Abstract
The preprophase band (PPB) of microtubules (MTs) marks the site of the future division plane irrespective of the orientation of the equatorial plane. Because the PPB MTs disappear during prometaphase, some positional information is thought to remain in the cortical cytoplasm after the disappearance of the PPB MTs. Cytoskeletal proteins are known to be excluded from the PPB site during mitosis. These depleted zones of cytoskeletal proteins are potential candidates for a “negative memory” system. However, how these depleted zones of the cytoskeletal proteins are produced remains unknown. In a recent paper, we have quantified the distribution of clathrin-coated pits and vesicles as well as of secretory structures during PPB formation using a combination of high-pressure freezing and electron tomography techniques. Our results demonstrated that the rate of endocytosis is enhanced in PPB regions. We postulate that the removal of membrane proteins by endocytosis plays a role in the creation of PPB "memory" structures.
The preprophase band (PPB) of microtubules (MTs) delineates the future site of cell plate fusion with the mother cell plasma membrane and it has been postulated to be involved in the determination of the division site.Citation1,Citation2,Citation3 The PPB originates during the G2-phase as a broad band of MTs that underlies the plasma membrane and narrows to reach its most compact, mature state during late prophase.Citation4 Because the cell plate fuses with the plasma membrane at the site defined by the narrow PPB, it has been postulated that the PPB leaves behind information or “memory” at or in the plasma membrane.Citation5 Nevertheless, how the PPB marks the future site of cell division is still unknown and has been the subject of many studies and discussions since its discovery in 1966.Citation1,Citation2,Citation3,Citation6
Current Background Information that Supports the PPB Memory Concept
Recently, several potential candidates of a “positive memory” system have been identified. For example, the tangled (tan) gene product and RanGAP1 both accumulate in the PPB and remain there after the disappearance of the PPB MTs.Citation7,Citation8 However, in addition to these “positive memory” structures, PPBs also give rise to “negative memory” structures. Thus, after the narrowing of the PPB MTs, cortical actinCitation9,Citation10 and the kinesin-like molecule, KCA1Citation11 both become depleted in the PPB zone, and these actin and the KCA1 depleted zones persist long after PPB MT breakdown. Yet to be determined is how these “negative memory” structures are established, and how they help define the future site of cell plate fusion with the plasma membrane.
The discovery by early electron microscopists of electron-dense vesicles in the cytoplasm underlying the PPB region in chemically fixed cells led many years ago to the hypothesis that the creation of the memory site could involve deposition of memory-forming molecules by secretion as postulated by the positive memory hypothesis.Citation12,Citation13,Citation14,Citation15 However, the presence of similar vesicles underlying non-PPB regions raised early doubts about the involvement of secretion in PPB formation. Citation15,Citation16 More recently, Dixit and Cyr (2002)Citation6 showed that Golgi secretion is not required for marking the PPB site.
Electron Tomographic Analysis of High Pressure Frozen Cells Demonstrates that PPB Memory Formation Involves Enhanced Endocytic Activity
Cryo-fixation preserves transient membrane systems much better than chemical fixation, and when employed in conjunction with electron tomography it is possible to obtain quantitative information both on the types and the distribution of vesicles in large volumes of cytoplasm in defined cellular domains.Citation17 In earlier ultrastructural studies of chemically fixed cells, the presence of “coated vesicles” was noted in the vicinity of PPBs of some cell types, but neither the nature of these vesicles nor their distribution were systematically analyzed.Citation15,Citation16 In our tomographic study of cryofixed/freeze-substituted cells, these vesicles were identified as being clathrin-coated and to be formed by clathrin-coated endocytic pits. Quantitative analysis of the tomograms indicated, furthermore, that the number of clathrin-coated vesicles in the cortical cytoplasm underlying the PPB regions increased compared to the adjacent, non-PPB regions or the cortical cytoplasm of interphase cells, whereas no differences in secretory structures were seen.Citation18 These observations led to the idea that endocytosis could be involved in the establishment and the maintenance of the division site.
