Abstract
The study of expectations of reward helps to understand rules controlling goal-directed behavior as well as decision making and planning. I shall review a series of recent studies focusing on how the food gathering behavior of honeybees depends upon reward expectations. These studies document that free-flying honeybees develop long-term expectations of reward and use them to regulate their investment of energy/time during foraging. Also, they present a laboratory procedure suitable for analysis of neural substrates of reward expectations in the honeybee brain. I discuss these findings in the context of individual and collective foraging, on the one hand, and neurobiology of learning and memory of reward.
Expectations of Reward
An expectation is said to be “a strong hope or belief that something that you want will happen”, or the action “to anticipate or look forward to the occurrence of an event.”Citation1 Clearly, the notions of expectation and anticipation are linked to each other and it is taken for granted that these two words are interchangeable. However, although one needs to expect in order to anticipate, expecting per se does not imply anticipation. In psychology, the term ‘incentive’ designates internal correlates of specific rewards guiding the behavior of subjects pursuing such rewards.Citation2 Incentive denotes what is referred to as a subject’s expectation of reward.Citation2 Animals develop memories of specific properties of reward,Citation2–Citation4 alongside memories arising from the contingency between salient stimuli (as conditioned stimuli, or CSs) and reward (as an unconditioned stimulus, or US). Behaviorally, an expectation of reward is seen as an adjustment of a response which depends upon the formation and subsequent activation of memories about specific properties of reward, whereas the recollection of such memories is triggered by the cues and events predicting reward. In this scheme, expectations of reward are determined by past reward experience and can guide reward-induced behavior.Citation2
The Study of Reward Expectations
Studying reward expectations helps us to understand rules controlling goal-directed behavior as well as decision making and planning. Early studies showed that non-human primatesCitation5 and rodentsCitation2,Citation6 learn to expect specific outcomes and that these expectations are linked to specific magnitudes or kinds of rewards. Monkeys trained in a simple choice task show “disappointment, hesitation, and searching behavior” when they find a non-preferred food item where a preferred food item used to be,Citation5 and the running time of rats in a runway changes dramatically when they experience a sudden shift in reward magnitude.Citation2,Citation6 Since these initial findings, reward expectations have extensively been addressed in pigeons,Citation7 rodents,Citation8,Citation9 non-human primatesCitation10 and humans.Citation11 However, very little is known about reward expectations in invertebrate species. Here, I shall review a series of four recent studies focusing on how the food gathering behavior of honeybees depends upon expectations of reward. The first two studies concern the behavior of free-flying bees foraging under conditions closely mimicking a natural situation.Citation12,Citation13 Using an approach frequent in behavioral ecology, they helped to ponder the role of reward expectations in the ecology of foraging. The remaining two studies concern a laboratory procedure suitable for analysis of neural substrates of reward expectations in the honeybee brain.Citation14,Citation15 They made use of an approach frequent in the study of reward expectations, which relies heavily on experiments with restrained subjects made under highly controlled conditions. Using this approach, for example, studies in mammals have shown that interaction between the basolateral complex of the amygdala and the orbitofrontal cortex is necessary for development and subsequent use of reward expectations involved in goal-directed behaviours.Citation4,Citation11,Citation16,Citation17 I shall discuss these studies in the context of individual and collective foraging, on the one hand, and the neurobiology of learning and memory of reward.
