Abstract
The CWI pathway cross-talks with TOR and RAS in both the oxidative and glucose starvation responses. Mtl1 is the cell-wall protein in charge of sensing and regulating this response. Rom2 and Rho1, which are the upper elements in the pathway, mediate this signal. Several outputs are involved and required for this response, one of which, ribosomal gene expression, seems to be regulated by Sfp1, amongst other possible transcription factors. Moreover, cross-talk also occurs in a reverse flow from TOR and RAS to the CWI pathway. Thus Tor1 and Ras2 inhibition also activates Slt2 in the absence of the Mtl1 protein and assures the proper adaptive response to oxidation and glucose deprivation.
Mtl1 is a cell-surface protein of Saccharomyces cerevisiae with a very high similarity to Mid2.Citation1 Mtl1 localization has not been described to date, however its sequence contains a transmembrane domain and a very long domain rich in serine-threonine residues.Citation2 This domain could be a suitable region of O-mannosylation, as it is similar to that described for Mid2.Citation3 Mtl1 has been described as a molecule required for survival under oxidative stress conditions.Citation4 Mtl1 is a putative sensor responsible for transmitting the oxidative signal to the CWI (Cell Wall Integrity pathway) and thereby inducing the activation of Slt2: the last kinase in the cascade; it is also responsible for the depolarization of the actin cytoskeleton.Citation4 We have recently demonstrated that Mtl1 is required to inactivate both the Tor1 and Ras2 functions in response to glucose depletion and oxidative stress provoked by hydrogen peroxide.Citation5 This is how Mtl1 transmits these signals to Rom2, the GAP (GTPase Activating Protein) of Rho1,Citation5 which then activates the Rho1 GTPase. From Rho1, the signal follows two simultaneous routes: (A) One leads to a cascade of activation Pkc1-Bck1-Mkk1/Mkk2, ending in Slt2 dual phosphorylation and the downstream events concomitantly with the activation of the pathway.Citation5 (B) The second signal, which is, in fact, a signal of repression, is transmitted to Tor1 and Ras2. This repression eventually has several outputs: ribosomal gene repression (an ATP housekeeping mechanism), cAMP descent and the activation of a wide subset of genes which are potentially regulated by the dual transcription factor Msn2/Msn4.Citation6 At this point, we do not know the exact nature of the connection between Rho1-Tor1-Ras2. We can speculate, however, that two models can be proposed: Rho1 first inactivates Tor1 protein from the TORC1 complex and Tor1 transmits the signal to Ras2 inactivation in response to glucose deprivation and hydrogen peroxide treatment. In the second model, Rho1 signals simultaneously, but independently, to Tor1 and Ras2 inactivation. According to our study, both models suit the results that we present in the paper. Saccharomyces cerevisiae is a model that facilitates study of the various cross-talks regulating the different signal transduction pathways. The relationship between Tor-Ras and the PKC1 pathway suggests that the three pathways are essential for survival in response to specific stresses. The repression of ribosomal gene transcription is a general defence mechanism in stress response. One question which arose from our study was whether the transcription factor that regulates ribosomal gene expression in the signal was mediated by Mtl1. Preliminary results from our lab suggest that Sfp1 could be involved in this process and in oxidative response. Sfp1 is a transcription factor whose nuclear localization determines the transcriptional induction of both ribosomal and RiBi genes.Citation7,Citation8 Sfp1 is positively regulated by TOR and RAS activity.Citation7,Citation8 In response to several types of stress, including oxidative treatment, Sfp1 translocates to the cytoplasm, which provokes the repression of ribosomal gene transcription.Citation6 We observed that Sfp1 overexpression reduced cell viability upon hydrogen peroxide treatment, especially in mtl1 mutants. As previously reported, ribosomal gene repression did not occur under these conditions.Citation6 The observation that Sfp1 overexpression severely impaired mtl1 cell viability upon hydrogen peroxide treatment () suggests that Sfp1 could be negatively regulated by Mtl1 in response to specific types of stress. An excess of ribosomal gene transcription under glucose starvation and oxidative stress conditions would constitute a waste of ATP and would be detrimental for cell viability. This result did not exclude the existence of other regulatory proteins in our system, but more studies of this mechanism are required before we can draw any further conclusions.
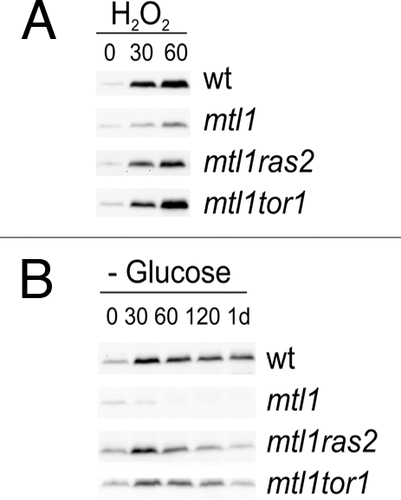
There is an information flow between different signal transduction pathways and the mechanisms that integrate information from different signal pathways in a common response.Citation9 However, how the different pathways talk is not completely understood. In a recent review, other authors have commented on CWI crosstalk with other pathways.Citation10 We described a process that connects the PKC1-MAPK pathway with TOR and RAS. In our study, the signal flowed from Mtl1, an element on the pathway to each of the other two routes: TOR and RAS. We also have data that suggest the existence of cross-talk in the opposite direction: from RAS2 and TOR1 to CWI. In both double mutants ras2mtl1 and tor1mtl1, we therefore observed an example of Slt2 phosphorylation in response to peroxide treatment and glucose depletion, which contrasted with the absence of Slt2 activation determined in the single mutant mtl1 (). The CWI activation observed in these mutants when stressed, correlated with an increase in cell viability that was similar to that determined in wild type cells.Citation6 These results demonstrate that both oxidative and nutritional stress provoked a simultaneous signalling flow from Mtl1 to several different pathways: CWI, TOR and RAS. RAS2 and TOR1 inhibition in the absence of Mtl1 also induced the activation of Slt2, the last kinase on the CWI pathway. These results indicate that the connection between the three pathways occurred at several different levels. On one hand, Mtl1 seemed to be the key regulator in response to external insults, as in the cases of oxidative and glucose depletion stress. In the absence of Mtl1, CWI was not activated and TOR1 and RAS2 were not inactivated; this severely impaired cell viability. However, according to the data shown here, another signal flowed from the inactivated RAS2 and TOR1 to activate CWI; this was independent of the cell wall receptor Mtl1 and assured that in response to certain specific types of stress, the downregulation of RAS2 and/or TOR1 guaranteed the activation of the CWI pathway. These mechanisms were necessary for cell survival and adaptation in the budding yeast system.
