Abstract
Mate choice can be sensitive to social cues from neighboring individuals, e.g., animals can copy mate choice decisions. Males that are at risk of being copied by others may respond to this with reduced preference expression (‘audience effects’). We review the various pathways by which sperm competition risk affects (1) male mate copying behavior, and (2) audience effects. For example, a recent study suggests that males gather complex social information on rivals’ sexual competitiveness (sexual activity and attractiveness to females) and respond with reduced expression of mating preferences only “when it matters”, i.e. when a sexually competitive rival is present.
Sexual selection, e.g., through mate choice, is an important evolutionary driver and individuals may integrate a wide range of information in their assessment of mate quality.Citation1 Mate choice is not only based on external characteristics (morphological, behavioral or other) of potential mating partners, but also the social environment in which mating occurs plays an essential role (non-independent mate choiceCitation2–Citation4). Especially in group-living animals5 communication events—such as communicatory interactions between sending (courting) males and receiving (choosing) females—typically do not occur in privacy, but in a public domain.Citation6–Citation11 Living in a social (or communication) networkCitation11–Citation14 enables animals to eavesdrop on other individuals' mating decisions and utilize the extracted information.Citation8,Citation10,Citation15–Citation17 The most intensely studied type of social eavesdropping is mate choice copying,Citation2,Citation3,Citation18–Citation21 but also males that are at risk of being copied by others may respond to this with reduced preference expression (“audience effects”Citation14,Citation22–Citation24). In livebearing fishes (family Poeciliidae), where females often mate multiply,Citation25,Citation26 sperm competition risk (SCR) is a decisive factor affecting males' fitness (reproductive success), but its role for non-independent mate choice was largely unknown. We demonstrate that SCR plays a significant role for (1) male mate copying behavior (with males copying less when SCR is highCitation27) and (2) audience effects. For instance, SCR leads males to deceive rivals about their actual mating preferences.Citation28 Moreover, a recent studyCitation29 uncovered a surprisingly complex case of social information use: males eavesdrop on rivals' sexual competitiveness (sexual activity and attractiveness to females) and respond with reduced preference expression only “when it matters,” i.e., when being observed by a sexually competitive rival.
Sexual selection theory often assumes that mate choice decisions are based on genetically inherited internal factors (i.e., innate search images) that enable individuals to select among several potential matesCitation1 () and accordingly, an increasing body of literature provides direct (e.g., through parent-offspring comparisons)Citation30,Citation31 or indirect evidence (e.g., from the investigation of common garden reared individuals)Citation32–Citation34 for a genetic basis of mating preferences. Still, several studies exemplified that mate choice in nature is a complex process that also involves the acquisition of information from the social environmentCitation2,Citation3,Citation21,Citation35 (). This does not come as a surprise as animal behavior in general typically is a product of both innate (genetic) and environmental factors.Citation36,Citation37
Much research has been conducted on female mate choice, and various theories have been forwarded to explain the adaptive significance of female mating preferences.Citation1,Citation38–Citation40 However, it is well established that also males express mating preferences,Citation41–Citation45 especially if mating or sperm production is costlyCitation46–Citation49 and if females differ in their resource value,Citation50,Citation51 such as numbers of oocytes in the female ovary.Citation52 In modern bony fishes (Teleostei), for example, female fecundity is typically a correlate of body size, and males prefer to mate with larger, more fecund females (e.g., in two-spotted gobies, Gobiusculus flavescens,Citation53 guppies, Poecilia reticulata,Citation54 Atlantic mollies, P. mexicana,Citation44,Citation45,Citation55 and haplochromine cichlids, Astatotilapia flaviijosephi).Citation56
Male teleosts assess females' quality not only on the basis of body size, but a range of other phenotypic traits may also be relevant.Citation50,Citation57,Citation58 However, in stark contrast to female mate choice, sperm competition risk (SCR) and intensity (SCI) can affect male mate choice.Citation59–Citation62 Obviously, a male's reproductive fitness is at stake when sperm from two or more males compete for fertilization of a clutch,Citation63,Citation64 or—in internally fertilizing species—within a single female's genital tract.Citation26,Citation65,Citation66 In this review, we provide a brief overview of recent findings in the field of socially influenced (non-independent) male mate choice. We emphasize the role played by SCR for various aspects of non-independent male mate choice.