Reward Expectations in Foraging Honeybees
Honeybees live in large colonies whose primary source of energy is the nectar found within flowers. Nectar offer varies continuously, both in space and time, depending on species-specific flowering patterns, weather conditions, and the activity of other pollinators.Citation18–Citation23 In spite of such variability, honeybees gather energy efficiently using their learning and memory skills.Citation24,Citation25 They learn, for example, the location and time of day when flowers are productive as well as their odors, colors and shapes.Citation26–Citation30 But, can they also learn that reward level increases or decreases over time? In a recent study,Citation12 we trained bees to forage individually on an artificial flower patch offering increasing (small-medium-large), decreasing (large-medium-small) or constant (small, medium or large) reward levels (). Next, after a long foraging pause, we recorded how persistently they searched for food at the patch in the absence of reward. We found that the bees that had previously experienced increasing reward levels searched for food more persistently than the bees that had experienced decreasing reward levels (). This difference could not be explained by the bees’ most recent reward experience or by the total amount of reward that they had previously collected. This conclusion was drawn from the fact that the bees that had collected small, medium and large constant rewards equally searched for food during testing (even if the last reward experience and the total amount of reward that they had previously collected were clearly different, ). We also found that the difference in persistence between the increasing and decreasing groups was independent of classical and/or operant associations between the offered reward and its predicting signals (see in Gil et al.Citation12). Taken together, the results of this study showed that bees can learn that the level of reward either increases or decreases over time and, subsequently, adjust their persistence during food searches accordingly. Can they also learn the magnitude of reward variations? We addressed this question in a second studyCitation13 in which we trained bees to forage individually on an artificial flower patch offering either a large (small-large) or a small (either small-medium or medium-large) increase in reward level (). After a long foraging pause following training, we recorded how persistently they searched for food at the patch in the absence of reward. We found that the bees that had previously experienced a large increase in reward level searched for food more persistently than the bees that had experienced a small increase in reward level (). As before, this difference could not be explained by the bees’ most recent reward experience or by the total amount of reward that they had previously collected. Honeybees, therefore, adjust their persistence to search for food in relation to both the sign and magnitude of past variations in reward level.Citation12,Citation13
The outcome of the above experiments is shown schematically in . When honeybees forage on a flower patch offering variable reward levels, two parallel learning processes take place. On the one hand, bees learn the sign and magnitude of reward variations across successive foraging visits. They may do this using a build-in change detector that computes the difference in reward magnitude across foraging events. This computation leads to an estimate of an expected reward; we refer to such estimate as to a reward memory. On the other hand, bees associate the offered reward (as the US) with signals and cues present at the feeding site (as CSs) and an associative memory is formed. When a bee visits the feeding site after a long foraging pause, these memories are retrieved by reward-predicting stimuli. Associative memories are revealed through the bee’s choice behaviour,Citation12 whereas reward memories are revealed through the bee’s persistence to search for food in the absence of reward.Citation12,Citation13 Hence, foraging honeybees adjust their investment of time/energy during food searches in relation to both the sign and magnitude of past variations in the level of reward. An increase in reward level leads to the formation of expectations of reward enhancing a forager’s reliance on a food source, and the strength of such reliance increases together with the magnitude of the past increase in reward level. This ability can make it more likely for them to successfully compete with other flower pollinators for limited resources as well as to maximize their individual rates of food collection by increasing their chances of finding food when forage is scarce. Because honeybees are social insects, one might ask how the colony as a whole benefits from a honeybee’s ability to develop expectations of reward. Such ability might enhance a colony’s selectivity among—variable—nectar sources. It would be interesting to examine the within-hive individual behavior as well as the pattern of collective behavior of honeybees foraging on multiple feeders offering increasing, decreasing and constant reward levels. One might also compare the ability of different species of bees to develop and use reward memories, and study how such ability relates to the specificities of their particular environments.
Optimal foraging theory attempts to predict foraging behavior in situations where sources of variable quality are heterogeneously distributed in space (reviewed in ref. Citation31). According to theory, foragers assess feeding site quality using an optimization rule that tends to maximize the rate of net energy intake.Citation32 Thus, a forager’s investment of time/energy at a given patch is positively correlated to food availability.Citation32–Citation34 This rule does not capture a forager’s investment of time/energy in the absence of reward.Citation31 When a honeybee searches for food at an empty source, the cost of searching influences its behavior in a way that memories on past reward experiences at the site will help the forager to determine how much time/energy is ought to be invested in the ongoing task. The results presented above show that a honeybee’s persistence to search for food on a negative energy budget relies on its—already developed—expectations of reward.Citation12,Citation13 Effort has been made to build models incorporating learning and memory phenomena into the context of foraging. One such model incorporates reward level variability into the forager’s evaluation of food patch quality.Citation35 It predicts that the foraging behavior of animals that have previously experienced variable rewards at a given patch will depend upon their memories of either the most recent reward level or the average reward level experienced, depending on the time elapsed since the last encounter with reward.Citation35 Our results do not match predictions from this model.Citation12,Citation13 Therefore, an alternative model is needed to explain how honeybees use reward memories during foraging.
