Abstract
Geranylgeranyltransferase I (GGT) is a protein prenyltransferase that mediates lipid modification of some proteins such as Rho family small GTPases. Since the activation of Rho GTPases mediates tumorgenesis and metastasis, GGT has become an attractive target for anti-tumor drug design. Although GGT is extensively expressed in the brain, the function of GGT in central nerves system (CNS) is totally unknown. We have previously shown that GGT was involved in neuromuscular synaptogenesis. In this study, we report that neuronal activity- and brain-derived neurotropic factor (BDNF)-dependent dendritic morphogenesis requires activation of GGT. Furthermore, GGT was activated by depolarization or BDNF in cultured neurons or in hippocampus of the mice under novelty exploration test, suggesting that neuronal activity activates GGT in vitro and in vivo. In this addendum, we further discuss the significance of this study and the possible implication to the field.
Geranylgeranyltransferase I (GGT) is a protein prenyltransferase, which catalyzes the attachment of 20 carbons long prenyl lipid anchors to the carboxyl terminal (C-terminal) of a variety of eukaryotic proteins.Citation1 It is composed of two subunits—a distinct β subunit (GGTβ) and a common α subunit (GGTα) that is shared with farnesyltransferase (FT). Substrate proteins of GGT and FT contain a C-terminal “CAAX” box, where C is an invariant cysteine residue fourth from the C terminus, A is an aliphatic amino acid, and X is any amino acid.Citation1,Citation2 Substrate proteins undergo geranylgeranylation if X is Leu or farnesylation if X is Ser, Met, Ala or Gln. Known targets of GGT include the γ subunit of brain heterotrimeric G proteins and Rho small GTPases such as Rac, RhoA and Cdc42. Substrates for FT in mammalian cells include all known Ras proteins, nuclear lamins A and B, the γ subunit of the retinal trimeric G protein transducin, rhodopsin kinase.Citation1 Because the targets of GGT and FT are tightly related to tumorgenesis and metastasis,Citation3–Citation5 GGT and FT have become very attractive targets for anti-tumor drug design.
Protein prenyltransferases are usually believed to function constitutively. However, recent studies show that FT and GGT can be regulated by multiple factors in non-neuronal cells, such as breast cancer cells or skeletal muscle cells.Citation6–Citation8 We found that motor neuron-derived glycoprotein agrin activates GGT through the muscle specific receptor tyrosine kinase MuSK, leading to acetylcholine receptor clustering at the postsynaptic membrane.Citation9 Although GGT is extensively expressed in the brain,Citation10,Citation11 the function of GGT in central nervous system (CNS) and whether it is also regulated through receptor tyrosine kinase during development of central neurons is totally unknown.
We found that the expression of GGTα or GGTβ in cultured hippocampal neurons gradually increased and peaked at approximately 8 days in vitro (DIV), a pattern that coincides with the period of extensive dendrite growth. Overexpression GGTβ, but not FTβ, promoted dendritic branching significantly in cultured hippocampal neurons compared with vector control. Treatment with GGT specific inhibitor GGTi-2147 reduced the dendritic development. The role of GGT in dendrite development was further confirmed by downregulating GGT using the approach of small interference RNA. This result indicates that GGT is sufficient and necessary for dendrite development.Citation12
Since neuronal activity and several secreted factors, including BDNF and insulin-like growth factor 1(IGF-1), are known to promote dendrite growth,Citation13–Citation18 we investigated whether these factors regulate GGT activity. We found that GGT was activated by high KCl-induced depolarization or BDNF in cultured neurons. Although insulin and IGF-1 can activate GGT in non-neuronal cells,Citation7,Citation19 neither insulin nor IGF-1 had any effect on GGT activity under our experimental conditions. In agreement with these results, high KCl treatment increased the membrane level and activity of Rac, the known substrate of GGT. Moreover, exploration of a novel environment that is known to increase neuronal activity caused activation of GGT in the mice hippocampus, suggesting that neural activity activates GGT in vivo.
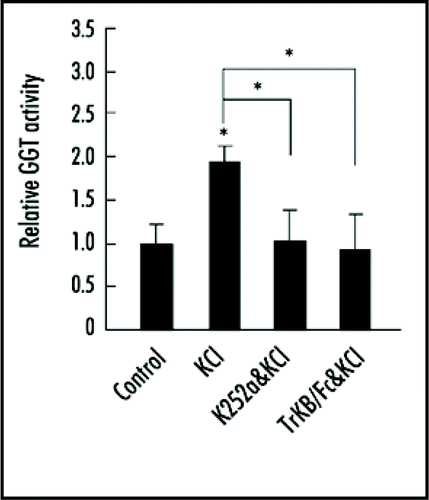
GGT is associated with muscle specific receptor tyrosine kinase MuSK at the neuromuscular junction.Citation9 Here we found that GGT was physically associated with tropomyosin-related kinase B (TrkB), the receptor for BDNF, and this association was enhanced by depolarization. When GGT-TrkB interaction was interrupted by dominant-negative GGTα mutant K164A or Y200F, which abolished GGT kinase activity, high KCl or BDNF induced dendritic arborization was suppressed. This result indicates that GGT-TrkB association is required for high KCl or BDNF induced dendritic arborization. To investigate the role of TrkB in neuronal activity-induced GGT activation, we treated hippocampal neurons with K252a, a tyrosine kinase inhibitor, or a specific inhibitor for TrkB signaling—TrkB-Fc chimera protein (extracellular domain of human TrkB fused to the C-terminal histidine-tagged Fc region of human IgG1). We found that high KCl induced GGT activation was blocked by pretreatment with K252a or TrkB-Fc (), suggesting that KCl activates GGT through TrkB.