Especially in internal fertilizing species, such as livebearing fishes (family Poecilidae), where females mate multiply,Citation67 and females can store sperm for several consecutive broods,Citation26,Citation68,Citation69 sperm competition is intense. Poeciliid broods are typically sired by several males (e.g., P. reticulata;Citation26,Citation70 sailfin molly, P. latipinna;Citation71,Citation72 green swordtails, Xiphophourus helleriiCitation73), which may be due, in part, to benefits for females of mating multiply,Citation25 but also to coercive male mating behavior, i.e., forced copulations.Citation74–Citation77 Obviously, females' behavior (i.e., their past and anticipated future sexual interactions) and especially the behavior of surrounding males can have a profound effect on SCR and thus, are likely to affect males' fitness (reproductive success). It seems straightforward to predict that the SCR a male is facing while choosing a mate is likely to affect his mate choice,Citation23,Citation61,Citation62,Citation78 and males are predicted to evolve counter-strategies to reduce SCR.Citation79
Social Eavesdropping: Males Copy Other Males' Choices
Social eavesdroppers are by-standing individuals that may extract information about the quality of the observed individuals by using information of signaling interactionsCitation3,Citation16 (). In the context of mate choice the most intensely studied form of social eavesdropping is mate choice copying, which has been defined as a process during which a female's probability of choosing a given male increases if other females have previously chosen that maleCitation18 (). Most studies on mate choice copying have focused on females,Citation2,Citation21,Citation35 but also males copy other males' choices (P. latipinna;Citation80,Citation81 pipefish, Syngnathus typhle;Citation82 three-spined stickleback, Gasterosteus aculeatusCitation83).
The adaptive significance of male mate choice copying in poeciliids could be linked to reduced costs for searching a receptive female.Citation80 Poeciliid females are receptive only as virgins or for few days post partumCitation68,Citation84,85 and accordingly, only a small proportion of females in a population are receptive at a time.Citation39,Citation69 As poeciliid males need to approach females to test their receptivity by nipping at the female's genital opening,Citation86 copying other males' mate choice may allow saving considerable energetic and opportunity costs (sensuCitation80).
Nevertheless, male mate choice copying in internally fertilizing species—like livebearing fishes—remains a conundrum, as males incur increased SCR when choosing another male's previous mate. In a recent study, we therefore asked whether male Atlantic mollies (P. mexicana) would copy less under increased SCR.Citation27 We created two copying situations with different levels of SCR: a fraction of the focal males were allowed to copy from visual interactions between a stimulus female and a model male (representing low SCR) while another fraction of males could observe direct (sexual) interactions (leading to increased SCR). As predicted, males from the latter group copied less, demonstrating that P. mexicana males indeed respond to perceived SCR when copying each other's mate choice.Citation27
Audience Effects: Risk of Being Copied Affects Mate Choice
Audience effects are defined as behavioral changes induced by the presence of other (by-standing) individuals that may or may not extract information from the observed communication eventsCitation23,Citation24 (). Beside audience induced changes in signaling and courtship behavior,Citation87,Citation88 the presence of a conspecific audience also has the potential to affect males' mate choice decisions.Citation23,Citation28,Citation29,Citation62,Citation89–Citation91 Poecilia mexicana males, e.g., cease expressing mating preferences and reduce their sexual activity when another male is present,Citation89 which may be a response to avoid unintended interception of information about their mating preferences.Citation23,Citation62 On top of that, it appears that P. mexicana males deceive rivals about their mating preferences by directing their first sexual interaction (directly after they were presented with the audience male) towards the female they had rejected beforehand,Citation28,Citation91 (). This behavior was interpreted as an attempt employed by males to lead the rival away from their preferred mate, thereby exploiting mate choice copying through dishonest signaling.Citation23 Ultimately, these behaviors may thus help to reduce the level of SCR.