Reward Expectations in Harnessed Bees
In addressing neural correlates of reward expectations in honeybees, one has to find a behavioral correlate of reward memories suitable for laboratory studies. Such behavioral correlate can be the honeybee proboscis extension response (PER).Citation36–Citation38 This response allows bees to gather sugar solution and is triggered when the gustatory receptors of the antennae, proboscis and tarsi are stimulated with sucrose.Citation37 In a recent study,Citation14 we asked whether harnessed bees can learn the sign of reward variations so as to subsequently adjust their PERs. We used an experimental design similar to that of our initial experiment with free-flying bees.Citation12 We first trained bees by coupling the stimulation of one antenna with either increasing, decreasing or constant reward levels offered to their proboscis throughout consecutive learning trials (). We then recorded the bees’ PE reaction-time to sucrose stimulation of the antenna in the absence of reward. We found that the bees that had experienced increasing reward levels subsequently extended their probosces earlier than the bees that had experienced decreasing or constant reward levels (). These results could not be accounted for by the bees’ most recent experience or the total amount of reward that they received during training. The bees that had experienced small, medium or large constant rewards showed similar reaction-times, although their last reward experience and the total amount of received reward were different (). Therefore, one can conclude that harnessed bees can learn that reward level increases or decreases over time and adjust their PERs accordingly.Citation14 But, further studies addressing neural correlates of reward memories in harnessed bees require within-animal controls. This is because recordings of neural activity are variable and, therefore, a reference from the same experimental subject is necessary for analysis of responses to any given stimulus. In a new series of experiments, we aimed to incorporate within-animal controls into our laboratory procedure. To this end, we asked whether bees can learn side-specifically that reward level increases or decreases over time.Citation15 Side-specific learning is well documented in honeybees.Citation39–Citation42 We developed a side-specific training in which bees were trained by coupling stimulation of one antenna with increasing reward levels and stimulation of the other antenna with decreasing reward levels throughout consecutive learning trials (). Next, at different times following training, we recorded the bees’ PE reaction-time to sucrose stimulation of each antenna in the absence of reward. We found that the bees extended their probosces earlier after stimulation of the antenna that had been linked to increasing reward levels than after stimulation of the antenna that had been linked to decreasing reward levels (). Therefore, bees can also learn side-specifically that reward increases or decreases over time. They develop both short- and long-term side-specific reward memories, and the long-term memories are extinguished by repetitive stimulation of their antennae (). Also, we found that these side-specific adjustments of PE response involve an interplay between gustatory and mechanosensory input, and correlate well with the activity of muscles responsible for controlling the movements of the proboscis.Citation15 Taken together, these findings constitute a basis on which further analyses of reward memories can be built. Such analyses will include within-animal controls and a physiological correlates of a robust behavioral measure.