Rac is important for dendritic development and its amino acid residue cystine in the C-terminal CAAX box is the key site for geranylgeranylation.Citation1,Citation20 We generated two mutated forms of Rac1—Rac1ΔC and Rac1C189S—by deleting the C-terminal CAAX box or substituting the cystine with serine, respectively. We found that although Rac1ΔC and Rac1C189S were unable to associate with plasma membrane, they remained the ability to interact with Tiam1, a specific Rac GEF that has been shown to mediate TrkB-mediated Rac activation.Citation21 We reasoned that these mutated forms of Rac1 might act as dominant-negatives by competing with endogenous Rac for GEFs such as Tiam1. In line with this notion, overexpression of these Rac mutants caused inhibition of high KCl-induced activation of endogenous Rac in cultured neurons. Furthermore, overexpression of these mutated forms of Rac attenuated dendrite arborization induced by GGT overexpression or treatments with KCl or BDNF. Therefore, the prenylation of Rac1 is required for depolarization- or BDNF-induced Rac activation and dendrite development.
In summary, we have identified GGT as important regulator for dendritic morphogenesis and found that GGT activity can be strictly regulated by neuronal activity or neurotrophin. Most studies on the regulatory mechanisms of Rho small GTPases focus on GTPase activating proteins (GAPs) or guanine nucleotide exchange factors (GEFs).Citation20,Citation22 For example, neuronal activity or BDNF regulates Rho small GTPases through Rho-GEF Trio or Rac1-GEF Tiam1.Citation21,Citation23 Here we identify an alternative mechanism by which neuronal activity or BDNF regulates Rac activity through activation of GGT. The prenyl moiety is derived from the mevalonate/cholesterol synthetic pathway and this pathway has been shown to affect neural plasticity or synaptogenesis.Citation24–Citation26 Since GGT is implicated in the development of neuromuscular junction and its partner TrkB is important for synapse formation by interacting with PSD95, the main scaffold protein of the postsynaptic structure,Citation27,Citation28 it shall be interesting to determine the role of protein prenylation in regulating other neuronal functions, such as synaptic plasticity.
Figures and Tables
Figure 1 KCl depolarization-induced GGT activation was blocked by K252a or TrkB/Fc. Neurons at DIV6 were pretreated with K252a or TrkB/Fc respectively for 45 min, then treated with KCl for another 45 min. After treatment, neurons were lysed and GGT activity was assayed by using Dansyl-GCVLL as substrate. Data shown are mean ± SEM of three independent experiments, with each performed in triplicate. All values were normalized to control neurons. *p < 0.05; ANOVA with Tukey test.
Acknowledgements
This work was supported by National Natural Science Foundation of China (30721004, 30825013), Key State Research Program of China (2006CB806600 and 2006CB943900), Chinese Academy of Sciences Grant (KSCX2-YW-R-102), and Program of Shanghai Subject Chief Scientist (08XD14050).
