Abstract
Group-living species produce signals that alter the behavior and even the physiology of their social partners. Social insects possess especially sophisticated chemical communication systems that govern every aspect of colony life, including the defining feature of eusociality: reproductive division of labor. Current evidence hints at the central importance of queen pheromones, but progress has been hindered by the fact that such pheromones have only been isolated in honeybees. In a pair of papers on the ant Lasius niger, we identified and investigated a queen pheromone regulating worker sterility. The cuticular hydrocarbon 3-methylhentriacontane (3-MeC31) is correlated with queen maturity and fecundity, and workers are also more likely to execute surplus queens that have low amounts of this chemical. Experiments with synthetic 3-MeC31 found that it inhibits ovarian development in queenless workers and lowers worker aggression towards objects coated with it. Production of 3-MeC31 by queens was depressed by an experimental immune challenge, and the same chemical was abundant on queen-laid eggs, suggesting that the workers’ responses to the queen are conditional on her health and fecundity. Together with other studies, these results indicate that queen pheromones are honest signals of quality that simultaneously regulate multiple social behaviors.
Parsimonious Regulation of Colony Life by Queen Pheromones
In combination with previous results, our new dataCitation1,Citation2 suggest that queen pheromones can simultaneously regulate multiple aspects of reproductive division of labor and other colony-level processes (). We found that the cuticular hydrocarbon 3-MeC31 is involved in signaling queen fertility, maturity and condition,Citation1,Citation2 regulating worker reproductionCitation2 and preventing worker aggression towards objects bearing the pheromone.Citation2 Several other functions are more tentatively supported. Our results on the execution of supernumerary queens in founding associations are consistent with selective worker aggression towards the queen(s) with the least 3-MeC31.Citation1 Workers that cannot directly identify their mother are predicted to attack queens that have produced the fewest workers, maximizing the chance that they are killing an unrelated queen,Citation1,Citation3 and 3-MeC31 is strongly correlated with queen productivity.
Queen-like chemicals have also been implicated in the aggressive response to nonpreferred reproductives, such as fertile workers and subordinate reproductives, in many other species of antsCitation4 (especially queenless antsCitation5), waspsCitation6 and bees.Citation7 Our results and those of Moore and LiebigCitation8 imply that queen-like hydrocarbons depress worker aggression, which is seemingly incongruous with reports of these same chemicals eliciting aggression in certain contexts, e.g., when expressed by fertile workers or subordinates.Citation4,Citation5 This disparity suggests either (A) queen pheromone has a dose-dependent effect on aggression, i.e., weak sources of pheromone attract aggression while strong sources repel it or (B) that there are multiple cues involved in the aggressive response, such that queen pheromone excites or inhibits aggression in combination with other stimuli. The dose-dependent hypothesis seems more parsimonious, although explicit tests are needed.
I also propose that queens pheromones may be used by queens to regulate their own reproductive output with respect to external cues, e.g., the number and developmental stage of brood and the presence of other queens competing to be the sole reproductive.Citation1 There is indirect evidence of this in L. niger: 3-MeC31 is present on queens, eggs and cocoons,Citation1,Citation2 and queens reduce their reproductive output when additional queens and brood are present.Citation1
Even more queen pheromone functions have been described in other taxa. Queen adoption,Citation9 supersedureCitation10 and replacementCitation10,Citation11 behavior in ants and honeybees is thought to be regulated by queen pheromones, and in honeybees there is extensive evidence that worker task allocation,Citation12 brain developmentCitation13 and learningCitation14 is influenced by queen pheromone exposure. Queen pheromones on the surface of eggs are likely to allow discrimination and differential rearing of eggs from different colony members, particularly queens and workers,Citation15,Citation16 but perhaps also from different queens.Citation16
Perspectives for Future Research
There is now near-unanimous support for the hypothesis that social insect queen pheromones are “honest signals” of fertility or condition, and that the worker response is not counter to their own fitness interests.Citation1,Citation2,Citation17–Citation20 I therefore suggest that researchers should focus on the ultimate explanation for this honesty. There are three principal reasons why signals should be honest:Citation18,Citation21 (1) dishonest signaling is not selected, (2) the signal is a costly “handicap”, such that only high-quality individuals benefit from investing heavily in signaling and (3) the signal is an unfakeable “index” that is inextricably linked to the trait it is signaling. As argued elsewhere,Citation18 hypothesis 1 is unlikely to be applicable to all social insects, including derived lineages where reproductive conflict is constrained.Citation22 For example, in L. niger, we expect that queens in multi-queen colonies would benefit from producing dishonestly high amounts of 3-MeC31 to avoid execution by workers. At present, almost all data on putative queen pheromones appear to be equally consistent with the handicap and index hypotheses. In L. niger, our immune challenge might have depressed reproductive physiology causing a corresponding drop in pheromone production (index) or reduced condition such that pheromone production was no longer possible (handicap).Citation2 Intriguingly, treatment with juvenile hormone reduced reproductive activity while slightly augmenting pheromone production in honeybee workersCitation23 (which appears to falsify the index hypothesis); however, in a comparable experiment in ants both reproduction and putative queen pheromones were suppressed by juvenile hormone.Citation24 Determining the underlying genetic architecture, biochemistry and/or fitness costs of pheromone production may be required to definitively discriminate between these hypotheses.
