Abstract
Heterokairy refers to plasticity in the timing of onset of developmental events at the level of an individual. When two developmental stages do not share the same ecological niche, referred to as ‘ontogenetic niches’, the control of the niche shift through a change in developmental timing can be advantageous for the individual (e.g. when mortality risk is different in the two niches). Heterokairy can arise either from plasticity in developmental rate (ontogenetic shift) or by a purely behavioural decision (behavioural shift). Parasitic species living inside of their hosts often inherit the predators of their hosts. To cope with the predation risk on their hosts, parasites and parasitoids show either host-manipulation abilities or either host-leaving strategies. Nevertheless, leaving the host should be associated with developmental costs, since the parasitic individuals are usually unable to parasitize another host. This process is thus related to the classical trade-off between size and developmental time. Recent studies provided examples of behavioural heterokairy in invertebrates. The goal of this publication is to review and discuss recent results on developmental plasticity in parasitic species in an evolutionary perspective.
Heterokairy: Definition
Phenotypic plasticity in development time allows individuals to deal with unpredictable environments.Citation1 Heterokairy refers to plasticity in the timing of onset of developmental events at the level of an individual.Citation2 “Heterokairy” is the individual equivalent of evolutionary “heterochrony,”Citation3 and can arise either from plasticity in developmental rate (ontogenetic shiftCitation4,Citation5) or by a purely behavioral decision.Citation6,Citation7 Exposure to predators have been shown to influence prey life history switch points, including those related to the timing of hatching, metamorphosis and reproduction ().Citation8
Induced heterokairy in response to predation risk has been mainly studied in amphibians.Citation3 Plasticity in both timing of egg hatch and larval development has been shown. Early hatching of eggs has been reported for tree frogs (Hyla regilla) and cascade frogs (Rana cascadae) exposed to predatory leeches,Citation8 for wood frogs (Rana sylvatica) and American toads (Bufo americanus) when infected by water mold,Citation9,Citation10 and for the red-eyed tree frog (Agalychnis callidryas) in the presence of wasp or snake predation.Citation11,Citation12 In the whitefish, Coregonus sp., eggs hatch sooner than controls when exposed to cues emitted by Pseudomonas-infected eggs.Citation13 Feeding rate/predation risk trade-off seems to constrain the ontogenetic niche-shift (from littoral to pelagic zone) in the bluegill sunfish, Lepomis macrochirus.Citation14
Females of the egg-carrying spider, Scytodes pallida adjust egg-hatching time (the interval between oviposition and hatching) in response to the threat of predation by Portia labiata.Citation15 On the other hand, it has been reported a counter-intuitive case where juveniles of the crustacean gammarid Gammarus insensibilis exited the marsupial brood pouch of infected females significantly later than those from uninfected mothers, despite the fact that infected mothers are reportedly more susceptible to predation.Citation16
Response to Mortality Risk for Parasitic Species
Parasitic species living inside of their hosts often inherit the predators of their hosts.Citation17 Parasitoids are organisms that develop inside or on another organism (host); they feed on the host tissues and kill the host as a direct or indirect result of its development. For a long time, parasitic species were considered to live in a predictable environment (hosts) that does not require adaptive phenotypic plasticity. Thomas et al.Citation17 challenged this view by discussing the state-dependent strategies adopted by parasitic species to maximize their reproductive success according to external conditions.Citation17 The environment of parasites is made up of at least two dimensions, which are ecologically different: the host (i.e., the immediate environment) and the habitat of the host (i.e., the ecosystem).Citation17 For instance, it is frequently observed that parasite virulence is higher when the host is in poor condition or experiencing stressful conditions.Citation18 It may be possible for parasites to perceive the external environment of their hosts, such as the predation risk of their hosts, and adjust their strategy accordingly. Free-living crustaceans can evaluate the abundance of fish predators, without the need to see them directly, by detecting components of the fish mucus in the water.Citation19
The role of predation in the evolutionary ecology of animal communities has long been a focus of ecological research but, to date, sparse attention has been given to the responses of parasites when their hosts are victims of predation.Citation6 One option for parasitic species to reduce the risk of mortality by predation of its host is to reduce the encounter rate with potential predators of host by altering the behavior of the host,Citation20–Citation22 a process known as “host manipulation.”Citation23 A step further is the induction of unusual patterns in the host’s behavior in order to reduce encounters with asitoid specific natural enemies, such as hyperparasitoids, a process known as “usurpation.”Citation24–Citation26 The option would be to leave the endangered host.
