1 It is generally accepted that egg parasitoids eavesdrop on indirect host related cues to reach areas where the probability of finding target host eggs is high, allowing the wasps to locate host eggs quickly, before they become unsuitable for parasitoid development ().Citation2,Citation3 Chemical residues left behind by immatures and adults of true bugs (Hemiptera: Heteroptera) on a substrate are examples of indirect host-related cues that act as contact kairomones for scelionid wasp parasitoids (). Experiments conducted in filter paper arenas showed that Telenomus and Trissolcus species reacted to chemical footprints of true bugs with an initial prolonged motionless period with the antennae held in contact with the surface (arrestment response), followed by a modified walking pattern characterized by reduced linear speed and increased turning rate (focused searching behavior). Furthermore, wasp flight is inhibited, so that wasps remain in the area where chemicals have been perceived and intensively search the potential host patch. If no host eggs are found, searching behavior slowly decreases and the normal walking pattern is gradually resumed.Citation4–Citation8
Plant cuticles consist of a cutin matrix with intrusions of wax termed the “intracuticular waxes”. The cuticle is overlaid by a second wax layer termed the “epicuticular waxes”, that can occur as minute crystals and/or as thin films. Because the wax layer is the primary interface between plants and insects, the epicuticular layer can influence the behavior of phytophagous insects, and their parasitoids and predators.Citation9–Citation11 Previous studies have shown that plant surfaces adsorb and release volatile/contact chemicals which mediate the foraging behaviour of parasitoid wasps.Citation12,Citation13 The role played by the plant surface in the effectiveness of entomophagous insects has been investigated mostly by using mutant plants with reduced leaf waxes.Citation14–Citation17 Recently, a wide range of adhesive methods were tested for removal of the outermost leaf wax layer without damaging the plant cutin, and keeping the intracuticular and epicuticular material separated.Citation18,Citation19 In a recent publication, we reported the removal of epicuticular waxes using gum arabic as an adhesive, in order to investigate the adsorption or dilution of insect semiochemicals by plant waxes.Citation20 To verify whether leaf cuticle can influence the foraging behaviour of the egg parasitoid Trissolcus basalis (Hymenoptera: Scelionidae), a natural enemy of Nezara viridula (Heteroptera: Pentatomidae), experiments were conducted on broad bean (BB), Vicia faba (L.), and on collard greens (CG), Brassica oleracea (L.). Leaves of BB and CG were tested under the following sets of conditions: (1) with intact epicuticular wax layer, or with the outermost wax layer mechanically removed; (2) either uncontaminated or contaminated by N. viridula footprints. Epicuticular waxes were stripped from the adaxial leaf surface by applying a 50% (w/w) aqueous solution of gum Arabic (Sigma-Aldrich) (approximately 0.05 g/cm2). After the solution had dried (about two hours), the thin polymer film with the epicuticular wax layer adhering to it, was peeled off with forceps. Dewaxed plants were then transferred into a climate-controlled room for bioassays. Treatments contaminated with bug footprints were prepared by allowing 2–3 mated N. viridula females to walk over the leaves for about 30 min. Then the adults were removed and the plants were transferred into a climate-controlled room for bioassays. In a first set of experiments, uncontaminated BB and CG plants with intact wax layers (1) were compared to used plants with intact wax layers contaminated with residues from host females (2). In a second set of experiments, BB and CG leaves were tested dewaxed without host chemical contamination (3), dewaxed after being contaminated (4), or dewaxed before being contaminated by host female footprints (5). For all experiments, <6-d-old, mated, naïve T. basalis females were used. Wasp behavior was observed manually, and was scored as “positive response” when a female, immediately after being released onto the adaxial leaf surface, displayed the typical arrestment posture, remaining motionless with the antennae held in contact on the plant surface.
In experiments with control leaves, i.e., leaves with intact adaxial wax layer and without host female chemical contamination, T. basalis showed no arrestment response (about 77% of females on BB and about 82% on CG showed no response). Wasps behaved similarly on dewaxed and uncontaminated leaves, suggesting that the mechanical removal of epicuticular waxes per se did not noticeably influence the parasitoids’ behavior. In contrast, wasps’ responses to leaves contaminated with host footprints were significantly affected by the presence/absence of epicuticular waxes on the adaxial leaf surface (). The frequency of females displaying the typical arrestment posture was significantly higher on contaminated plants with intact epicuticular waxes (about 70% of wasps showed positive responses on BB and about 82% on CG). The rate of response decreased in the treatments in which the epicuticular waxes had been removed, both when contamination was carried out prior to wax removal, and when contamination was performed after leaves had been dewaxed. These results suggested that the detection of contact kairomones by T. basalis is due to the adsorption of host footprints onto the epicuticular waxes of V. faba and B. oleracea. The effect of BB and CG epicuticular waxes on semiochemical communication in the system N. viridula and T. basalis resembles the effects recorded in other multitrophic systems that have been investigated.Citation21 Furthermore, N. viridula residues adhering to the wax crystals were transferred, maintaining the kairomonal proprieties, onto the surface of the gum arabic film peeled off the leaves. That is, in laboratory bioassays, the polymer film with adhering waxes stripped from the adaxial leaf surface of BB contaminated by host footprints elicited positive responses from T. basalis females (95% of females tested), whereas no significant responses were observed to films from uncontaminated leaves (15% arrestment).