As SCR is clearly linked to the present motivational state of the by-stander, we asked whether P. mexicana males are able to asses and remember rivals' sexual motivation and attractiveness to females, and if they might integrate this information strategically into their mating behavior.Citation29 Male-dyads were allowed to familiarize with one another in two adjacent compartments of a tank that was separated by a transparent Plexiglas divider. One of them was kept together with a female and thus, was perceived by the other male as sexually active, while the other male was alone and thus, perceived as sexually inactive. In subsequent mate choice tests with different stimulus females, the males from each dyad served as focal and audience males, and vice versa. Focal males ceased to show mating preferences only when they had perceived their rivals as sexually active (). In addition, focal males that were observed by a sexually active rival showed a stronger behavioral response when rivals were larger and thus, more attractive to femalesCitation29 (). This suggests that male fish are indeed able to remember and strategically exploit information about rivals when performing mate choice; in essence, males respond to an audience only “when it matters.”
Altogether, the aforementioned studies exemplify the various effects of social eavesdropping on male mate choice. Males make use of socially acquired information (mate choice copying), but also males that are at risk of being copied respond to this (audience effects), which may be a strategy to reduce SCR.
Figures and Tables
Figure 1 (A) Schematic view of an idealized mate choice situation in a classical binary choice test. Mate choice is thought of as a process of mate quality assessment involving the choosing individual (C) and two potential mating partners (stimuli, S1 and S2). (B) Mate choice in a communication network involving multiple senders and receivers of information. For simplicity, most studies to date have focused on interactions between four individuals. In this example, a by-standing individual (the audience, A) may affect the focal individual's mate choice, but also interactions between S1 and S2, and between A and both stimuli are acknowledged.
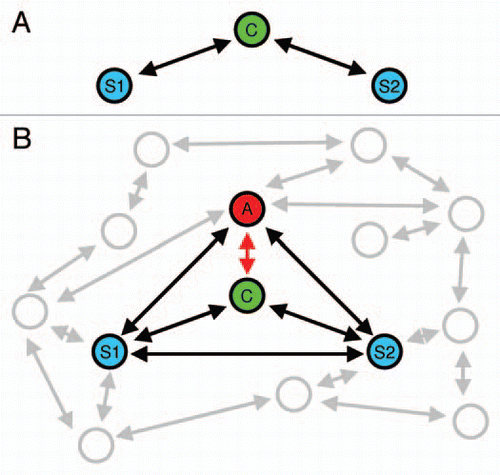
Figure 2 Different forms of social eavesdropping during mate choice in communication networks. (A) Eavesdropping on interactions between two potential mating partners: the choosing individual (C) extracts information about mate quality by observing aggressiveCitation92,Citation93 or acoustic interactionsCitation17 between two potential mating partners (S1 and S2). (B) A special form of social eavesdropping is mate choice copying, where a by-standing individual (A) extracts information about mate quality from observing sexual interactions. In the present example, A is more likely to approach S1 after having observed (C) sexually interact with S1. (C) Audience-induced changes in mate choice. The mere presence of a by-stander (the audience, (A) affects the mate choice behavior of the choosing individual (C), which may be interpreted as a strategy to reduce sperm competition risk, as A might copy C's mate choice at a later point in time.
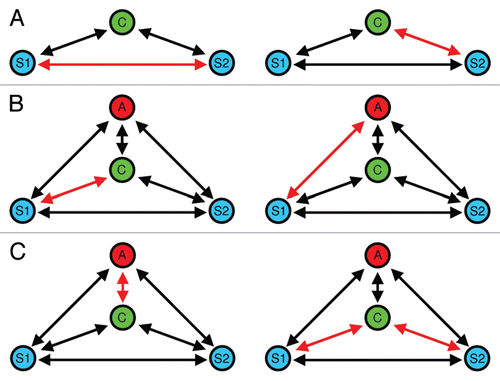
Figure 3 Deceptive signaling during male mate choice.Citation28 In (A) the choosing male (C) exhibits a mating preference for S1. (B) When an audience male (A) enters the mate choice arena, C initially interacts with S2, (C) which will be copied by (A).