Figures and Tables
Figure 1 Sfp1 overexpression is toxic in the absence of Mtl1 when cells are exposed to hydrogen peroxide treatment. (A) Serial dilutions of wt, mtl1, wt+pSfp1, mt1l+pSfp1 were plated onto SD plates containing or not containing hydrogen peroxide 1 mM. Sfp1 was cloned in pUG35 plasmid containing the URA3 gene. Sfp1 ORF is under the MET25 promoter in this plasmid. (B) A temptative model depicting the possible interactions between Mtl1, TOR1, RAS2 and Sfp1.
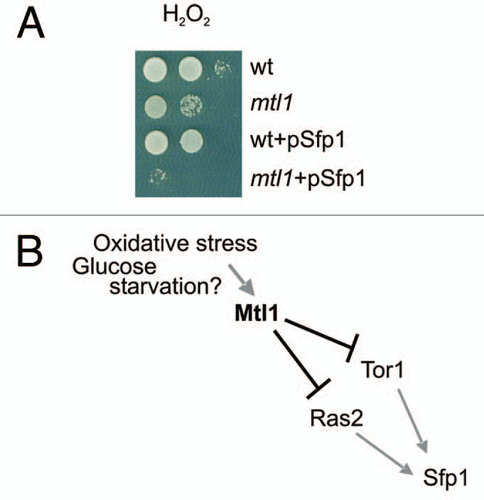
Figure 2 The simultaneous absence of Mtl1-Tor1 and Mtl1-Ras2 induces Slt2 phosphorylation in response to hydrogen peroxide treatment and in response to glucose depletion. (A) western blot analysis of Slt2 activity using the p44/42 polyclonal antibody in samples from wt, mtl1, mtl1ras2 and mtl1tor1 growing exponentially and treated with 1 mM hydrogen peroxide for the indicated times. (B) The same as in (A) but the strains were exponentially grown in SD, washed four times with minimum medium without glucose, and then transferred to SD minus glucose for incubation at 25°C. Samples were collected at the indicated times for western blot analysis. Anti-GSTSlt2 antibody was used to detect total Slt2 protein; this was equivalent in all the samples taken in the experiment (not shown for simplification).
Acknowledgements
This work was supported by Ministerio de Educacion y Ciencia (Spanish Government) Grant BFU2009-11215. N.P. was founded by a contract associated to the former Grant. M.I.P was supported by a fellowship from the Generalitat de Catalunya (Spain).
Addendum to:
References
- Ketela T, Geen R, Bussey H. Saccharomyces cerevisiae Mid2p is apontential cell wall stress sensor and upstream activator of the PKC1-MPK1 cell integrity pathway. J Bacteriol 1999; 181:3330 - 3340
- Rajavel M, Philip B, Buehrer BM, Errede B, Levin DE. Mid2 is a putative sensor for cell integrity signalling in Saccharomyces cerevisiae. Mol Cell Biol 1999; 19:3969 - 3976
- Philip B, Levin DE. Wsc1 and Mid2 are cell surface sensors for cell wall integrity signalling that act through Rom2, a guanine nucleotide exchange factor for Rho1. Mol Cell Biol 2001; 21:271 - 280
- Vilella F, Herrero E, Torres J, de la Torre-Ruiz MA. Pkc1 and the upstream elements of the cell integrity pathway in Saccharomyces cerevisiae, Rom2 and Mtl1, are required for cellular responses to oxidative stress. J Biol Chem 2005; 280:9149 - 9159
- Levin DE. Cell wall integrity signalling in Saccharomyces cerevisiae. Microbiol Mol Biol Rev 2005; 69:262 - 291
- Petkova M, Pujol-Carrion N, Arroyo J, García-Cantalejo J, de la Torre-Ruiz MA. Mtl1 is required to activate general stress response through Tor1 and Ras2 inhibition Under conditions of glucose starvation and oxidative stress. J Biol Chem 2010; 285:19521 - 19531
- Marion RM, Regev A, Segal E, Barash Y, Koller D, Friedman N, et al. Sfp1 is a stress-and nutrient-sensitive regulator of ribosomal protein gene expression. Proc Natl Acad Sci USA 2004; 101:14315 - 14322
- Jorgesen P, Rupes I, Sharon JR, Schneper L, Broach JR, Tyers M. A dynamic transcriptional network communicates growth potential to ribosome synthesis and critical cell size. Genes Dev 2004; 18:2491 - 2505
- Albert R, Oltvai ZN. Shaping specificity in signalling networks. Nat Gen 2007; 39:286 - 287
- Fuchs BB, Mylonakis E. Our paths might cross: the role of the fungal cell wall integrity pathway in stress response and cross talk with other stress response pathways. Eukaryotic Cell 2009; 8:1616 - 1625