Abstract
Current discussion about reciprocal altruism is plagued by a few points of continuing disagreement/misunderstanding. In order to facilitate progress in understanding the role of reciprocity in animal societies, in this paper we try to highlight these points of disagreement/misunderstanding. Our contribution can be summarized by the following statements: 1) A temporal contingency between action and reciprocation is not the sole valid evidence for reciprocal altruism; 2) Reciprocity is not (always) cognitively demanding; 3) Kin biases in altruism are not necessarily and entirely due to kin selection; 4) Mutualism can also involve reciprocal partner choice; 5) Biological market theory is an extension of reciprocal altruism theory.
Reciprocal altruism is one of those topics that generate endless debate. We believe that part of this debate is based on a continuing misunderstanding of a few key issues. Here we focus on clarifying these issues (we will not, however, deal with semantic issues because these have been dealt with elsewhere recently, reviewed in ref. Citation1 and Citation2). The issues that we want to clarify are related to three important findings of recent research on reciprocal altruism: (1) reciprocal partner choice is widespread; (2) reciprocal partner choice is not constrained to occur over short time frames; (3) reciprocity plays a larger role than kinship in determining partner choice (reviewed in ref. Citation2 and Citation3). In the following sections, we tackle and explain five main issues with the hope of clarifying the essence of the current debate on reciprocal altruism.
A Temporal Contingency between Action and Reciprocation Is Not the Sole Valid Evidence for Reciprocal Altruism
A short-term contingent temporal relation between action and reciprocation in conceptually isolated dyads is generally considered as the hallmark of reciprocal altruism. This is possibly a consequence of the preeminent role played by theoretical models based on the Iterated Prisoner's Dilemma that can only be tested by assessing the effect of previous cooperation/defection on subsequent cooperation/defection (i.e., a contingent temporal relation) within single, isolated dyads.Citation4
In the last 15 years, biological market theory has emphasized the role of partner choice in reciprocation.Citation5,Citation6 According to biological market theory, outbidding competition for the best partners is at the basis of the social choices of group-living animals. An often noted consequence of outbidding competition and partner choice is that the “value” of services exchanged among animals depends on the present status of the marketplace and particularly on supply/demand ratios. Another important, but less frequently highlighted, consequence of outbidding competition and partner choice is that animals are predicted to deploy their altruism preferentially towards those group members that reciprocate most, i.e., to show reciprocal partner choice.
Unfortunately, very few attempts have been made both theoretically and experimentally to assess the importance of reciprocal partner choice. Theoretical models of the evolution of altruism by reciprocal partner choice are rare, possibly because of the difficulties of handling payoff matrices involving multiple individuals.Citation7,Citation8 Similarly, experimental tests of contingent partner choice in relation to altruism received are equally rare.Citation9 Nevertheless, a vast amount of correlational findings shows that reciprocal partner choice is widespread among primates and possibly beyond (reviewed in ref. Citation2). These correlational findings constitute evidence consistent with contingent partner choice and, although obviously less adept at demonstrating cause-effect relations, are so abundant that cannot be dismissed. At the same time, evidence is accumulating that animals (at least, primates) are largely indifferent to single events of altruism received in the recent past, and make instead their social choices on the basis of the long-term accumulation of such events.Citation10–Citation13 It is clear that the theoretical and empirical efforts at demonstrating the importance of short-term temporal contingencies between giving and receiving have been misdirected, and that a greater priority should be given to exploring the role of contingent partner choice based on long-term accounts of benefits given and received.
Reciprocity Is Not (Always) Cognitively Demanding
The widespread view that reciprocation is too cognitively demanding is based on the unfounded assumption that all reciprocity is cognitively based. Until recently, this assumption has been mostly “implicit”, but in the last few years, thanks to the work of Stevens and colleagues,Citation14,Citation15 hypotheses about the cognitive implications of reciprocity have been made explicit. These authors have argued that reciprocity is so cognitively complex that it cannot possibly evolve in (cognitively limited) animals. In so doing, they are in fact assuming that all reciprocity is “calculated” (i.e., based on an explicit understanding of the present costs and future benefits associated to the exchange of altruistic acts) and are therefore ignoring the possibility of emotionally based mental accounting.
We have argued elsewhereCitation16 cognitively based “calculated” reciprocity is proximately motivated by the expectation of a future benefit, thus requiring some form of planning. In contrast, emotionally based reciprocity is proximately motivated by the emotional bond that develops as a consequence of altruism received,Citation2 and does not thus require planning or other complex cognition.