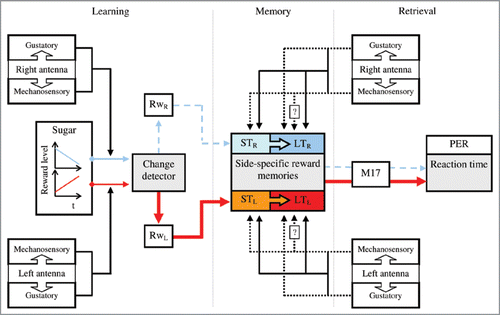
The events involved in this side-specific learning are schematically shown in . Bees learn to associate gustatory and mechanical stimulation of each antenna with either increasing or decreasing rewards offered to their probosces throughout consecutive learning trials. They do this using a built-in change detector that computes differences in reward level (linked to each antenna) across feeding events. This computation leads to the formation of an internal estimate of an expected reward associated with each input side, and then, to the formation of side-specific reward memories. After training, gustatory and mechanosensory input can activate both short- and long-term side-specific reward memories. Activation of such memories leads to side differences in a honeybee’s PE reaction-time, also evinced by activity of the muscles (M17s) involved in movement of the proboscis. It would be interesting to address how the magnitude and frequency of reward variations relate to the adjustment of a honeybee’s PER as well as the relative involvement of mechanical and gustatory inputs in side-specific learning. In addition, the fact that the above procedure allows analysis of within-animal behavioral correlates of reward memories makes it suitable for pharmacological, electrophysiological and optophysiological study of neural substrates underlying these memories. These studies can be combined with neuro-anatomical studies identifying brain areas where projections of gustatory receptors from the antenna and proboscis, on the one hand, and mechanosensory receptors from the antenna, on the other, converge. Previous studies showed that gustatory receptors of the antenna project into the ipsilateral antennal lobe (AL), dorsal lobe (DL) and suboesophageal ganglion (SOG).Citation43 The gustatory receptors of the proboscis project into the SOG and ascend to the DLs.Citation44 The mechanoreceptors of the antenna project into the ipsilateral DL and SOG.Citation43,Citation45 Hence, the DL and the SOG seem the first-order neuropils for processing mechanosensory and gustatory input from the antennae and the proboscis. Electrophysiological and optophysiological experiments will address how neural activity in these neuropils relates to adjustment of a honeybee’s PER occurring after activation of reward memories. Pharmacological approaches would also prove fruitful in this context. For example, it would be interesting to evaluate the role of the octopamine (OA, a bioamine involved in associative learning, memory retrieval, and food arousal in honeybees)Citation46–Citation49 during formation and retrieval of reward memories.
Conclusions
The results of the above studies show that honeybees learn the sign and magnitude of reward variations and develop long-term reward expectations allowing adjustment of time/energy investment during foraging. The results also show that honeybees adjust their PE reaction-time in relation to the sign of reward variations; this form of learning involves the joint action of gustatory and mechanosensory input of the antennae, and can be side-specific. These studies constitute a basis on which three lines of investigation can be built. The first line concerns the role of reward expectations in honeybee foraging. In particular, further experiments are needed to address how the colony as a whole benefits from a honeybee’s ability to develop expectations of reward. The second line of investigation concerns the question of how reward expectations can be incorporated into theoretical accounts of individual and collective foraging. The third line concerns identifying neural substrates involved in the development of reward memories in honeybees. Progress in these three lines of investigation will bring together behavioral, theoretical and physiological data to better understand the role and underlying mechanisms of reward memories in honeybees.
Figures and Tables
Figure 1 (A) Reward schedules offered during training by the artificial flower patch in the first experiment.Citation12 Each bee collected either increasing (I, small-medium-large), decreasing (D, large-medium-small), small (S), medium (M), or large (L) reward levels throughout successive visits to the patch. (B) Persistence (mean ± s.e.m., in s) to search for food in the absence of reward 24 h after training for the five different reward schedules (increasing, I; decreasing, D; small, S; medium, M; and large, L). Asterisks indicate planned comparisons p < 0.05. (Modified from Gil et al.Citation12). (C) Increasing reward schedules offered during training by the artificial flower patch in the second experiment.Citation13 Each bee experienced either a large increase in reward level, from small to large (S/L), or a small increase in reward level, either from small to medium (S/M) or from medium to large (M/L). (D) Persistence (mean ± s.e.m., in s) to search for food in the absence of reward 24 h after training for the three different reward schedules (S/L, S/M and M/L). Different letters indicate LSD multiple comparisons p < 0.05 after one-way ANOVA. (Modified from Gil and De MarcoCitation13). (B, D) Number of tested bees is given within the bars.