Our studyCitation2 shows how queen primer pheromones may be unambiguously identified, and I believe that it will be fruitful to isolate more in additional taxa. This will allow numerous novel questions to be addressed, e.g., how fast do queen pheromones evolve, are they predominantly single- or multi-component blends, and have similar pheromones evolved independently in phylogenetically-distant taxa? Answering these questions will provide insight into social evolution as a whole. For example, fast-evolving and multicomponent signals imply coevolution and possibly conflict.Citation17,Citation21 Convergent evolution of homologous queen pheromones would suggest that certain chemicals are particularly suited to the job: they might be particularly costly to produce (handicap hypothesis) or biochemically linked to reproduction (index hypothesis). There is tantalizing evidence that alkanes with a methyl group on the third carbon (like 3-MeC31) are also queen pheromones in other species of social insects. 3-methyl-alkanes have been correlated with fertility and/or caste in other highly-social formicine ants (Camponotus floridanusCitation25 and Formica fuscaCitation26) and in more basal ants (Myrmecia gulosa,Citation27 Diacamma ceylonese,Citation28 Pachycondyla inversaCitation29 and Platythyrea punctataCitation30). Even more surprisingly, these compounds are characteristic of reproductives in the distantly-related termitesCitation31 and have been indirectly linked to the regulation of worker reproduction in the wasp Ropalidia marginata.Citation32
Figures and Tables
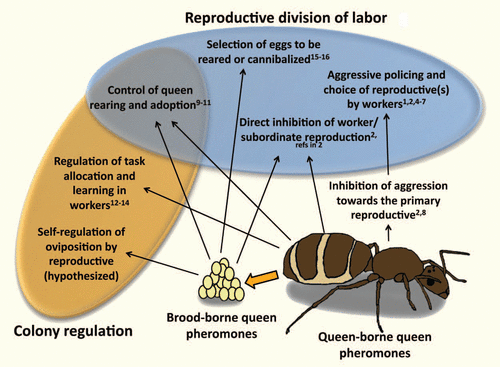
Figure 1 Known and hypothesized functions of queen pheromones produced by queens (or other reproductive individuals) and carried on their brood. The numbers give a non-exhaustive list of studies providing evidence consistent with each function.
Acknowledgements
I am grateful to all members of the Centre for Social Evolution, Copenhagen for providing a stimulating work environment. This work was supported by a Marie Curie Intra-European Fellowship (#235403; CHEMDOC).
Addendum to: and
References
- Holman L, Dreier S, d'Ettorre P. Selfish strategies and honest signalling: reproductive conflicts in ant queen associations. Proc R Soc B 2010; 277:2007 - 2015
- Holman L, Jørgensen CG, Nielsen J, d'Ettorre P. Identification of an ant queen pheromone regulating worker sterility. Proc R Soc B 2010; 277:3793 - 3800
- Balas MT. Conditions favoring queen execution in young social insect colonies. Insectes Sociaux 2005; 52:77 - 83
- Smith AA, Hölldober B, Liebig J. Cuticular hydrocarbons reliably identify cheaters and allow enforcement of altruism in a social insect. Curr Biol 2009; 19:78 - 81
- Peeters C, Liebig J. Gadau J, Fewell J. Fertility signaling as a general mechanism of regulating reproductive division of labor in ants. Organization of Insect Societies: From Genome to Socio-Complexity 2009; Harvard University Press
- Dapporto L, Romana Dani F, Turillazzi S. Social dominance molds cuticular and egg chemical blends in a paper wasp. Curr Biol 2007; 17:504 - 505
- Amsalem E, Twele R, Francke W, Hefetz A. Reproductive competition in the bumble-bee Bombus terrestris: do workers advertise sterility?. Proc R Soc B 2009; 276:1295 - 1304
- Moore D, Liebig J. Mixed messages: fertility signaling interferes with nestmate recognition in the monogynous ant Camponotus floridanus. Behav Ecol Sociobiol 2010; 64:1011 - 1018
- Vander Meer RK, Alonso LE. Queen primer pheromone affects conspecific fire ant (Solenopsis invicta) aggression. Behav Ecol Sociobiol 2002; 51:122 - 130