Heterokairy as an Antipredator Strategy for Parasitic Species
Host-leaving event leads to “heterokairy” since the individual will end its endoparasitic development to enter another step of development. Timing of the onset of developmental events is plastic according to the mortality risks within and outside of the host.
One example of such anti-predatory strategy has been found for the hairworm Paragordius tricuspidatus (Nematomorpha: Gordiida). These worms parasitize mainly crickets, which they manipulate to commit “induced suicide” by jumping into the water. In the water, the worm comes out to freely live its adult life.Citation23 First, it has been shown that if a parasitized cricket that enters the water is eaten by a fish or a frog, the hairworm is able to escape not only from its insect host but also from the digestive tract of the predator.Citation27 The worm emerges alive from the mouth, gills or nose of the predators and continues its life cycle without any apparent fitness costs.Citation28 Moreover, worm emergence from the host has been shown to be induced by predatory notonectid.Citation6
Recently, it has been shown that the predation on host induced early emergence in the endoparasitic fly Endaphis fugitiva (Diptera: Cecidomyiidae).Citation7 As a larva, this aphid parasitoid develops inside the host body and emerges as a mature larva to pupate in the soil.Citation29 Plasticity in the timing of emergence allows the parasitoid to respond to mortality risk occurring while inside its host. Artificial injuries on host, as well as predator attack on host, induced parasitoid larvae to leave the host. When a parasitized aphid is bitten by a predatory Hemerobiid larva, the parasitoid larva quickly flees from its host. Moreover, it has been observed that the predator goes on feeding on the remains of the host’s body, while the parasitoid is emerging. This allows the parasitoid larvae to perform jumps and escape from further direct predator attack.Citation29
Costs of Heterokairy and Evolutionary Processes
Whether to grow larger at the cost of longer development time, or to develop more rapidly at the cost of reduced size is a classical trade-off in life history evolution. However, it might be that the developmental constraints on some parasitic species will influence the evolution of heterokairy. Indeed, in a few hymenopterous endoparasitoid clades (e.g., Microgastrinae, Braconidae), parasitoid larvae consume only a small fraction of available host tissues (primarily fat body and hemolymph) and emerge from the still living host by perforating the host cuticle with their mandibles and pupating externally.Citation30 It is hypothesized that developmental constraints might come from differences in the way species feed on host tissues: leading some species to emerge when fixed critical size is reached, while other species have to consume the entire host before emergence.Citation31 It might be that the evolution of heterokairy has been driven by the selective advantages of individuals that freed themselves from the constraint of total consumption of the host body before shifting to the next developmental step.
Organisms should switch life history stages when their mortality/growth ratio is lower in the following stage than the current stage.Citation8 Obviously, mortality risk due to host predation should be frequent enough to induce a selective pressure on the parasitic individual. Therefore, predators should not be able to discriminate between parasitized and unparasitized hosts. In the Endaphis system, we tested this assumption by studying the behavior of a single aphid predator foraging in a patch containing 3 healthy and 3 parasitized hosts (n = 13). We showed that the probability of being attacked was not related to the presence of a parasitoid larva in the host (). The bionomial response (attack or leave) of the predator to aphid encounter was not statistically different between the healthy and parasitized hosts (Wald = 0.872, df = 1, p = 0.35; Generalized Estimating Equations, binomial distribution, logit link function, “geepack” for RCitation32). This suggests that selective pressure on the parasitoid is not reduced by discrimination abilities from the predator.