BB and CG leaves were observed by scanning electron microscopy (S.E.M.) (FEI Quanta 200F, Holland) to examine the effect of the mechanic removal of the epicuticular waxes on the morphological leaf structure. Samples of about 1 cm2 of both leaves were divided into two parts, with one being dewaxed with gum arabic as described above, whereas the other was left untreated. On BB leaves, epicuticular waxes occurred as a film, densely crystallized into irregularly shaped platelets with spherical granules randomly distributed among them (). In CG leaves, epicuticular waxes showed an entirely different morphology, occurring as tubules and rods (). Inspection by S.E.M. showed that in both plants a single treatment with gum arabic removed the epicuticular wax crystals, and this effect was clearly visible along the border line between the dewaxed and untreated area of each leaf sample. This removal apparently was sufficient to prevent the adsorption of the chemical footprints left by N. viridula females, resulting in the reduced responses by T. basalis.
What remains unknown is the adsorption mechanism of the footprints by the plant epicuticular waxes. Recent research found that cuticular lipids of N. viridula females contain high molecular weight glycerides, long-chain alcohols, and fatty acids.Citation22 Furthermore, the presence or absence of nonadecane (nC19) a sex-specific component of the cuticular hydrocarbons of male N. viridula, allowed wasp females to distinguish between residues left by male or female bugs, respectively.Citation23 This suggested that wasp females might use both hydrocarbon and more polar components present in host footprints, that can adhere to or be adsorbed by V. faba and B. oleracea epicuticular waxes. However, plant cuticular waxes consist of mixtures of long-chain aliphatic and cyclic compoundsCitation24 which among species, so that the chemical proprieties of wax layers will vary from system to system. Therefore, an investigation of the epicuticular wax composition of BB and CG plants in terms of the chemistry and polarity of wax constituents is necessary to better understand the adsorption mechanism of N. viridula footprints. A deeper knowledge of wax chemical classes also could clarify how T. basalis females distinguish the components of the contact kairomones from similar components present in the epicuticular waxes of foliage and other plant tissues, that can potentially create confounding or masking effects.
Figures and Tables
Figure 1 Overview of the chemical cues involved in the host location process in the host-egg parasitoid associations of true bugs and scelinoid wasps. To find host eggs, parasitoids can use semiochemicals originating from the host eggs (direct host-related cues) or indirect host-related cues from adults of the host species.
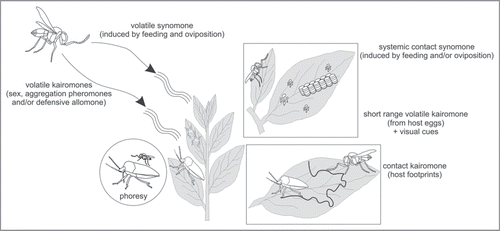
Figure 2 Effects of adaxial leaf epicuticular waxes of Vicia faba and Brassica oleracea on Trissolcus basalis response to Nezara viridula female footprints that constitute a contact kairomone. Green bars indicate the percentage of wasps that displayed arrestment responses after being released; red bars represent the percentage of those that did no show arrestment responses. Leaves were contaminated by allowing 2–3 mated, gravid N. viridula to walk over them for 30 min. Leaves were mechanically dewaxed with an aqueous solution of gum arabic. Asterisks indicate significant differences at p < 0.01 (Pearson χ2 test).
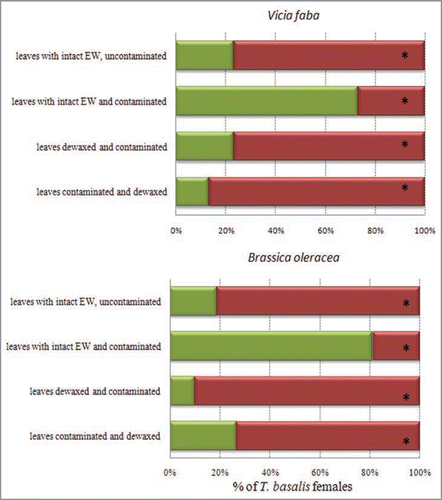
Figure 3 Scanning electron micrographs of the adaxial leaf surface of Vicia faba (A) and Brassica oleracea (B) with the epicuticular waxes partly removed after a single treatment with gum Arabic, revealing the smooth surface of the cutin matrix. The line between the dewaxed area (bottom half of the pictures) from the neighbouring intact area is highlighted by arrows.
Acknowledgements
Funding for this work was provided by “Progetti di ricerca di interesse nazionale—cofinanziamento 2007” entitled “Enhancing foraging behaviour of insect egg parasitoids: the role of volatile organic compounds and the epicuticular layers of plants” Chairperson Prof. Stefano Colazza. Gianandrea Salerno provided the drawing. We thank Jocelyn Millar for critically reading and commenting the manuscript.
