Abstract
Studies in several vertebrate species have shown that visual sensitivity and a number of other retinal phenomena are regulated by circadian mechanisms. For example, ultra-structural studies of 5 day old zebrafish larvae have shown that synaptic ribbons in photoreceptor terminals undergo dramatic diurnal alterations. These synaptic ribbons are very prominent during the day, but are almost completely absent at night. The implications of this circadian driven process on visual function are not well understood. We recently showed that larval zebrafish essentially lose visual responsiveness at night. This shut-down of retinal function at night is regulated by at least two mechanisms: the disassembly of synaptic ribbons in cone pedicles and a decrease of outer segment activity. Here, we summarize our recently reported observations and further discuss our hypothesis on how this phenomenon of shutting-down retinal function at night may provide a means for zebrafish larvae to conserve energy.
In the retina, photoreceptors and bipolar cells make specialized synapses called ribbon synapses. The ribbon is an electron-dense plate-like structure that extends from the site of transmitter release well into the synaptic terminal cytoplasm and tethers to a large number of regularly aligned synaptic vesicles.Citation1 Synaptic ribbons (SRs) relate to a tonic release of neurotransmitter and are thought to channel synaptic vesicles in an orderly, conveyor-belt fashion to the plasma membrane.Citation2,Citation3 The rate at which neurotransmitter is released at ribbon synapses is manifold higher than at conventional synapses.Citation4–Citation7
In larval (5–6 dpf) zebrafish, ultrastructural analysis of cone pedicles suggested that diurnal alterations of synaptic ribbons are circadian driven.Citation8 In our recent study,Citation9 we aimed at investigating the functional consequences of this structural plasticity. We first recorded electroretinograms (ERGs) at different times of the day and night using 0.5 second flashes of light. We observed stable ERG amplitudes during the day but a drastic reduction in ERG amplitudes 3 h after lights were turned OFF. Indeed, ERG responsiveness was severely suppressed at night as full field light flashes over 5 log units of light intensity did not evoke a significant increase in ERG amplitude. Light exposure at night could, however, partially restore visual responsiveness. When fish were exposed to light during the night, ERG amplitudes were substantial, although they were smaller as compared to ERG amplitudes recorded during the day.
To assess whether this day/night cycle of retinal responsiveness is circadian driven, we raised zebrafish larvae under normal light/dark conditions for 4 days and then maintained the fish in the dark for 2 days. Over the two days of dark, we found ERG amplitudes were much smaller during subjective night as compared to subjective day, indicating that retinal responsiveness is circadian driven.
Our study then focused on the dynamics of the loss and recovery of ERG responsiveness. At lights OFF at night, there is a gradual decrease in ERG amplitude over the first 1 h and the ERG is virtually abolished after 90 minutes. Conversely, exposure to light after 3 h of darkness at night steadily increased ERG amplitudes with significant changes observed within 10–20 minutes.
Next we asked how the loss of ERG responsiveness relates to the visual behavior of larval zebrafish. To test this, we performed two behavioral assays. The optokinetic reflex (OKR) measures eye movements of larval fish in response to black and white stripes moving in either direction. The visual-motor response (VMR) measures the motor activity of fish in response to light onset and offset. Both behavioral assays indicated that vision is essentially lost at night in zebrafish larvae. Further, in agreement with the ERG measurements, light exposure at night partially restored the visual behaviors.
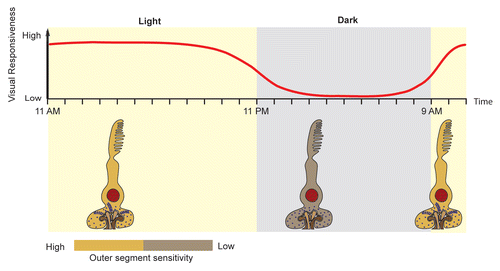
Do the changes in visual responsiveness we observe correlate with changes in SRs of the cone pedicles? EM sections of the retina of 5 day old larval retinas revealed that the numbers of SRs are high during the day when both the ERG signals and visual behavioral responses are robust. At night however, when SRs have disassembled, ERG and visual behavioral responses have essentially disappeared (see for summary). Further, dark-adaptation during the day did not initiate the disassembly of SRs, whereas continuous light exposure at night prevented the disassembly of SRs. Thus, the disassembly of SRs is one possible mechanism by which visual responsiveness in larval zebrafish is reduced at night.
We also asked whether outer segment activity of photoreceptors is regulated by a circadian rhythm. Thus, we measured ERG a-wave amplitudes in response to test flashes. Because the a-wave, which reflects the hyperpolarization of the outer segments in response to light, is largely masked by the depolarizing b-wave, we incubated the fish in a drug cocktail which had been previously shown to eliminate the b-wave in zebrafish.Citation10,Citation11 This set of experiments revealed that a-wave amplitudes are also decreased at night (although not completely) and thus, a decrease in outer segments activity is another mechanism by which visual responsiveness is regulated at night.