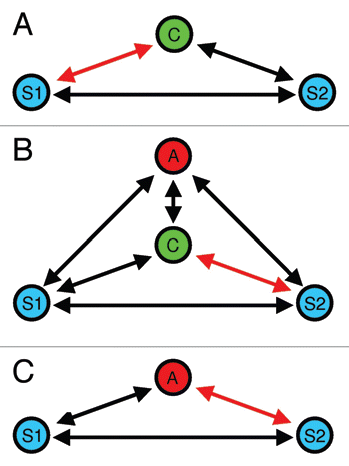
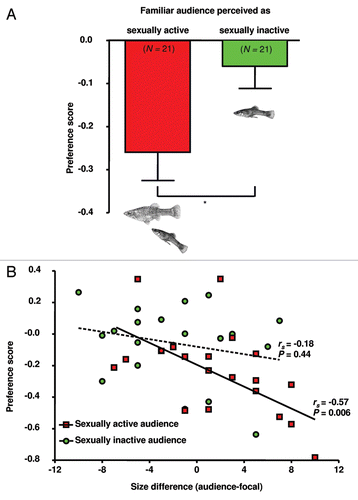
Figure 4 Changes in P. mexicana male mating preferences induced by the presence of a familiar audience male.Citation29 Males of a dyad (one sexually active and one sexually inactive male) were tested for their preferences while successively serving both as focal and audience males. (A) A comparison of preference scores (fraction of time near preferred female during 2nd-1st test parts, means ± SEM) reveals that male preferences were more affected when the rival was perceived as sexually active (paired t-test; *p < 0.05). (B) Correlation between body size differences and changes in male mating preferences (preference scores). Body size differences in the tested male dyads (N = 21) were determined as the audience male's standard length (SL)—focal male's SL. rs- and p-values are from Spearman rank order tests. The strength of male preferences decreased with increasing rival body size when the audience was perceived as sexually active, but not in the case of sexually inactive audience males.
Acknowledgements
We thank Holger Geupel for help with animal care. This work is supported by the German Research Foundation (DFG; PL 470/3-1).
References
- Andersson M. Sexual Selection 1994; Princeton Princeton University Press
- Westneat DF, Walters A, McCarthy TM, Hatch MI, Hein WK. Alternative mechanisms of non-independent mate choice. Anim Behav 2000; 59:467 - 476
- Bonnie KE, Earley RL. Expanding the scope for social information use. Anim Behav 2007; 74:171 - 181
- Earley RL. Social eavesdropping and the evolution of conditional cooperation and cheating strategies. Phil Trans R Soc B 2010; 365:2675 - 2686
- Krause J, Ruxton GD. Living in Groups 2002; Oxford Oxford University Press
- Valone TJ, Templeton JJ. Public information for the assessment of quality: a widespread social phenomenon. Phil Trans R Soc Lond B 2002; 357:1549 - 1557
- Danchin E, Giraldeau LA, Valone TJ, Wagner RH. Public information: from noisy neighbors to cultural evolution. Science 2004; 305:487 - 491
- Dabelsteen T. McGregor PK. Public, private or anonymous? Facilitating and countering eavesdropping. Animal Communication Networks 2005; Cambridge Cambridge University Press 38 - 62
- Grosenick L, Clement TS, Fernald RD. Fish can infer social rank by observation alone. Nature 2007; 445:429 - 432
- Valone TJ. From eavesdropping on performance to copying the behavior of others: a review of public information use. Behav Ecol Sociobiol 2007; 62:1 - 14
- Druen M, Dugatkin LA. Evans J, Pilastro A, Schlupp I. Communication networks. Ecology and Evolution of Livebearing Fishes (Poeciliidae) 201; Chicago University Press
- McGregor PK, Peake T. Communication networks: social environments for receiving and signaling behaviour. Acta Ethol 2000; 2:71 - 81
- Earley RL, Dugatkin LA. McGregor PK. Fighting, mating and networking: pillars of poeciliid sociality. Animal communication networks 2005; Cambridge Cambridge University Press 84 - 113
- Matos R, Schlupp I. McGregor PK. Performing in front of an audience: signallers and the social environment. Animal Communication Networks 2005; Cambridge Cambridge University Press 63 - 83
- Naguib M, Amrhein V, Kunc HP. Effects of territorial intrusions on eavesdropping neighbors: communication networks in nightingales. Behav Ecol 2004; 15:1011 - 1015
- Peake TM. McGregor PK. Eavesdropping in communication networks. Animal Communication Networks 2005; Cambridge Cambridge University Press 13 - 37
- Fitzsimmons LP, Foote JR, Ratcliffe LM, Mennill DJ. Eavesdropping and communication networks revealed through playback and an acoustic location system. Behav Ecol 2008; 19:824 - 829