A consequence of the assumption that all reciprocity is cognitively demanding is that it gives rise to a paradoxical contrast between the strong evidence that calculated reciprocity is beyond the cognitive capabilities of animals,Citation14,Citation15 and the equally strong evidence that reciprocity is in fact common.Citation2 The solution to this apparent paradox, we believe, is that most reciprocity is not calculated but emotionally based. For example, a recent study has shown that mandrills did not groom the alpha male of their group just before engaging in aggression, although by doing so they would have benefited from his greater willingness to support recent groomers. These results suggested mandrills do not plan their social interactions and that the expectation of reciprocation does not motivate them to groom.Citation17
Kin Biases in Altruism Are Not Necessarily and Entirely Dueto Kin Selection
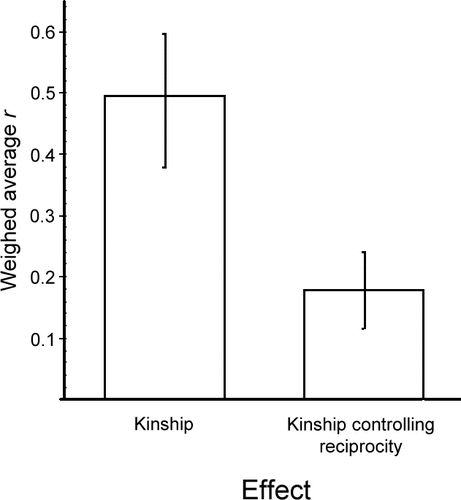
Kin biases in altruism are widespread and well known.Citation18 Nevertheless, it appears that in testing for the roles of kinship and reciprocity in shaping social preferences, researchers have been adopting a double standard. When testing for reciprocity, kinship is always controlled for (statistically or otherwise), while when testing for kinship, reciprocity is generally ignored. In a first test that compared quantitatively the roles of kinship and reciprocity in explaining primate altruism using data on 25 groups representing 14 different species, we found that controlling for reciprocity causes a large decrease in the proportion of variance in the distribution of grooming that is explained by kinship3 (). Much of the observed kin bias seems thus to be mediated by reciprocity.Citation19
Mutualism Can Also Involve Reciprocal Partner Choice
It is often assumed reciprocity plays no role in the evolution of behaviors involving benefits for both the actor and the recipient (i.e., mutualistic behaviors).Citation20 In this respect, the exclusive focus on assessing whether the benefits of social interaction (in the conceptually isolated dyad) are the inevitable consequence of one own's action or are flexibly returned by the partner led to a lack of appreciation of the role of reciprocal partner choice. In fact, both within-dyad exchanges (being them immediate/inevitable or delayed/flexible) and contingent reciprocal partner choice can contribute to fitness maximization. Therefore, we argue that whenever delayed (reciprocated) benefits can be added to the immediate/inevitable benefits of a mutualistic behavior, natural selection favors the selective deployment of this behavior among group members (i.e., partner choice) in relation to the extent of reciprocation. This is a mechanism we have called reciprocal mutualism.Citation2 For example, a female will select male sexual partners in relation to the extent of subsequent received benefits (e.g., in terms of agonistic support) even if mating already has obvious immediate benefits.
Biological Market Theory Is an Extension of Reciprocal Altruism Theory
The relation between biological market theory and reciprocal altruism theory is unclear. Two related sources of confusion are terminology and the time frame of the costs and benefits involved. Proponents of biological market theory emphasize short-term exchanges benefiting both partners and thus prefer using the term “cooperation” over “altruism“.Citation4 However, what proponents of biological market theory call goods and services are in fact behaviors having costs and benefits. Even when the exchange is very short term an altruistic behavior (having its own cost) is exchanged with another behavior (providing a benefit to the recipient). The net result of the complete interaction (giving and receiving) is that both partners benefit from the transaction, and this is exactly what is also predicted by reciprocal altruism theory.
The key difference between biological market theory and “classical” reciprocal altruism theory is not, therefore, in the timing of the costs and benefits involved, as these are essentially inconsequential (for example, one of the best experimental evidence to date of a market effect did not involve immediate exchangesCitation21). The key difference is that only biological market theory allows for interacting individuals to choose among several potential partners and thus introduces outbidding competition. As already noted, two consequences follow: reciprocal partner choice and fluctuating exchange rates in relation to the supply/demand ratio. Biological market theory is thus, in our opinion, best conceived as an extension of reciprocal altruism theory that highlights the role of partner choice and explains the existence of skewed benefits.
