Abstract
Tactile working memory was found to be more developed in completely blind (congenital and acquired) than in semi-sighted subjects, indicating that experience plays a crucial role in shaping working memory. A model of working memory, adapted from the classical model proposed by Baddeley and Hitch (1) and Baddeley (2), is presented where the connection strengths of a highly cross-modal network are altered through experience.
Working memory (WM) is a cognitive system with limited capacity, involved in short term integration, disposal and retrieval of information. While there are several models of WM (reviewed in ref. Citation3), the original model of Baddeley and HitchCitation1 remains the standard reference. The tenets of this model is that WM functions evolve from the operation of specialized slave systems that act as buffers for the active manipulation and storage of information and that prefrontal cortex is a critical brain substrate for these systems.Citation4 The model consists of three separate but interconnected subsystems, a central executive with limited attentional capacity and reciprocal links to a verbal/phonological loop and a visuo-spatial sketch pad. Verbal input is actively represented in a phonological store, with representations being maintained by subvocal rehearsal in the phonological loop which, when blocked, impairs recall of verbal information. The non-verbal system or sketch pad is responsible for storing visuospatial information and creating and manipulating mental images. However, this latter system has had much less attention directed towards it and is less well understood than the verbal system.Citation5 More recently, there has been an elaboration on the original model with the addition of a fourth component, an episodic buffer.Citation6 Both versions of this WM model have been used in cognitive and neuroscience studies and commentaries. Baddeley,Citation2 reviewing the research motivated by the standard WM model, underlines the need for investigating complementary aspects of the WM system such as how learning may shape WM.
The WM model as currently conceived thus accentuates the role of language processing and visual imagery in the short-term retention of information; however, an apparent limitation is that it has no space for the storage of nonverbal input such as olfactory or tactile stimuli. This limitation is important in light of the current research, focusing specifically on WM concerned with the processing of stimuli in the tactile modality in individuals deprived of vision.
Research in blind individuals over the last decade has clearly shown that there is a large potential for functional reorganization and plasticity in the CNS. The plasticity of the prefrontal and hippocampal synapses have been linked to learning possibilities within the WM system.Citation7 There is also an increase in activity in prefrontal association cortex and parietal lobe associated with improved working memory after training.Citation8 These studies and others (reviewed in ref. Citation9) point to the plasticity of the working memory system, an increase in the capacity of the system as long as the input conforms to the described architecture of the model, i.e., verbal or visual.
It has recently been found that congenitally blind and early blind subjects performed better than late-blind subjects on several auditory perception tasks.Citation10 Early blind subjects are also better than sighted individuals in judging the direction of pitch change,Citation11 in discriminating haptic anglesCitation12 and in monaural sound localization.Citation13 Taken together, these studies and others point to the plasticity of the sensory systems (sensory and functional distribution) when the organism is confronted with the loss of a modality, especially if the loss occurs early in life. However, these studies do not inform us about the plasticity of the working memory system, only about the intriguing possibility of plasticity of one of the components, i.e., the visuo-spatial scratch pad that might be replaced by another sensory modality. A study reporting that blind subjects, regardless of whether they were early- or late-blind, performed better than normally sighted subjects on a blind route learning task provides partial support for this hypothesis.Citation14 Working memory, the sequential recording and recall of tactile (touching the walls) and proprioceptive cues are necessary for the task. More direct evidence of a reorganization of the working memory system comes from a recent studyCitation15 in which the authors found that congenitally blind (CB) participants performed better than subjects with residual vision (BRV) but equally fluent in Braille reading on a tactile working memory task.
More specifically, it was found that when the task was designed to tax the working memory for Braille stimuli, CB subjects performed better than those with residual vision (BRV), indicating that working memory for tactile stimuli is more efficient in CB individuals. Other CB and BRV subjects were also asked to recall sequences of Braille consonants with mental arithmetic problems and a mental displacement of blocks as blocking tasks. The mental displacement task was more affected by concurrently carrying out the Braille block displacement task, which indicates that tactile shortterm memory for Braille is more spatial than verbal in nature. Finally, we report here that there was a direct relationship between the scores of CB subjects on the auditory digit span task and on the tactile task with articulatory suppression. The increases or decreases in the scores on the digit span task correlated with increases or decreases on the tactile task for CB (r = 0.807, p = 0.003), but not for BRV subjects (r = −0.04, p = 0.906). There were no differences between the performance of the CB group on the tactile task, and the performance of the sighted and the BRV groups on the visual task. These findings indicate that it is likely that CB subjects have a short-term memory span specific for tactile stimuli, equivalent in capacity to the short-term memory span for visual stimuli possessed by subjects with partial or full sight. The results strongly suggest that certain aspects of the classical model should be reformulated, taking into consideration the influence of early experience in shaping the slave components in working memory.