The discovery that PPB formation involves increased rates of endocytosis at the PPB site leads to the question as to what types of plasma membrane molecules could be selectively retrieved from this site by means of the clathrin-coated vesicles. If molecules, that are necessary for the attachment of actin filaments or KCA1 molecules to the plasma membrane were selectively removed by endocytosis, then this could lead to the formation of actin or KCA1 depleted zones. One class of candidate proteins might be the plasma membrane-associated, actin filament-nucleating proteins called formin homology (FH) proteins.Citation19,Citation20 Several plant formins have been shown to have the ability to nucleate actin filaments, and overexpression of AtFH1 induces the formation of arrays of actin cables that project into the cytoplasm from the plasma membrane. Citation21 Thus, one possible function of the enhanced endocytic activity at forming PPBs might be the retrieval of actin-nucleating/binding proteins from these plasma membrane domains to create an actin-free zone to which the expanding cell plate is guided and where it can fuse (). A similar function for the removal of KCA1 can also be envisaged. Together, our data suggest a mechanism for how a “negative memory” structure could be created by PPBs.
Evidence for enhanced rates of endocytosis confined to PPB regions has also been obtained in studies of the uptake of the dye FM4-64 by tobacco BY-2 cells.Citation22 However, in our study, both the tomographic data and immunofluorescent microscopy with anti-clathrin antibodies clearly showed that the density of clathrin-related structures does not decrease abruptly at the edge of the PPB region but decreases gradually. Thus, our tomographic models demonstrate that a significant percentage of the clathrin-bearing structures (clathrin-coated pits and vesicles) are formed adjacent to, but outside the MT band. Based on this observation we postulate that the formation of clathrin-coated pits and vesicles is not tightly coupled to PPB MTs. Instead, the distribution of MTs and the endocytic vesicles in the PPB can be better explained by the formation of some kind of gradient in the PPB region that stimulates the independent assembly of MTs and endocytic vesicles. In this context, the function of the PPB MT array might be both to create a planar reference structure and an associated membrane domain in which the molecules involved in defining the division site can become organized. Therefore, the PPB region is a localizing center of not only MTs but also clathrin-mediated endocytic activity.
Figures and Tables
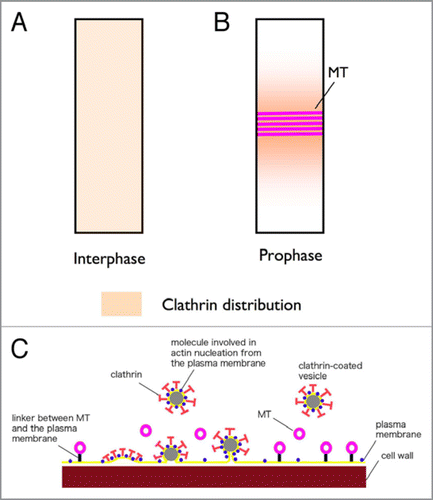
Figure 1 Schematic diagrams showing the formation of clathrin-coated pits and vesicles in the region of PPB MT s. (A and B) Longitudinal views of an interphase (A) and a prophase (B) epidermal cell, and the differences in distribution of cortical clathrin molecules inI such cells. Clathrin molecules are distributed evenly over the surface of interphase cells (A), but concentrated around the PPBs of prophase cells (B). (C) A magnified cross sectional view of the PPB. This model postulates that the function of the endocytic activity at the PPB might be the removal of actin-nucleating/binding proteins from these plasma membrane domains, to create the characteristic actin filament-depleted regions of PPBs.
Acknowledgments
This work was supported by the visiting research associate program of MEXT to IK, JSPS grant 11740454 and 14740454 to IK, JSPS grant 12640651 and 17207006, and MEXT grant 17049019 to YM, NIH grant GM61306 to LAS and Japan-US Cooperative Science Program to Y.M. and L.A.S.