- Pruett-Jones S. Independent versus non-independent mate-choice: do females copy each other?. Am Nat 1992; 140:1000 - 1009
- Dugatkin LA. Sexual selection and imitation: females copy the mate choice of others. Am Nat 1992; 139:1384 - 1389
- Nordell SE, Valone TJ. Mate choice copying as public information. Ecol Letters 1998; 1:74 - 76
- Witte K. Brown C, Laland KN, Krause J. Learning and mate choice. Fish cognition and behavior 2006; Oxford Blackwell Publishing 70 - 95
- Marler P, Dufty A, Pickert R. Vocal communication in the domestic chicken II. Is a sender sensitive to the presence and nature of a receiver?. Anim Behav 1986; 34:194 - 198
- Plath M, Schlupp I. Misleading mollies—the effect of an audience on the expression of mating preferences. Comm Integr Biol 2008; 1:199 - 203
- Zuberbühler K. Audience effects. Curr Biol 2008; 11:189 - 190
- Evans JP, Magurran AE. Multiple benefits of multiple mating in guppies. Proc Nat Ac Sci USA 2000; 97:10074 - 10076
- Becher SA, Magurran AE. Multiple mating and reproductive skew in Trinidadian guppies. Proc R Soc Lond B 2004; 271:1009 - 1014
- Bierbach D, Kronmarck C, Hennige-Schulz C, Stadler S, Plath M. Sperm competition risk affects male mate choice copying. Behav Ecol Sociobiol 2011;
- Plath M, Richter S, Tiedemann R, Schlupp I. Male fish deceive competitors about mating preferences. Curr Biol 2008; 18:1138 - 1141
- Bierbach D, Girndt A, Hamfler S, Klein M, Mücksch F, Penshorn M, et al. Male fish use prior knowledge about rivals to adjust their mate choice. Biol Lett 2011; 7:349 - 351
- Bakker TMC. The study of intersexual selection using quantitative genetics. Behaviour 1999; 136:1237 - 1265
- Pierotti MER, Knight ME, Immler S, Barson NJ, Turner GF, Seehausen O. Individual variation in male mating preferences for female coloration in a polymorphic cichlid fish. Behav Ecol 2008; 19:483 - 488
- Rodd FH, Hughes KA, Grether GF, Baril CT. A possible non-sexual origin of mate preference: are male guppies mimicking fruit?. Proc R Soc Lond B 2001; 269:475 - 481
- Endler JA, Houde AE. Geographic variation in female preferences for male traits in Poecilia reticulata. Evolution 1995; 49:456 - 468
- Brooks R, Endler JA. Female guppies agree to differ: phenotypic and genetic variation in mate-choice behaviour and the consequences for sexual selection. Evolution 2001; 55:1644 - 1655
- Dugatkin LA. Heyes CM, Galef BG Jr. Copying and mate choice. Social learning in animals: the roots of culture 1996; New York Academic Press 85 - 105
- Jennions MD, Petrie M. Variation in mate choice and mating preferences: a review of causes and consequences. Biol Rev 1997; 72:283 - 327
- Cotton S, Small J, Pomiankowski A. Sexual selection and condition-dependent mate preferences. Curr Biol 2006; 16:755 - 765
- Ryan MJ. Krebs JR, Davies NB. Sexual selection and mate choice. Behavioural ecology, an evolutionary approach 1997; Fourth edition Oxford Blackwell 179 - 202
- Houde AE. Sex, Color and Mate Choice in Guppies 1997; Princeton, New Jersey Princeton University Press
- Clutton-Brock TH, McAuliffe K. Female mate choice in mammals. Quat Rev Biol 2009; 84:1 - 27
- Bonduriansky R. The evolution of male mate choice in insects: a synthesis of ideas and evidence. Biol Rev 2001; 76:305 - 339
- Sæthers SA, Fiske P, Kalas JA. Male mate choice, sexual conflict and strategic allocation of copulations in a lekking bird. Proc R Soc Lond B 2001; 268:2097 - 2102
- Wong BBM, Jennions MD, Keogh JS. Sequential male mate choice in a fish, the Pacific blue-eye Pseudomugil signifer. Behav Ecol Sociobiol 2004; 56:253 - 256