Most people have good auditory and visual working memory, these being the primary sensory modalities and the most specialized in the human brain. This would explain why two of the components in Baddeley and Hitch'sCitation2 working memory model are a visuo-spatial sketch pad and a phonological loop: they correspond to the areas of working memory which are usually the most developed. However, the present findings suggest that the tactile modality—and perhaps the other sensory modalities should be added to the model. If the tactile patterns are coded spatially as in shapes, then memory span for tactile patterns should be the same as that in the visual modality.Citation16 In support of this contention, we found that memory span for Braille consonants was comparable to that for visually presented consonants in individuals with residual vision. Further support for this parallel between tactile and visual stimuli comes from studies that show that the neural processing of tactile information is effected by the same cortical areas involved in visuospatial processing.Citation17
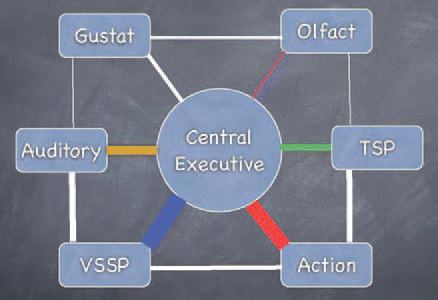
In light of the above, we propose a WM model where the network and connection strengths are altered by experience (see ). This modified model would account for our findings that individuals with residual or normal vision performed similarly on the visual task. The participants with partial vision still experience visual input maintaining the connection strength to the visual module. It is interesting to note that participants with normal vision performed slightly better than those with residual vision here, possibly because the latter do not experience quite as much visual input, and the connection strengths would not be as strong.
This model of WM takes into account the crucial role of experience in shaping WM. If a subject receives little input in one modality, then there will be weak connections so that WM for stimuli presented in that modality is poor. The opposite would be true for stimuli presented in a frequently experienced modality. The question remains, however, as to whether there is a limit to the strength of connections. If such a limit does exist, then in order for a connection to strengthen in one area, another one would have to weaken to maintain the balance. This would be in line with MillerCitation18 who found that people have a limited working memory capacity, only being able to remember seven items plus or minus two. However, if working memory is organized in line with the model proposed here, then it is likely that more stimuli can be remembered over the short-term if they are presented in a variety of different modalities (for example four visual, four auditory and four tactile). This might suggest that rather than there being a general limit on working memory, there is a limit to the amount of information which can be retained in storage modules for the different modalities. Thus, strength of connection can vary, such that an individual with much experience of auditory stimuli may remember significantly more auditory than tactile stimuli, for example, of which they have very little experience.
In short, this network is highly cross-modal with numerous interconnections, the strengths of which are altered through experience. It can be predicted that when one area is active, it sends activation to the other areas to which it is connected, depending on the connection strengths. This would account for Deibert et al.'s finding that there was activation in the visual system as well as in the somatosensory and motor systems of sighted subjects, during a tactile object recognition task. Such activation has, however, not been found for blind subjects and it may be possible that this is because they do not have an active visuo-spatial sketch pad. In individuals with no visual experience, the connection strengths to the visuo-spatial sketch pad would be minimal, if any. However, Melzer et al. did find such an activation in the visual system (occipital, temporal and parietal) and somatosensory system in blind subjects with and without early visual experience, but only for Braille reading and not for object recognition. In the case of Braille reading, it is likely that the visual system pathway is taken over by the somatosensory system that is then tightly linked to the classical visual pathways for verbal tasks (reviewed in ref. Citation21).
Conclusion
Tactile working memory was found to be more developed in blind (congenital and acquired) than in semi-sighted or sighted subjects, indicating that experience plays a crucial role in shaping working memory. In line with this finding, a model of working memory is proposed where the connection strengths of a highly cross-modal network are altered through experience. This model contains a tactuo-spatial module rather than one which is purely spatial, because the Braille task completed by blind subjects was found to involve a spatial component indicating that blind subjects retain some spatial abilities despite an absence of visual input.