Addendum to:
References
- Pickett-Heaps JD, Northcote DH. Organization of microtubules and endoplasmic reticulum during mitosis and cytokinesis in wheat meristems. J Cell Sci 1966; 1:109 - 120
- Pickett-Heaps JD, Northcote DH. Cell division in the formation of the stomatal complex of the young leaves of wheat. J Cell Sci 1966; 1:121 - 128
- Mineyuki Y. The preprophase band of microtubules: its function as a cytokinetic apparatus in higher plants. Int Rev Cytol 1999; 187:1 - 49
- Mineyuki Y, Wick SM, Gunning BES. Preprophase bands of microtubules and the cell cycle: kinetics and experimental uncoupling of their formation from the nuclear cycle in onion root-tip cells. Planta 1988; 174:518 - 526
- Mineyuki Y, Gunning BES. A role for preprophase bands of microtubules in maturation of new cell walls, and a general proposal on the function of preprophase band sites in cell division in higher plants. J Cell Sci 1990; 97:527 - 537
- Dixit R, Cyr R. Golgi secretion is not required for marking the preprophase band site in cultured tobacco cells. Plant J 2002; 29:99 - 108
- Walker KL, Mueller S, Moss D, Ehrhardt DW, Smith LG. Arabidopsis TANGLED identifies the division plane throughout mitosis and cytokinesis. Curr Biol 2007; 17:1827 - 1836
- Xu XM, Zhao Q, Rodrigo-Peiris T, Brkljacic J, He CS, Muller S, et al. RanGAP1 is a continuous marker of the Arabidopsis cell division plane. Proc Natl Acad Sci 2008; 105:18637 - 18642
- Liu B, Palevitz BA. Organization of cortical microfilaments in dividing root cells. Cell Motil Cytoskel 1992; 23:252 - 264
- Cleary AL, Gunning BES, Wasteneys GO, Hepler PK. Microtubule and F-actin dynamics at the division site in living Tradescantia stamen hair cells. J Cell Sci 1992; 103:977 - 988
- Vanstraelen M, Van Damme D, De Rycke R, Mylle E, Inze D. Cell cycle dependent targeting of a kinesin at the plasma membrane demarcates the division site in plant cells. Curr Biol 2006; 16:308 - 314
- Packard MJ, Stack SM. The preprophase band: possible involvement in the formation of the cell wall. J Cell Sci 1976; 22:403 - 411
- Burgess J, Northcote DH. The relationship between the endoplasmic reticulum and microtubular aggregation and disaggregation. Planta 1968; 80:1 - 14
- Gunning BES, Hughes JE, Hardham AR. Formative and proliferative cell divisions, cell differentiation, and developmental changes in the meristem of Azolla roots. Planta 1978; 143:121 - 144
- Galatis B, Mitrakos K. On the differential divisions and preprophase microtubule bands involved in the development of stomata of Vigna sinensis L. J Cell Sci 1979; 37:11 - 37
- Galatis B, Apostolakos P, Katsaros C, Loukari H. Preprophase microtubule band and local wall thickening in guard cell mother cells of some Leguminosae. Ann Bot 1982; 50:779 - 791
- Donohoe BS, Kang BH, Staehelin LA. Identification and characterization of COPIa- and COPIb-type vesicle classes associated with plant and algal Golgi. Proc Natl Acad Sci 2007; 104:163 - 168
- Karahara I, Suda J, Tahara H, Yokota E, Shimmen T, Misaki K, et al. The preprophase band is a localized center of clathrin-mediated endocytosis in late prophase cells of the onion cotyledon epidermis. Plant J 2009; 57:819 - 831
- Banno H, Chua NH. Characterization of the Arabidopsis formin-like protein AFH1 and its interacting protein. Plant Cell Physiol 2000; 41:617 - 626
- Favery B, Chelysheva LA, Lebris M, Jammes F, Marmagne A, de Almeida-Engler J, et al. Arabidopsis formin AtFH6 is a plasma membrane-associated protein upregulated in giant cells induced by parasitic nematodes. Plant Cell 2004; 16:2529 - 2540
- Cheung AY, Wu H. Overexpression of an Arabidopsis formin stimulates supernumerary actin cable formation from pollen tube cell membrane. Plant Cell 2004; 16:257 - 269
- Dhonukshe P, Mathur J, Hulskamp M, Gadella TWJ. Microtubule plus-ends reveal essential links between intracellular polarization and localized modulation of endocytosis during division-plane establishment in plant cells. BMC Biology 2005; 3:11