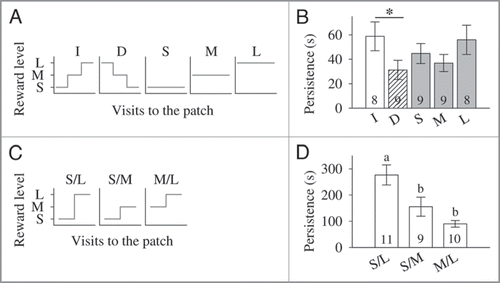
Figure 2 Schematic representation of the effects of constant (grey lines), decreasing (blue lines), and increasing sugar reward levels (red and orange lines for either a large or small increase in reward level, respectively) on a honeybee’s foraging behavior. A built-in change detector computes differences in reward level across foraging events. This leads to the formation of a specific reward memory. In parallel, bees associate the reward with stimuli present at the food source (as the CS), and an associative memory is formed. After a long foraging pause, the reward’s related stimuli retrieve these two forms of memory. Associative and reward memories are evinced through honeybees’ choice behavior and foraging persistence, respectively. In this scheme, differences in persistence are interpreted as evidence of different expectations of reward.
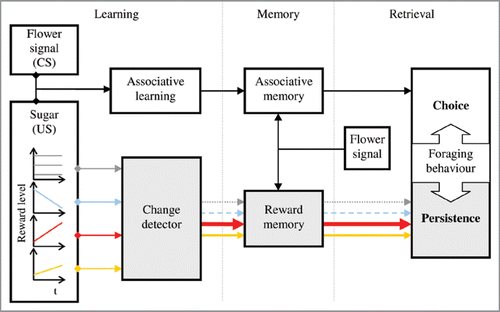
Figure 3 (A) Reward schedules offered during training.Citation14 Each bee was presented with either increasing (I, small-medium-large), decreasing (D, large-medium-small), small (S), medium (M) or large (L) reward levels throughout three consecutive training trials (first, an antenna was stimulated with sucrose, and then, when the bee extended the proboscis it was fed with a given amount of sugar reward). (B) PE reaction-time (mean ± s.e.m., in ms, defined as the time between the stimulation of the antenna and the first movement of the proboscis) 24 h after training for the five different reward schedules (increasing, I; decreasing, D; small, S; medium, M; and large, L). Different letters indicate Dunn’s multiple comparisons p < 0.001. Number of tested bees is given within the bars. (Modified from Gil et al.Citation14) (C) Side-specific training in which the stimulation of each antenna was coupled with either increasing or decreasing reward levels.Citation15 Each antenna was stimulated three times in an alternate way. For example, in the 1st trial we stimulated the left antenna and fed the bee with a small reward; in the 2nd trial we stimulated the right antenna and fed the bee with a large reward; in the 3rd trial we stimulated the left antenna and fed the bee with a medium reward; and so on. (D) PE reaction-time (mean ± s.e.m.) following the stimulation of the antenna that was linked to increasing rewards (white bars) and the antenna that was linked to decreasing rewards (dashed bars). After training, the bees were divided into two groups. One group was tested four times (1, 2, 3 and 24 h after training) under extinction conditions (panels in gray), and the other group was tested only once 24 h after training (panel in white, “we” stands for without extinction). Asterisks indicate Wilcoxon signed-rank test p < 0.01. Number of tested bees is given in the upper right corner of each panel. (Modified from Gil et al.Citation15).
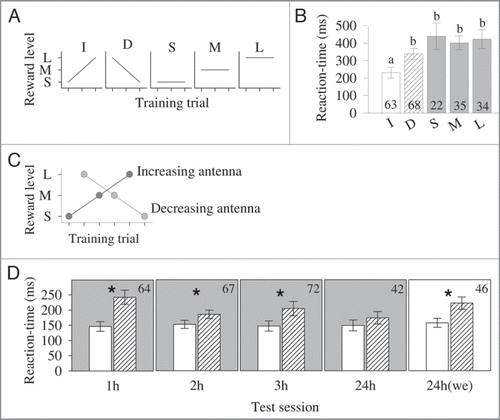
Figure 4 Schematic representation of the events involved in side-specific learning. When a honeybee experiences gustatory and mechanical stimulation of each antenna coupled with either increasing or decreasing rewards (red and blue lines, respectively) throughout consecutive training trials, a built-in change detector computes the differences in reward level associated with each antenna. This computation leads first to internal estimate of an expected reward associated to the input side (RwL, RwR), and then to side-specific reward memories. During memory retrieval, the combined stimulation of gustatory and mechanosensory receptors of the antenna (solid lines) activates both short- and long-term (ST, LT) side-specific reward memories. Mechanosensory stimulation of the antennae retrieves short-, and probably also long-term, side-specific reward memories (dashed lines). Specific gustatory input alone leads to the retrieval of long-term side-specific reward memories (dashed lines). The activation of such memories leads to side differences in a honeybee’s PE reaction-time, also evinced by the activity of the muscles M17s.