Addendum to:
References
- Zhang FL, Casey PJ. Protein prenylation: molecular mechanisms and functional consequences. Annu Rev Biochem 1996; 65:241 - 269
- Sinensky M. Recent advances in the study of prenylated proteins. Biochim Biophys Acta 2000; 1484:93 - 106
- Gelb MH, Scholten JD, Sebolt-Leopold JS. Protein prenylation: from discovery to prospects for cancer treatment. Curr Opin Chem Biol 1998; 2:40 - 48
- Sahai E, Marshall CJ. RHO-GTPases and cancer. Nat Rev Cancer 2002; 2:133 - 142
- Winter-Vann AM, Casey PJ. Post-prenylation-processing enzymes as new targets in oncogenesis. Nat Rev Cancer 2005; 5:405 - 412
- Goalstone M, Carel K, Leitner JW, Draznin B. Insulin stimulates the phosphorylation and activity of farnesyltransferase via the Ras-mitogen-activated protein kinase pathway. Endocrinology 1997; 138:5119 - 5124
- Chappell J, Golovchenko I, Wall K, Stjernholm R, Leitner JW, Goalstone M, et al. Potentiation of Rho-A-mediated lysophosphatidic acid activity by hyperinsulinemia. J Biol Chem 2000; 275:31792 - 31797
- Goalstone ML, Leitner JW, Wall K, Dolgonos L, Rother KI, Accili D, et al. Effect of insulin on farnesyltransferase. Specificity of insulin action and potentiation of nuclear effects of insulin-like growth factor-1, epidermal growth factor and platelet-derived growth factor. J Biol Chem 1998; 273:23892 - 23896
- Luo ZG, Je HS, Wang Q, Yang F, Dobbins GC, Yang ZH, et al. Implication of geranylgeranyltransferase I in synapse formation. Neuron 2003; 40:703 - 717
- Yokoyama K, Goodwin GW, Ghomashchi F, Glomset JA, Gelb MH. A protein geranylgeranyltransferase from bovine brain: implications for protein prenylation specificity. Proc Natl Acad Sci USA 1991; 88:5302 - 5306
- Ericsson J, Runquist M, Thelin A, Andersson M, Chojnacki T, Dallner G. Distribution of prenyltransferases in rat tissues. Evidence for a cytosolic all-trans-geranylgeranyl diphosphate synthase. J Biol Chem 1993; 268:832 - 838
- Zhou XP, Wu KY, Liang B, Fu XQ, Luo ZG. TrkB-mediated activation of geranylgeranyltransferase I promotes dendritic morphogenesis. Proc Natl Acad Sci USA 2008; 105:17181 - 17186
- Wayman GA, Impey S, Marks D, Saneyoshi T, Grant WF, Derkach V, et al. Activity-dependent dendritic arborization mediated by CaM-kinase I activation and enhanced CREB-dependent transcription of Wnt-2. Neuron 2006; 50:897 - 909
- Yu X, Malenka RC. Beta-catenin is critical for dendritic morphogenesis. Nat Neurosci 2003; 6:1169 - 1177
- McAllister AK, Lo DC, Katz LC. Neurotrophins regulate dendritic growth in developing visual cortex. Neuron 1995; 15:791 - 803
- McAllister AK, Katz LC, Lo DC. Opposing roles for endogenous BDNF and NT-3 in regulating cortical dendritic growth. Neuron 1997; 18:767 - 778
- Xu B, Zang K, Ruff NL, Zhang YA, McConnell SK, Stryker MP, et al. Cortical degeneration in the absence of neurotrophin signaling: dendritic retraction and neuronal loss after removal of the receptor TrkB. Neuron 2000; 26:233 - 245
- Niblock MM, Brunso-Bechtold JK, Riddle DR. Insulin-like growth factor I stimulates dendritic growth in primary somatosensory cortex. J Neurosci 2000; 20:4165 - 4176
- Goalstone ML, Leitner JW, Berhanu P, Sharma PM, Olefsky JM, Draznin B. Insulin signals to prenyltransferases via the Shc branch of intracellular signaling. J Biol Chem 2001; 276:12805 - 12812
- Van Aelst L, Cline HT. Rho GTPases and activity-dependent dendrite development. Curr Opin Neurobiol 2004; 14:297 - 304
- Miyamoto Y, Yamauchi J, Tanoue A, Wu C, Mobley WC. TrkB binds and tyrosine-phosphorylates Tiam1, leading to activation of Rac1 and induction of changes in cellular morphology. Proc Natl Acad Sci USA 2006; 103:10444 - 10449
- Hall A. Rho GTPases and the actin cytoskeleton. Science 1998; 279:509 - 514
- Estrach S, Schmidt S, Diriong S, Penna A, Blangy A, Fort P, et al. The Human Rho-GEF trio and its target GTPase RhoG are involved in the NGF pathway, leading to neurite outgrowth. Curr Biol 2002; 12:307 - 312
- Matthies H Jr, Schulz S, Hollt V, Krug M. Inhibition by compactin demonstrates a requirement of isoprenoid metabolism for long-term potentiation in rat hippocampal slices. Neuroscience 1997; 79:341 - 346
- Mauch DH, Nagler K, Schumacher S, Goritz C, Muller EC, Otto A, et al. CNS synaptogenesis promoted by glia-derived cholesterol. Science 2001; 294:1354 - 1357
- Kotti T, Head DD, McKenna CE, Russell DW. Biphasic requirement for geranylgeraniol in hippocampal long-term potentiation. Proc Natl Acad Sci USA 2008; 105:11394 - 11399
- Ji Y, Pang PT, Feng L, Lu B. Cyclic AMP controls BDNF-induced TrkB phosphorylation and dendritic spine formation in mature hippocampal neurons. Nat Neurosci 2005; 8:164 - 172
- Yoshii A, Constantine-Paton M. BDNF induces transport of PSD-95 to dendrites through PI3K-AKT signaling after NMDA receptor activation. Nat Neurosci 2007; 10:702 - 711