Conclusions
Recent reviews on reciprocal altruism have often come to sharply different conclusions (e.g., compare ref. Citation22 and ref. Citation2). We believe much of this controversy depends on a few critical misunderstandings, and have tried to highlight them. A full clarification of these controversial points will be needed in order to allow progressing towards a consensus about the role of reciprocity in the social life of animals.
Figures and Tables
Figure 1 Weighed average effect size of the effect of kinship on grooming, based on a meta-analysis of grooming distribution in 25 primate groups. The figure compares the effect of kinship on grooming when reciprocity is or is not controlled for. Data are from reference Citation3.
Acknowledgements
We thank Frantisek Baluska for inviting us to write this paper.
Addendum to:
References
- West SA, Griffin AS, Gardner A. Social semantics: altruism, cooperation, mutualism, strong reciprocity and group selection. J Evol Biol 2007; 20:415 - 432
- Schino G, Aureli F. Reciprocal altruism in primates: partner choice, cognition and emotions. Adv Study Behav 2009; 39:45 - 69
- Schino G, Aureli F. The relative roles of kinship and reciprocity in explaining primate altruism. Ecol Letters 2010; 13:45 - 50
- Noë R. Cooperation experiments: coordination through communication versus acting apart together. Anim Behav 2006; 71:1 - 18
- Noë R, Hammerstein P. Biological markets: supply and demand determine the effect of partner choice in cooperation, mutualism and mating. Behav Ecol Sociobiol 1994; 35:1 - 11
- Noë R, Hammerstein P. Biological markets. Trends Ecol Evol 1995; 10:336 - 339
- Roberts G. Competitive altruism: from reciprocity to the handicap principle. Proc R Soc B 1998; 265:427 - 431
- Sherratt TN, Roberts G. The evolution of generosity and choosiness in cooperative exchanges. J Theor Biol 1998; 193:167 - 177
- Krams I, Krama T, Igaune K, Mänd R. Experimental evidence of reciprocal altruism in the pied flycatcher. Behav Ecol Sociobiol 2007; 62:599 - 605
- Gomes CM, Boesch C. Wild chimpanzees exchange meat for sex on a long-term basis. PLoS ONE 2009; 4:5116
- Schino G, Pellegrini B. Grooming in mandrills and the time frame of reciprocal partner choice. Am J Primatol 2009; 71:884 - 888
- Schino G, Di Giuseppe F, Visalberghi E. The time frame of partner choice in the grooming reciprocation of Cebus apella. Ethology 2009; 115:70 - 76
- Schino G, Polizzi di Sorrentino E, Tiddi B. Grooming and coalitions in Japanese macaques (Macaca fuscata): partner choice and the time frame of reciprocation. J Comp Psychol 2007; 121:181 - 188
- Stevens JR, Hauser MD. Why be nice? Psychological constraints on the evolution of cooperation. Trends Cognitive Sci 2004; 8:60 - 65
- Stevens JR, Cushman FA, Hauser MD. Evolving the psychological mechanisms for cooperation. Ann Rev Ecol Evol Syst 2005; 36:499 - 518
- Schino G, Aureli F. Primate reciprocity and its cognitive requirements. Evol Anthropol 2010; 19:130 - 135
- Schino G, Pellegrini B. Grooming and the expectation of reciprocation in mandrills. Int J Primatol 2010; In press
- Chapais B, Berman CM. Kinship and behavior in primates 2004; Oxford Oxford University Press
- Chapais B. Kappeler PM, van Schaik CP. Kinship, competence and cooperation in primates. Cooperation in primates and humans: mechanisms and evolution 2006; Berlin Springer 47 - 64
- Clutton-Brock TH. Cooperation between non-kin in animal societies. Nature 2009; 462:51 - 57
- Fruteau C, Voelkl B, van Damme E, Noë R. Supply and demand determine the market value of food providers in wild vervet monkeys. Proc Nat Acad Sci USA 2009; 106:12007 - 12012
- Hammerstein P. Hammerstein P. Why is reciprocity so rare in social animals? A protestant appeal. Genetic and cultural evolution of cooperation 2003; Cambridge MIT Press 83 - 94