Acknowledgements
I am indebted to R.J. De Marco for his permanent encouragement, fruitful discussions and valuable comments. I thank R. Menzel for his helpful comments and support. This work was supported by the Deutsche Forschungsgemeinschaft (DFG).
References
- Merriam-Webster’s Collegiate Dictionary 1995; Springfield MA, USA Merriam-Webster, Inc
- Logan FA. Incentive 1960; New Haven Yale University Press
- Tolman EC. Koch S. Principles of purposive behaviour. A study of a science 1959; New York McGraw-Hill Psychology Vol. 2
- Schultz W. Multiple reward signals in the brain. Nature Rev Neurosc 2000; 1:199 - 207
- Tinklepaugh OL. An experimental study of representative factors in monkeys. J Comp Psychol 1928; 8:197 - 236
- Crespi LP. Quantitative variation in incentive and performance in the white rat. Am J Psychol 1942; 40:467 - 517
- Peterson GB, Wheeler RL, Armstrong GD. Expectancies as mediators in the differential reward conditional discrimination performance of pigeons. Anim Learn Behav 1978; 6:279 - 285
- Trapold MA. Are expectancies based upon different positive reinforcing events discriminably different?. Learn Motiv 1970; 1:129 - 140
- Holland PC, Straub JJ. Differential effects of two ways of devaluing the unconditioned stimulus after Pavlovian appetitive conditioning. J Exp Psychol 1979; 5:65 - 78
- Watanabe M, Cromwell H, Tremblay L, Hollerman JR, Hikosaka K, Schultz W. Behavioral reactions reflecting differential reward expectations in monkeys. Exp Brain Res 2001; 140:511 - 518
- O’Doherty J, Kringelbach ML, Rolls ET, Hornak J, Andrews C. Abstract reward and punishment representations in the human orbitofrontal cortex. Nat Neurosc 2001; 4:95 - 102
- Gil M, De Marco RJ, Menzel R. Learning reward expectations in honeybees. Learn Mem 2007; 14:491 - 496
- Gil M, De Marco RJ. Honeybees learn the sign and magnitude of reward variations. J Exp Biol 2009; 212:2830 - 2834
- Gil M, Menzel R, De Marco RJ. Does an Insect’s Unconditioned Response to Sucrose Reveal Expectations of Reward?. Tags for format 3PLoS ONE 2008; 3:2810 doi: 10.1371/journal.pone.0002810.
- Gil M, Menzel R, De Marco RJ. Side-specific reward memories in honeybees. Learn Mem 2009; 16:426 - 432
- Holland PC, Gallagher M. Amygdala-frontal interactions and reward expectancy. Curr Op Neurobiol 2004; 14:148 - 155
- Gallagher M, McMahan RW, Schoenbaum G. Orbitofrontal cortex and representation of incentive value in associative learning. J Neurosc 1999; 19:6610 - 6614
- Núñez JA. Nectar flow by melliferous flora and gathering flow by Apis mellifera ligustica. J Insect Physiol 1977; 23:265 - 275
- Teuber LR, Barnes DK. Environmental and genetic influences on quantity and quality of alfalfa nectar. Crop Sci 1979; 19:874 - 878
- Vogel S. Lange OL, Nobel PS, Osmond CB, Ziegler H. Ecophysiology of zoophilic pollination. Physiological plant ecology III 1983; Berlin, Heidelberg, New York Springer
- Baker HG, Baker I. Bentley B, Elias T. A brief historical review of the chemistry of floral nectar. The Biology of Nectaries 1983; New York Columbia University Press
- Real LA, Rathcke BJ. Patterns of individual variability in floral resources. Ecol 1988; 69:728 - 735
- Rathcke B, Lacey EP. Phenological patterns of terrestrial plants. Annu Rev Ecol Sys 1985; 16:179 - 214
- Gould JL, Gould CG. The Honey Bee 1988; New York Scientific American Library
