Abstract
Recent recordings of the stridulations of Myrmica ants revealed that their queens made distinctive sounds from their workers, although the acoustics of queens and workers, respectively, were the same in different species of Myrmica. Queen recordings induced enhanced protective behavior when played to workers in the one species tested. Larvae and pupae of the butterfly genus Maculinea inhabit Myrmica colonies as social parasites, and both stages generate sounds that mimic those of a Myrmica queen, inducing similar superior treatments from workers as their model. We discuss how initial penetration and acceptance as a colony member is achieved by Maculinea through mimicking the species-specific semio-chemicals of their hosts, and how acoustical mimicry is then employed to elevate the parasite’s membership of that society towards the highest attainable level in their host’s hierarchy. We postulate that, if acoustics is as well developed a means of communication in certain ants as these studies suggest, then others among an estimated 10,000 species of ant social parasite may supplement their well-known use of chemical and tactile mimicry to trick host ants with mimicry of host acoustical systems.
Perhaps 10,000 species of insect have evolved as social parasites to exploit the abundant resources concentrated within dominant ant colonies.Citation1 The adaptations by which infiltration is achieved have been studied in few social parasites,Citation2 but generally involve the corruption, by mimicry, of the honest signals that are continuously transmitted between different members of an ant colony in order to function cohesively as a super-organism.Citation3–Citation5 Beyond the parasitism of one ant colony by another closely-related ant,Citation3 the best understood examples of social manipulation are by Lomechusa (Atemeles) beetles,Citation3 Microdon hoverfliesCitation6 and Maculinea butterflies.Citation2,Citation7 Lomechusa and Maculinea employ physical, behavioural and chemical cues to manipulate ants. We have now demonstrated that the larvae and pupae of Maculinea butterflies are also effective mimics of their hosts’ acoustical communication systems.Citation8–Citation10
Two types of social exploitation have evolved within the genus Maculinea.Citation2,Citation7 All five European species have free-flying adults that oviposit on specific initial foodplant(s). The young larva feeds briefly on growing seeds, developing rapidly through three instars but acquiring <2% of its ultimate biomass.Citation11 On reaching the fourth and final instar, the Maculinea larva (hereafter called caterpillar) falls to the ground and awaits discovery by a foraging Myrmica worker ant, which is tricked by its physical and chemical resemblance to a Myrmica larva into carrying it underground into the brood chambers of its nest.Citation2,Citation12,Citation13 The caterpillar remains within the ant colony for 10–22 monthsCitation14 before pupating, still inside the nest. Those of Maculinea arion and M. teleius have onciform bodies and thick protective cuticles (), and generally live in the outer cells of the nest, making occasional forays to the better protected brood chambers to binge-feed on ant larvae.Citation15 In contrast the ‘cuckoo’ Maculinea, M. rebeli and M. alcon, remain within their host’s brood chambers where they are tended and fed directly by nurse ants on trophic eggs, foraged food and by trophallaxis.Citation13
In most but not all studied populations, each regional phenotype of a Maculinea species is host-specific to a single primary Myrmica host species, whose hydrocarbon profile it mimics by secreting allomones.Citation16,Citation17 In M. alcon, a population-scale evolutionary arms race has been found between the local chemical profiles of the host Myrmica species and its mimetic parasite.Citation18 Although chemical mimicry appears to be amplified in cuckoo Maculinea due to their closer integration with hosts, on current knowledge this cue is used by all Maculinea species to gain acceptance as a member of a host’s society. This, by definition, renders individuals less likely to survive with other Myrmica species whose chemical profiles they imperfectly match.Citation2
While social acceptance is achieved through semio-chemical communication, the caterpillars of cuckoo Maculinea induce nurse ants to feed them by adopting a similar begging behaviour to that of a Myrmica larvaCitation13 (). However, it was evident that caterpillars were often treated as superior individuals within the hierarchy of a Myrmica society, a phenomenon that could not be explained by chemical mimicry.Citation16 Thus when a nest is disturbed, workers rescue M. rebeli caterpillars and their own pupae ahead of kin larvae and eggs;Citation14 they also feed caterpillars at the expense of their larvae when food is scarce.Citation19 Sometimes workers treat M. rebeli caterpillars and pupae as if they were queen ants.Citation9 This social elevation is now explained by acoustical mimicry.