Addendum to:
References
- Vinson SB. The general host selection behavior of parasitoid Hymenoptera and a comparison of initial strategies utilized by larvaphagous and oophagous species. Biol Control 1998; 11:79 - 97
- Vet LEM, Dicke M. Ecology of infochemical use by natural enemies in a tritrophic context. Annu Rev Entomol 1992; 37:141 - 172
- Godfray HCJ. Parasitoids. Behavioral and Evolutionary Ecology 1994; Princeton, New Jersey Princeton University Press,
- Colazza S, Salerno G, Wajnberg E. Volatile and contact chemicals released by Nezara viridula (Heteroptera: Pentatomidae) have a kairomonal effect on the egg parasitoid Trissolcus basalis (Hymenoptera: Scelionidae). Biol Control 1999; 16:310 - 317
- Borges M, Colazza S, Ramirez-Lucas P, Chauhan KR, Moraes MCB, Aldrich JR. Kairomonal effect of walking traces from Euschistus heros (Heteroptera: Pentatomidae) on two strains of Telenomus podisi (Hymenoptera: Scelionidae). Physiolog Entomol 2003; 28:349 - 355
- Conti E, Salerno G, Bin F, Vinson SB. The role of host semiochemicals in parasitoid specificity: a case study with Trissolcus brochymenae and Trissolcus simoni on pentatomid bugs. Biol Control 2004; 29:435 - 444
- Salerno G, Conti E, Peri E, Colazza S, Bin F. Kairomone involvement in the host specificity of the egg parasitoid Trissolcus basalis (Hymenoptera: Scelionidae). Eur J Entomol 2006; 103:311 - 318
- Peri E, Sole MA, Wajnberg E, Colazza S. Effect of host kairomones and oviposition experience on the arrestment behaviour of an egg parasitoid. J Exp Biol 2006; 209:3629 - 3635
- Espelie KE, Bernays EA, Brown JJ. Plant and insect cuticular lipids serve as behavioral cues for insects. Arch Insect Biochem Physiol 1991; 17:223 - 233
- McAuslane HJ, Simmons AM, Jackson DM. Parasitism of Bemisia argentifolii on collard with reduced or normal leaf wax. Flo Entomol 2000; 83:428 - 437
- Brennan EB, Weinbaum SA. Stylet penetration and survival of three psyllid species on adult leaves and ‘waxy’ and ‘de-waxed’ juvenile leaves of Eucalyptus globulus. Entomolog Experimental Applicata 2001; 100:355 - 363
- Noldus LPJJ, Potting RPJ, Barendregt HE. Moth sex-pheromone adsorption to leaf surface—bridge in time for chemical spies. Physiol Entomol 1991; 16:329 - 344
- Müller C, Riederer M. Plant surface properties in chemical ecology. J Chem Ecol 2005; 31:2621 - 2651
- White C, Eigenbrode SD. Effects of surface wax variation in Pisum sativum L. on herbivorous and entomophagous insects in the field. Environ Entomol 2000; 29:776 - 780
- Chang G, Neufeld J, Durr D, Duetting PS, Eigenbrode SD. Waxy bloom in peas influences the performance and behavior of Aphidius ervi, a parasitoid of the Pea Aphid. Entomol Exp et Appl 2004; 110:257 - 265
- Eigenbrode SD. The effects of plant epicuticular waxy blooms on attachment and effectiveness of predatory insects. Arthropod Struct Dev 2004; 33:91 - 102
- Rutledge C, Eigenbrode SD, Ding HJ. A plant surface mutation mediates predator interference among Ladybird larvae. Ecolog Entomol 2008; 33:464 - 472
- Neinhuis C, Barthlott W, Ensikat H. Direct access to plant epicuticular wax crystals by a new mechanical isolation method. Internat J Plant Sci 2000; 161:143 - 148
- Schaffer S, Riederer M, Jetter R. Leaf cuticular waxes are arranged in chemically and mechanically distinct layers: evidence from Prunus laurocerasus L. Plant, Cell Environ 2000; 23:619 - 628
- Colazza S, Lo Bue M, Lo Giudice D, Peri E. The response of Trissolcus basalis to footprint contact kairomones from Nezara viridula females is mediated by leaf epicuticular waxes. Naturwissenschaften 2009; 96:975 - 981
- Rostás M, Ruf D, Zabka V, Hildebrandt U. Plant surface wax affects parasitoid’s response to host footprints. Naturwissenschaften 2008; 95:997 - 1002
- Lo Giudice D. Contact chemo-orientation in insect parasitoids: the case study of mate-finding in Metaphycus luteolus and host-finding in Trissolcus basalis 2009; Palermo University of Palermo,
- Colazza S, Aquila G, De Pasquale C, Peri E, Millar J. The egg parasitoid Trissolcus basalis uses n-nonadecane, a cuticular hydrocarbon from its stink bug host Nezara viridula, to discriminate between female and male hosts. J Chem Ecol 2007; 33:1405 - 1420
- Koch K, Ensikat HJ. The hydrophobic coatings of plant surfaces: Epicuticular wax crystals and their morphologies, crystallinity and molecular self-assembly. Micron 2008; 39:759 - 772