If mortality risk is high enough, the benefit of saving its life against the costs associated with early emergence as well as the development and maintenance of sensory machinery is clearly biased towards the selection of responsive parasitoids. Working on the spitting spider system, Li showed that hatchings from predator exposed eggs were smaller than unexposed ones.Citation15 Muratori et al. did not find costs associated to early emergence in parasitic fly, Endaphis.Citation7 The energy cost of escape in itself is likely to be negligible, as shown for predator-induced early emergence in hairworms (Paragordius tricuspidatus), which produce similar offspring as controls.Citation16 While costs associated with heterokairy have been shown in many vertebrate systems,Citation8,Citation11 studies on induced niche-shift in invertebrate parasitic species are unfortunately too scarce to draw any conclusions.
Emphasis has been placed on studying adaptive developmental plasticity in the context of anti-predator defenses, but it is also likely to be effective in response to environmental heterogeneity in other factors (abiotic, pathogens, competition).Citation9 Several sources of selection for plasticity in itself might have had a positive effect on the conservation of this trait. On the evolutionary point of view, it might be that adaptive niche shift is a process common for concealed species that emerge as larvae to pupate outside their shelter/host. For example, Endaphis fugitiva belongs to the Cecidomyiidae family in which both predatory, parasitoid and phytophagous species are found.Citation29 Many of the phytophagous Cecidomyiids are galling insects. It would be interesting to test if adaptive niche-shift is present in these species.
Figures and Tables
Figure 1 Number of attacks on healthy and Endaphis parasitized hosts by the aphid predator, Micromus timidus. The aphid predator does not discriminate parasitized host, which keeps a strong selection pressure on the parasitoid for induced emergence response.
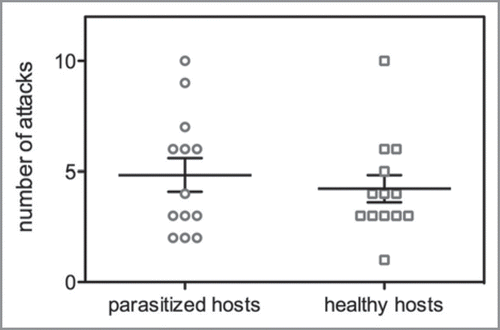
Table 1 Species for which heterokairy have been shown
References
- DeWitt TJ, Langerhans RB. DeWitt TJ, Scheiner SM. Integrated Solutions to Environmental Heterogeneity: Theory of Multimoment Reaction Norms. Phenotypic Plasticity: functional and conceptual approaches 2004; Oxford University Press
- Burggren W, Warburton S. Comparative developmental physiology: An interdisciplinary convergence. Annu Rev Physiol 2005; 67:203 - 223
- Warkentin KM. Oxygen, gills and embryo behavior: mechanisms of adaptive plasticity in hatching. Comp Biochem Physiol A 2007; 148:720 - 731
- Rowe L, Ludwig D. Size and timing of metamorphosis in complex life cycles: time constraints and variation. Ecology 1991; 72:413 - 427
- Spicer JI, Burggren WW. Development of physiological regulatory systems: altering the timing of crucial events. Zoology 2003; 106:91 - 99
- Sanchez MI, Ponton F, Misse D, Hughes DP, Thomas F. Hairworm response to notonectid attacks. Anim Behav 2008; 75:823 - 826
- Muratori FB, Borlée S, Messing RH. Induced niche shift as an anti-predator response for an endoparasitoid. Proc R Soc B 2010; on line first. doi: 10.1098/rspb.2009
- Chivers DP, Kiesecker JM, Marco A, DeVito J, Anderson MT, Blaustein AR. Predator-induced life history changes in amphibians: egg predation induces hatching. Oikos 2001; 92:135 - 142
- Gomez-Mestre I, Touchon JC, Warkentin KM. Amphibian embryo and parental defences and a larval predator reduce egg mortality from water mold. Ecology 2006; 87:2570 - 2581
- Gomez-Mestre I, Touchon JC, Saccoccio VL, Warkentin KM. Genetic variation in pathogen-induced early hatching of toad embryos. J Evol Biol 2008; 21:791 - 800