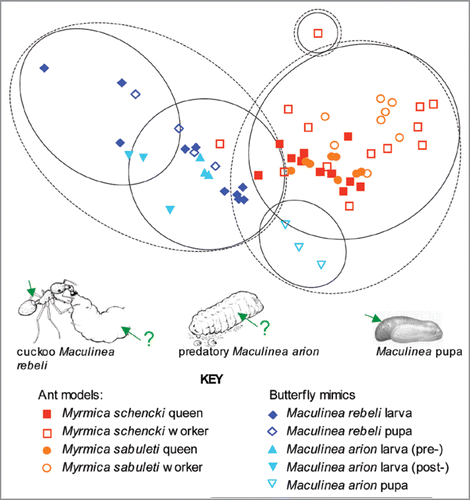
Evidence that Maculinea caterpillars mimic Myrmica worker stridulations was first obtained from highly alarmed experimental ants and caterpillars.Citation8 This nevertheless established that Myrmica larvae are mute and thus the caterpillars’ sounds, if confirmed as mimetic, were mimicking an adult attribute. It also showed that Maculinea acoustics bore a much closer resemblance to Myrmica ant stridulations than did mutualistic species of lycaenid butterfly. However, the match between Maculinea mimic and Myrmica model appeared to exist at the generic rather than the species level. Recently, using modern recording equipment and benign ant cultures, we found that queen Myrmica schencki ants can generate qualitatively different stridulations to those of their workers, and that when the sounds of queen M. schencki were played to workers, they stationed themselves on and around the speakers in a characteristic on-guard posture to that adopted when protecting real queens.Citation9 We also found that peaceful caterpillars and pupae of Maculinea rebeli emitted sounds that resembled those of their host in pulse length, dominant frequency, and pulse repetition frequency, but both were significantly closer to queen ant calls than to those of worker ants (). Playing butterfly pupal sounds to worker ants resulted in the same enhanced onguard behaviour induced by queen ant sounds; so, to a lesser (and not statistically significant) extent, did those of the caterpillars.Citation9 The closeness of mimicry achieved is remarkable given the different organs used by the butterfly to generate acoustics (): the ants rub a ridged sclerotised ‘plectrum’ against a file on an adjacent segment; the butterfly pupa has a paired tooth-and-comb organ, again on opposite abdominal segments; but the soft-bodied caterpillar probably emits sounds by muscular contractions and compressing air through trachaeae.Citation9,Citation20
Although the pupae of Maculinea arion inhabit similar niches within host colonies to those of M. rebeli, we predicted that its caterpillars, as predatory social parasites of Myrmica sabuleti, would posses less well-developed acoustics than its closely integrated congener.Citation10 We were incorrect. Although morphologically dissimilar, the two caterpillars produced closely overlapping sounds that could not be distinguishedCitation10 (). In fact the pupae of the two butterflies differed more, although the sample size is small. Again, just as the two types of mimic generated very similar sounds, so too did the same castes of their two model ant species.Citation10 In other words, although queens in Myrmica schencki colonies produce distinctive sounds that elicit royal treatment from their workers, the queens of M. sabuleti colonies make indistinguishable calls that presumably prompt the same enhanced on-guard behaviour from M. sabuleti workers as was observed in M. schencki. Thus in Myrmica ants, it appears that colony identity is established by distinctive semio-chemicals,Citation21 but the top hierarchical status of the queens within each colony is signalled, at least in part, by acoustics.Citation9,Citation10 The young stages of Maculinea appear to mimic both processes: caterpillars infiltrate and are accepted as members of host societies by employing chemical mimicry,Citation15–Citation17 but once established raise their social status above that of immature ants by using acoustical mimicry.Citation9,Citation10
Because acoustical mimicry of Myrmica by Maculinea signals caste rather than nest-mate recognition, and functions at a generic level, we consider it unlikely that local arms races between host and parasite will have evolved in this trait. We are, however, largely ignorant about how acoustical communication functions even in ant societies. For example, this was the first unequivocal demonstration for any ant species that the queens produce different sounds from workers and that the workers responded differently to them. Just how variably and how frequently different castes make distinctive sounds to elicit particular responses in undisturbed natural ant colonies are questions for future study. Moreover, acoustics, pheromones or tactile behaviour are known to be employed in combinations to moderate the information transmitted:Citation3 if acoustical communication is more informative than was generally recognised, there may be further dimensions of sophistication available for information exchange between society members. If confirmed, it also seems likely that acoustical mimicry will have evolved in other groups of social parasite as a means of infiltrating or surviving within ant societies, particularly in taxa, such as Staphilinid beetles and Orthoptera, where the non-parasitic species are capable of sound production.
Figures and Tables
Figure 1 MDS plot of the normalised Euclidean distances of the mean pulse length, dominant frequency, and pulse repetition frequency of the acoustical signals made by individual queens and workers of two Myrmica ant species and by the pupae and caterpillars of their Maculinea butterfly mimics. M. rebeli is a cuckoo social parasite, fed directly by worker ants; M. arion is a predator of ant brood, less closely integrated with its host society. The final instar larval stages of M. arion are separated into ‘pre-adoption’ before the larvae had contact with ants and ‘post-adoption’ recorded after they were in contact with ants. The contours indicate the normalised Euclidean distance separating the groups. ANOSIM indicates significant intra-specific separation between queen and worker acoustics in each ant species (p < 0.001) but no inter-specific differences in either the queen or worker sounds of the two species. The sounds of both Maculinea species’ pupae and caterpillars were significantly closer to queen Myrmica ant stridulations than to those of worker ants. Green arrows indicate known or presumed source of signals
Acknowledgements
This research was supported by CLIMIT under the EU-Biodiversa funding scheme.