- Plath M, Seggel U, Burmeister H, Heubel KU, Schlupp I. Choosy males from the underground: male mate choice in surface- and cave dwelling Atlantic mollies, Poecilia mexicana (Poeciliidae, Teleostei). Naturwissenschaften 2006; 93:103 - 109
- Plath M, Kromuszczynski K, Tiedemann R. Audience effect alters male but not female mating preferences. Behav Ecol Sociobiol 2009; 63:381 - 390
- Milinski M, Bakker TCM. Costs influence sequential mate choice in sticklebacks, Gasterosteus aculeatus. Proc R Soc Lond B 1992; 250:229 - 233
- Bro-Jørgensen J. Reversed sexual conflict in a promiscuous antelope. Curr Biol 2007; 17:2157 - 2161
- Kokko H, Jennions MD. Sexual conflict: The battle of the sexes reversed. Curr Biol 2008; 18:121 - 123
- Amundsen T, Forsgren E. Male mate choice selects for female coloration in a fish. Proc Natl Acad Sci USA 2001; 98:13155 - 13160
- Berglund A, Rosenqvist G. Male pipefish prefer dominant over attractive females. Behavioral Ecology 2001; 12:402 - 406
- Parker GA. Bateson P. Mate quality and mating decisions. Mate Choice 1983; Cambridge Cambridge University Press 141 - 166
- Pélabon C, Borg ÅA, Bjelvenmark J, Forsgren E, Barber I, Amundsen T. Do male two-spotted gobies prefer large fecund females?. Behav Ecol 2003; 14:787 - 792
- Herdman EJ, Kelley CD, Godin JGJ. Male mate choice in the guppy (Poecilia reticulata): Do males prefer larger females as mates?. Ethology 2004; 110:97 - 111
- Riesch R, Plath M, Schlupp I. Toxic hydrogen sulfide and dark caves: life history adaptations in a livebearing fish (Poecilia mexicana, Poeciliidae). Ecology 2010; 91:1494 - 1505
- Werner NY, Lotem A. Choosy males in a hamplochromine cichlid: first experimental evidence for male mate choice in a lekking species. Ethology 2003; 112:657 - 663
- Kolm N. Female courtship in the Banggai cardinalfish: Honest signals of egg maturity and reproductive output?. Phil Trans R Soc Lond B 2004; 361:319 - 334
- Pierotti MER, Martín-Fernández JA, Seehausen O. Mapping individual variation in male mating preference space: Multiple choice in a color polymorphic cichlid fish. Evolution 2009; 63:2372 - 2388
- Wedell N, Gage MJG, Parker GA. Sperm competition, male prudence and sperm-limited females. Trends Ecol Evol 2002; 17:313 - 320
- Aspbury AS. Sperm competition effects on sperm production and expenditure in sailfin mollies, Poecilia latipinna. Behav Ecol 2007; 18:776 - 780
- Wong BBM, McCarthy M. Prudent male mate choice under perceived sperm competition risk in the eastern mosquito fish. Behav Ecol 2009; 20:278 - 282
- Ziege M, Mahlow K, Hennige-Schulze C, Kronmarck C, Tiedemann R, Streit B, et al. Audience effects in the Atlantic molly (Poecilia mexicana)—prudent male mate choice in response to perceived sperm competition risk?. Front Zool 2009; 6:17
- Largiadér CR, Fries V, Bakker TCM. Genetic analysis of sneaking and egg-thievery in a natural population of the three-spined stickleback (Gasterosteus aculeatus L.). Heredity 2001; 86:459 - 468
- Eizaguirre C, Yeates SE, Lenz TL, Kalbe M, Milinski M. MHC-based mate choice combines good genes and maintenance of MHC polymorphism. Mol Ecol 2009; 18:3316 - 3329
- Parker GA. Sperm competition and its evolutionary consequences in insects. Biol Rev 1970; 45:525 - 567
- Constanz GD. Smith RL. Sperm competition in poeciliid fishes. Sperm competition and the evolution of animal mating systems 1984; Orlando Academic Press 465 - 485
- Haskins CP, Haskins EF, McLaughlin JJA, Hewitt RE. Polymorphism and Population Structure in Lebistes reticulatus, an Ecological Study 1961; Austin University of Texas Press
- Rosenthal HL. Observations of reproduction of the poeciliid Lebistes reticulatus (Peters). Biological Bulletin 1952; 102:30 - 38