- Seeley TD. The Wisdom of the Hive 1995; Cambridge, Mass Harvard University Press
- Wahl O. Neue Untersuchungen über das Zeitgedächtnis der Bienen. Z vergl Physiol 1932; 16:529 - 589
- Kleber E. Hat das Zeitgedächtnis der Bienen biologische Bedeutung?. Z vergl Physiol 1935; 22:221 - 262
- Kolterman R. Lern- und Vergessensprozesse bei der Honigbiene-aufgezeigt anhand von Duftdressuren Z vergl. Physiol 1969; 63:310 - 334
- Frisch K. von. The Dance Language and Orientation of Bees 1967; Harvard University Press
- Menzel R. Kesner RP, Olten DS. Learning, Memory and ‘Cognition’ in Honey Bees. Neurobiology of Comparative Cognition 1990; Hillsdale, NJ Erlbaum,
- Pyke GH. Optimal foraging theory: a critical review. Annu Rev Ecol Syst 1984; 15:523 - 575
- Charnov EL. Optimal foraging, the marginal value theorem. Theor Pop Biol 1976; 9:129 - 136
- Schmid-Hempel P, Kacelnik A, Houston AJ. Honeybees maximize efficiency by not filling their crop. Behav Ecol Sociobiol 1985; 17:61 - 66
- Varjú D, Núñez JA. What do foraging honeybees optimize?. J Comp Physiol A 1991; 169:729 - 736
- Devenport LD, Devenport JA. Time-dependent averaging of foraging information in least chipmunks and golden-mantled ground squirrels. Anim Behav 1994; 47:787 - 802
- Takeda K. Classical conditioned response in the honey bee. J Insect Physiol 1961; 6:168 - 179
- Kuwabara M. Bildung des bedingten Reflexes von Pavlovs Typus bei der Honigbiene, Apis mellifica. J Fac Hokkaido Univ Ser VI Zol 1957; 13:458 - 464
- Bitterman ME, Menzel R, Fietz A, Schäfer S. Classical conditioning of proboscis extension in honeybees (Apis mellifera). J Comp Psychol 1983; 97:107 - 119
- Masuhr T, Menzel R. Wehner R. Learning Experiments on the Use of Side-Specific Information in the Olfactory and Visual System in the Honeybee (Apis mellifica). Information Processing in the Visual Systems of Arthropods 1972; Berlin-Heidelberg-New York Springer
- Macmillan CS, Mercer AR. An investigation of the role of dopamine in the antennal lobes of the honeybee, Apis mellifera. J Comp Physiol 1987; 160:359 - 366
- Sandoz J-C, Hammer M, Menzel R. Side-specificity of olfactory learning in the honeybee: US input side. Learn Mem 2002; 9:337 - 348
- Giurfa M, Malun D. Associative Mechanosensory Conditioning of the Proboscis Extension Reflex in Honeybees. Learn Mem 2004; 11:294 - 302
- Suzuki H. Antennal movements induced by odour and central projection of the antennal neurons in 21:831 - 847
- Haupt SS. Das Gustatorische System und antennales Lernen in der Honigbiene (Apis mellifera L.) 2005; Germany Technischen Universit↦ Berlin, PhD Thesis,
- Maronde U. Common projection areas of antennal and visual pathways in the honeybee brain, Apis mellifera. J Comp Neurol 1991; 309:328 - 340
- Hammer M. An identified neuron mediates the unconditioned stimulus in associative olfactory learning in honeybees. Nature 1993; 366:59 - 63
- Hammer M, Menzel R. Multiple sites of associative odor learning as revealed by local brain microinjections of octopamine in honeybees. Learn Mem 1998; 5:146 - 156
- Mercer AR, Menzel R. The effects of biogenic amines on conditioned and unconditioned responses to olfactory stimuli in the honeybee, Apis mellifera. J Comp Physiol 1982; 145:363 - 368
- Braun G, Bicker G. Habituation of an appetitive reflex in the honeybee. J Neurophysiol 1992; 67:588 - 598