Addendum to: and
References
- Elmes GW. Biological diversity of ants and their role in ecosystem function Lee Kim Sun 96: Proceedings of Inaugural Seminar of KIBIO 1996 Chonbuk, Korea 33 - 48
- Thomas JA, Schönrogge K, Elmes GW. Specialisations and host associations of social parasites of ants. Fellowes Holloway Rolff 05 Insect Evolutionary Ecology, CABI, Wallingford 2005; 479 - 518
- Hölldobler B, Wilson EO. The Ants. 1992; Berlin, Springer
- Nash DR, Boomsma JJ. Communication between hosts and social parasites. d’Ettorre Hughes 08 Sociolbiology of Communication 2008; Oxford Oxford University press, 55 - 80
- Hughes DP. The extended phenotype within the colony and how it obscures social communication. d’Ettorre Hughes 08 Sociolbiology of Communication 2008; Oxford Oxford University press, 171 - 190
- Schönrogge K, et al. Ecology Letters 2006; 9:1032
- Thomas JA, Settele J. Nature 2004; 432:283 - 284
- De Vries PJ, Cocroft RB, Thomas JA. Comparison of acoustical signals in Maculinea butterfly caterpillars and their obligate host Myrmica ants. Biol J Linn Soc 1993; 49:229 - 238
- Barbero F, Thomas JA, Bonelli S, Balletto E, Schönrogge K. Queen ants make distinctive sounds that are mimicked by a butterfly social parasite. Science 2009; 373:782 - 785 a
- Barbero F, Thomas JA, Bonelli S, Balletto E, Schönrogge K. Acoustical mimicry in a predatory social parasite of ants. J Exp Biol 2009; 212:4084 - 4090
- Elmes GW, Thomas JA, Munguira ML, Fiedler K. Larvae of lycaenid butterflies that parasitise ant colonies provide exceptions to normal insect growth rules. Biol J Linn Soc 2001; 73:259 - 278
- Thomas JA. Larval niche selection and evening exposure enhance adoption of a predacious social parasite, Maculinea arion (large blue butterfly), by Myrmica ants. Oecologia 2002; 122:531
- Elmes GW, Thomas JA, Wardlaw JC. Larvae of Maculinea rebeli, a Large Blue butterfly, and their Myrmica host ants: wild adoption and behaviour in ant nests. J Zool 1991; 223:447 - 460
- Thomas JA, Elmes GW, Wardlaw JC. Polymorphic growth in larvae of the butterfly Maculinea rebeli, a social parasite of Myrmica ant colonies. Proc R Soc Lond B 1998; 265:1895 - 1901
- Thomas JA, Wardlaw JC. Oecologia 1992; 91:101
- Akino T, Knapp JJ, Thomas JA, Elmes GW. Chemical mimicry and host specificity in the butterfly Maculinea rebeli, a social parasite of Myrmica ant colonies. Proc R Soc Lond B 1999; 266:1419 - 1426
- Schönrogge K, Wardlaw JC, Peters AJ, Everett S, Thomas JA, Elmes GW. Changes in chemical signature and host specificity from larval retrieval to full social integration in the myrmecophilous butterfly Maculinea rebeli. J Chem Ecol 2004; 30:91 - 107
- Nash DR, Als TD, Maile R, Jones GR, Boomsma JJ. A mosaic of chemical coevolution in a large blue butterfly. Science 2008; 319:88 - 90
- Elmes GW, Wardlaw JC, Schönrogge K, Thomas JA. Food stress causes differential survival of socially parasitic larvae of Maculinea rebeli (Lepidoptera, Lycaenidae) integrated in colonies of host and nonhost Myrmica species (Hymenoptera, Formicidae). Ent Exp et Appl 2004; 110:53 - 63
- Schurian KG, Fiedler K. Zur Biologie von Polyommatus (Lysandra) dezinus (De Freina &Witt) (Lepidoptera: Lycaenidae). Nachr Entomol Vereins Apollo 1994; 14:339 - 353
- Elmes GW, Akino T, Thomas JA, Clarke RT, Knapp JJ. Interspecific differences in cuticular hydrocarbon profiles of Myrmica ant species are sufficiently consistent to explain host specificity in Maculinea (Large blue) butterflies. Oecologia 2002; 130:525 - 535