Figures and Tables
Figure 1 Model of experience-based multisensory working memory. All modules are linked. The thickness of the connections represents the organism's experience in a specific modality. Experience shapes the connecting patterns in the network. Additional modules (e.g., musical) can evolve and complement an individual's working memory complex. VSSP, visuo-spatial sketch pad; TSP, Tactuo-spatial pad; Gustat, Gustatory module; Olfact, Olfactory module.
Addendum to:
References
- Baddeley AD, Hitch GJ. Bower GA. Working memory. Recent Advances in Learning and Motivation 1975; New York Academic Press 47 - 89
- Baddeley AD. Working Memory: looking Back and looking Forward. Nat Rev Neurosci 2003; 4:829 - 839
- Miyake A, Shah P. Models of working memory: Mechanisms of active maintenance and executive control 1999; New York Cambridge University Press
- Postle BR. Working memory as an emergent property of the mind and brain. Neurosc 2006; 139:23 - 38
- Becker JT, Morris RG. Working memory(s). Br Cognit 1999; 41:1 - 8
- Baddeley AD. The episodic buffer: a new component of working memory?. Trends Cognit Sci 2000; 4:417 - 423
- Laroche S, Davis S, Jay TM. Plasticity at hippocampal to prefrontal cortex synapses: Dual roles in working memory and consolidation. Hippocampus 2000; 10:438 - 446
- Olesen PJ, Westerberg H, Klingberg T. Increased prefrontal and parietal activity after training of working memory. Nat Neurosc 2004; 7:75 - 79
- Klinberg T, Forssberg H, Westerberg H. Training of Working memory in Children With ADHD. J Clin Exper Neuropsych 2002; 24:781 - 791
- Wan CY, Wood AG, Reutens DC, Wilson SJ. Early but not late-blindness leads to enhanced auditory perception. Neuropsychologia 2010; 48:344 - 348
- Gougoux F, Lepore F, Lassonde M, Voss P, Zatorre RJ, Belin P. Pitch Discrimination in the Early Blind. Nat 2004; 430:309
- Alary F, Goldstein R, Duquette M, Chapman C, Voss P, Lepore F. Tactile acuity in the blind: a psychophysical study using a two-dimensional angle discrimination task. Exper Br Res 2008; 187:587 - 594
- Gougoux F, Zatorre RJ, Lassonde M, Voss P, Lepore F. A functional neuroimaging study of sound localization: visual cortex activity predicts performance in early-blind individuals. PLoS Biol 2005; 3:324 - 333
- Fortin M, Voss P, Lord C, Lassonde M, Pruessner J, Saint-Amour D, et al. Wayfinding in the blind: larger hippocampal volume and supranormal spatial naviation. Brain 2008; 131:2995 - 3005
- Cohen H, Voss P, Lepore F, Scherzer P. The nature of working memory for braille. PLoS ONE 2010; 5:10833; http://dx.doi.org/10.1371/journal.pone.0010833
- Millar S. Memory in touch. Psicothema 1999; 11:747 - 767
- Ricciardi E, Bonino D, Gentili C, Sani L, Pietrini P, et al. Neural correlates of spatial working memory in humans: a functional magnetic resonance imaging study comparing visual and tactile processes. Neurosc 2006; 139:339 - 349
- Miller GA. The magical number seven, plus or minus two: Some limits on our capacity for processing information. Psych Rev 1956; 63:81 - 97; http://dx.doi.org/10.1037/h0043158
- Deibert E, Kraut M, Kremen S, Hart J. Neural pathways in tactile object recognition. Neurology 1999; 52:1413 - 1417
- Melzer P, Morgan VL, Pickens DR, Price RR, Wall RS, Ebner FF. Cortical activation during Braille reading is influenced by early visual experience in subjects with severe visual disability: A correlational fMRI study. Hum Br Mapping 2001; 14:186 - 195
- Amedi A, Floel A, Knecht S, Zohary E, Cohen LG. Transcranial magnetic stimulation of the occipital pole interferes with verbal processing in blind subjects. Nat Neurosci 2004; 7:1266 - 1270