- Winston ML. The Biology of the Honey Bee 1987; Harvard University Press
- Edwards JP. Caste regulation in the pharaoh's ant Monomorium pharaonis: the influence of queens on the production of new sexual forms. Physiol Entomol 1987; 12:31 - 39
- Robinson GE, Winston ML, Huang Z, Pankiw T. Queen mandibular gland pheromone influences worker honey bee (Apis mellifera L.) foraging ontogeny and juvenile hormone titers. 1998; 44:685 - 692
- Beggs KT, Glendining KA, Marechal NM, Vergoz V, Nakamura I, Slessor KN, et al. Queen pheromone modulates brain dopamine function in worker honey bees. Proc Natl Acad Sci USA 2007; 104:2460 - 2464
- Vergoz V, Schreurs HA, Mercer AR. Queen pheromone blocks aversive learning in young worker bees. Science 2007; 317:384 - 386
- van Zweden JS, Heinze J, Boomsma JJ, d'Ettorre P. Ant queen egg-marking signals: matching deceptive laboratory simplicity with natural complexity. PLoS One 2009; 4:4718
- Endler A, Liebig J, Hölldobler B. Queen fertility, egg marking and colony size in the ant Camponotus floridanus. Behav Ecol Sociobiol 2006; 59:1 - 10
- Heinze J, d'Ettorre P. Honest and dishonest communication in social Hymenoptera. J Exp Biol 2009; 212:1775 - 1779
- van Zweden JS. The evolution of honest queen pheromones in insect societies. Commun Integr Biol 2010; 3:50 - 52
- Le Conte Y, Hefetz A. Primer pheromones in social hymenoptera. Annu Rev Entomol 2008; 53:523 - 542
- Keller L, Nonacs P. The role of queen pheromones in social insects: queen control or queen signal?. Anim Behav 1993; 45:787 - 794
- Maynard Smith J, Harper D. Animal Signals 2003; Oxford University Press
- Khila A, Abouheif E. Reproductive constraint is a developmental mechanism that maintains social harmony in advanced ant societies. Proc Natl Acad Sci USA 2008; 105:17884 - 17889
- Malka O, Katzav-Gozansky T, Hefetz A. Uncoupling fertility from fertility-associated pheromones in worker honeybees (Apis mellifera). J Insect Physiol 2009; 55:205 - 209
- Cuvillier-Hot V, Lenoir A, Peeters C. Reproductive monopoly enforced by sterile police workers in a queenless ant. Behav Ecol 2004; 15:970 - 975
- Endler A, Liebig J, Schmitt T, Parker JE, Jones GR, Schreier P, et al. Surface hydrocarbons of queen eggs regulate worker reproduction in a social insect. Proc Natl Acad Sci USA 2004; 101:2945 - 2950
- Hannonen M, Sledge MF, Turillazzi S, Sundström L. Queen reproduction, chemical signalling and worker behaviour in polygyne colonies of the ant Formica fusca. Anim Behav 2002; 64:477 - 485
- Dietemann V, Peeters C, Liebig J, Thivet V, Hölldobler B. Cuticular hydrocarbons mediate discrimination of reproductives and nonreproductives in the ant Myrmecia gulosa. Proc Nat Acad Sci USA 2003; 100:10341 - 10346
- Cuvillier-Hot V, Cobb M, Malosse C, Peeters C. Sex, age and ovarian activity affect cuticular hydrocarbons in Diacamma ceylonense, a queenless ant. 2001; 47:485 - 493
- Heinze J, Stengl B, Sledge MF. Worker rank, reproductive status and cuticular hydrocarbon signature in the ant, Pachycondyla cf. inversa. Behav Ecol Sociobiol 2002; 52:59 - 65
- Hartmann A, D'Ettorre P, Jones GR, Heinze J. Fertility signaling—the proximate mechanism of worker policing in a clonal ant. Naturwissenschaften 2005; 92:282 - 286
- Weil T, Hoffmann K, Kroiss J, Strohm E, Korb J. Scent of a queen—cuticular hydrocarbons specific for female reproductives in lower termites. Naturwissenschaften 2009; 96:315 - 319
- Bhadra A, Mitra A, Deshpande SA, Chandrasekhar K, Naik DG, Hefetz A, et al. Regulation of reproduction in the primitively eusocial wasp Ropalidia marginata: on the trail of the queen pheromone. J Chem Ecol 2010; 36:424 - 431