The dramatic fluctuations in visual responsiveness in larval zebrafish during the day and night were surprising considering that in adult zebrafish such fluctuations are relatively small.Citation12 So why do zebrafish larvae essentially shut-down their visual system at night? Our suggestion is that zebrafish larvae at 5 days may need to conserve energy because they have essentially depleted their yolk sac and are just beginning to eat. Support for this hypothesis is the observation that zebrafish larvae generally reduce their motor activity at night.Citation9,Citation13,Citation14 Furthermore, photoreceptors are depolarized in the dark which requires a significant amount of ATP,Citation15 estimated to be 108 ATPs/sec in a single mouse rod.Citation16 Further, photoreceptors also release transmitter continuously in the dark, and this too requires energy. Hence, it is safe to assume that the retina consumes a significant amount of energy. Thus, decreasing photoreceptor activity at night is likely to decrease energy consumption significantly in larval zebrafish.
Figures and Tables
Figure 1 Model summarizing circadian oscillations of visual responsiveness in larval zebrafish retina. Upper panel illustrates the degree of visual responsiveness that is observed both behaviorally by the optokinetic response test (OKR) and the visual-motor response (VMR) and electrophysiologically by electroretinogram (ERG) recordings from 5 day old zebrafish larval retina. Lower panel illustrates the decrease in photoreceptor outer segment sensitivity and the disassembly of synaptic ribbons in cone pedicles at night.
Acknowledgements
We thank Dr. Alan R. Adolph and Dr. Edward Soucy for their generous technical and intellectual help and Manija Emran for her help with the art work. This work was supported by National Institute of Health Grants EY000811 (J.E.D.) and F32 EY018044-01A2 (F.E.).
Addendum to:
References
- Rao-Mirotznik R, Harkins AB, Buchsbaum G, Sterling P. Mammalian rod terminal: architecture of a binary synapse. Neuron 1995; 14:561 - 569
- Bunt AH. Enzymatic digestion of synaptic ribbons in amphibian retinal photoreceptors. Brain Res 1971; 25:571 - 577
- Gray EG, Pease HL. On understanding the organisation of the retinal receptor synapses. Brain Res 1971; 35:1 - 15
- Heidelberger R, Heinemann C, Neher E, Matthews G. Calcium dependence of the rate of exocytosis in a synaptic terminal. Nature 1994; 371:513 - 515
- Stevens CF, Tsujimoto T. Estimates for the pool size of releasable quanta at a single central synapse and for the time required to refill the pool. Proc Natl Acad Sci USA 1995; 92:846 - 849
- von Gersdorff H, Vardi E, Matthews G, Sterling P. Evidence that vesicles on the synaptic ribbon of retinal bipolar neurons can be rapidly released. Neuron 1996; 16:1221 - 1227
- Heidelberger R, Thoreson WB, Witkovsky P. Synaptic transmission at retinal ribbon synapses. Prog Retin Eye Res 2005; 24:682 - 720
- Allwardt BA, Lall AB, Brockerhoff SE, Dowling JE. Synapse formation is arrested in retinal photoreceptors of the zebrafish nrc mutant. J Neurosci 2001; 21:2330 - 2342
- Emran F, Rihel J, Adolph AR, Dowling JE. Zebrafish lose vision at night. Proc Natl Acad Sci USA 2010; 107:6034 - 6039
- Emran F, Rihel J, Adolph AR, Wong KY, Kraves S, Dowling JE. OFF ganglion cells cannot drive the optokinetic reflex in zebrafish. Proc Natl Acad Sci USA 2007; 104:19126 - 19131
- Wong KY, et al. Glutamatergic Mechanisms in the outer retina of larval zebrafish: analysis of electroretinogram b- and d-waves using a novel prepartation. Zebrafish 2004; 1:121 - 131
- Li L, Dowling JE. Zebrafish visual sensitivity is regulated by a circadian clock. Vis Neurosci 1998; 15:851 - 857
- Prober DA, Rihel J, Onah AA, Sung RJ, Schier AF. Hypocretin/orexin overexpression induces an insomnia-like phenotype in zebrafish. J Neurosci 2006; 26:13400 - 13410
- Emran F, Rihel J, Dowling JE. A behavioral assay to measure responsiveness of zebrafish to changes in light intensities. J Vis Exp 2008; http://dx.doi.org/10.3791/923
- Wangsa-Wirawan ND, Linsenmeier RA. Retinal oxygen: fundamental and clinical aspects. Archives of ophthalmology 2003; 121:547 - 557
- Okawa H, Sampath AP, Laughlin SB, Fain GL. ATP consumption by mammalian rod photoreceptors in darkness and in light. Curr Biol 2008; 18:1917 - 1921