- Magurran AE. Evolutionary Ecology: The Trinidadian Guppy 2005; Oxford Oxford University Press
- Neff BD, Pitcher TE, Ramnarine IW. Inter-population variation in multiple paternity and reproductive skew in the guppy. Mol Ecol 2008; 17:2975 - 2984
- Travis J, Trexler JC, Mulvey M. Multiple paternity and its correlates in female Poecilia latipinna (Poeciliidae). Copeia 1990; 3
- Trexler JC, Travis J, Dinep A. Variation among populations of the sailfin molly in the rate of concurrent multiple paternity and its implications for mating-system evolution. Behav Ecol Sociobiol 1997; 40:297 - 305
- Tatarenkov A, Healey CIM, Grether GF, Avise JC. Pronounced reproductive skew in a natural population of green swordtails, Xiphophorus helleri. Mol Ecol 2008; 17:4522 - 4534
- Evans JP, Pilastro A, Ramnarine IW. Sperm transfer through forced matings and its evolutionary implications in natural guppy (Poecilia reticulata) populations. Biol J Linn Soc 2003; 78:605 - 612
- Plath M, Makowicz AM, Schlupp I, Tobler M. Sexual harassment in live-bearing fishes: comparing courting and non-courting species. Behav Ecol 2007; 18:680 - 688
- Plath M. Male mating behavior and costs of sexual harassment for females in cavernicolous and extremophile populations of Atlantic mollies (Poecilia mexicana). Behaviour 2008; 145:73 - 89
- Köhler A, Hildenbrand P, Schleucher E, Riesch R, Arias-Rodriguez L, Streit B, et al. Effects of male sexual harassment on female time budgets, feeding behavior and metabolic rates in a tropical livebearing fish (Poecilia mexicana). Behav Ecol Sociobiol 2011;
- Dosen LD, Montgomerie R. Mate preferences by male guppies (Poecilia reticulata) in relation to the risk of sperm competition. Behav Ecol Sociobiol 2004; 55:266 - 271
- Bretman A, Fricke C, Chapman T. Plastic responses of male Drosophila melanogaster to the level of sperm competition increase male reproductive fitness. Proc R Soc B Biol Sci 2009; 276:1705 - 1711
- Schlupp I, Ryan MJ. Male sailfin mollies (Poecilia latipinna) copy the mate choice of other males. Behav Ecol 1997; 8:104 - 107
- Witte K, Ryan MJ. Mate-choice copying in the sailfin molly (Poecilia latipinna) in the wild. Anim Behav 2002; 63:943 - 949
- Widemo MS. Male but not female pipefish copy mate choice. Behav Ecol 2006; 17:255 - 259
- Frommen JG, Rahn AK, Schroth SH, Waltschyk N, Bakker TCM. Mate-choice copying when both sexes face high costs of reproduction. Evol Ecol 2008; 23:435 - 446
- Liley NR. Ethological isolating mechanisms in four sympatric species of Poeciliid fishes. Behav Suppl 1966; 13:1 - 197
- Parzefall J. Zur vergleichenden Ethologie verschiedener Mollienesia-Arten einschließlich einer Höhlenform von Mollienesia sphenops. Behaviour 1969; 33:1 - 37
- Fisher H, Rosenthal GG. Male swordtails court with an audience in mind. Biol Lett 2007; 3:5 - 7
- Doutrelant C, McGregor PK, Oliveira RF. The effect of an audience on intrasexual communication in male Siamese fighting fish, Betta splendens. Behav Ecol 2001; 12:283 - 286
- Plath M, Blum D, Schlupp I, Tiedemann R. Audience effect alters mating preferences in Atlantic molly (Poecilia mexicana) males. Anim Behav 2008; 75:21 - 29
- Plath M, Blum D, Tiedemann R, Schlupp I. A visual audience effect in a cavefish. Behaviour 2008; 145:931 - 947
- Plath M, Richter S, Schlupp I, Tiedemann R. Misleading mollies: surface- but not cave-dwelling Poecilia mexicana males deceive competitors about mating preferences. Acta Ethol 2010; 13:49 - 56
- Doutrelant C, McGregor PK. Eavesdropping and mate choice in female fighting fish. Behaviour 2000; 137:1655 - 1669
- Ophir AG, Galef BG. Female Japanese quail that “eavesdrop” on fighting males prefer losers to winners. Anim Behav 2003; 66:399 - 407