- Warkentin KM. Adaptive plasticity in hatching age: a response to predation risk trade-offs. Proc Natl Acad Sci USA 1995; 92:3507 - 3510
- Warkentin KM. Wasp predation and wasp-induced hatching of red-eyed treefrog eggs. Anim Behav 2000; 60:503 - 510
- Wedekind C. Induced hatching to avoid infectious egg disease in whitefish. Curr Biol 2002; 12:69 - 71
- Werner EE, Hall DJ. Ontogenetic habitat shifts in bluegill: The foraging rate-predation risk trade-off. Ecology 1988; 69:1352 - 1366
- Li DQ. Hatching responses of subsocial spitting spiders to predation risk. Proc R Soc B 2002; 269:2155 - 2161
- Ponton F, Duneau D, Sanchez M, Courtiol A, Terekhin AT, Budilova EV, et al. Effect of parasite-induced behavioral alterations on juvenile development. Behav Ecol 2009; 20:1020 - 1025
- Thomas F, Brown SP, Sukhdeo M, Renaud F. Understanding parasite strategies: a state-dependent approach?. Trends Parasitol 2002; 18:387 - 390
- Brown MJF, Loosli R, Schmid-Hempel P. Conditiondependent expression of virulence in a trypanosome infecting bumblebees. Oikos 2000; 91:421 - 427
- Forward RB, Rittschof D. Alteration of photoresponses involved in diel vertical migration of a crab larva by fish mucus and degradation products of mucopolysaccharides. J Exp Mar Biol Ecol 2000; 245:277 - 292
- Levri EP. The influence of non-host predators on parasite induced behavioral changes in a freshwater snail. Oikos 1998; 81:531 - 537
- Lafferty KD, Thomas F, Poulin R. Poulin R, Morand S, Skorping A. Evolution of host phenotype manipulation by parasites and its consequences. Evolutionary Biology of Host-Parasite Relationships: Theory Meets Reality 2000; Amsterdam Elsevier Science 117 - 127
- Haine ER, Rigaud T. Conflict between parasites with different transmission strategies infecting an amphipod host. Proc R Soc B 2005; 272:2505 - 2510
- Thomas F, Schmidt-Rhaesa A, Martin G, Manu C, Durand P, Renaud F. Do hairworms (Nematomorpha) manipulate the water seeking behaviour of their terrestrial hosts?. J Evol Biol 2002; 15:356 - 361
- Brodeur JN, McNeil J. Seasonal microhabitat selection by an endoparasite through adaptive modification of host behavior. Science 1989; 244:226 - 228
- Harvey JA, Kos M, Nakamatsu Y, Tanaka T, Dicke M, Vet L, et al. Do parasitized caterpillars protect their parasitoids from hyperparasitoids? A test of the ‘usurpation hypothesis’. Anim Behav 2008; 76:701 - 708
- Grosman AH, Janssen A, de Brito EF, Cordeiro EG, Colares F, Fonseca JO, et al. Parasitoid increases survival of its pupae by inducing hosts to fight predators. PLoS ONE 2008; 3:2276
- Ponton F, Lebarbenchon C, Lefèvre T, Biron DG, Duneau D, Hughes DP, et al. Parasite survives predation on its host. Nature 2006; 440:756
- Ponton F, Lebarbenchon C, Lefèvre T, Thomas F, Duneau D, Marché L, et al. Hairworms anti-predator strategy: a study of causes and consequences. Parasitol 2006; 133:631 - 638
- Muratori FB, Gagné RJ, Messing RH. Ecological traits of a new aphid parasitoid, Endaphis fugitiva (Diptera: Cecidomyiidae), and its potential for biological control of the banana aphid, Pentalonia nigronervosa (Hemiptera: Aphididae). Biol Control 2009; 50:185 - 193
- Harvey JA, Strand MR. The developmental strategies of endoparasitoid wasps vary with host feeding ecology. Ecology 2002; 83:2439 - 2451
- Harvey JA, Kadash K, Strand MR. Differences in larval feeding behavior correlate with altered developmental strategies in two parasitic wasps: implications for the size-fitness hypothesis. Oikos 2000; 88:621 - 629
- Yan J. geepack: Yet another package for generalized estimating equations. R-News 2002; 2/3:12 - 14 http://www.r-project.org/doc/Rnews/Rnews_2002-3.pdf