Abstract
Commensal microflora engages in a symbiotic relationship with their host, and plays an important role in the development of colorectal cancer (CRC). Pathogenic bacteria promote chronic intestinal inflammation and accelerate tumorigenesis. In sporadic CRC, loss of an effective epithelial barrier occurs at early stage of CRC development. As a result, non-pathogenic bacteria and/or their products infiltrate tumor stroma, drive “tumor-elicited inflammation” and promote CRC progression by activating tumor-associated myeloid and immune cells that produce IL-23 and IL-17. In this article we will summarize the recent advances in understanding the relationship between gut flora and CRC.
Commensal bacteria reside on the epithelial surfaces of the gastro-intestinal tract, lung and skin and engage in a symbiotic relationship with their host.Citation1 Commensal microflora assist host metabolism, protect from infection by pathogenic bacteria and fungi, and serve as guides and educators for development of host immune system.Citation2-Citation5 However, the seemingly happy marriage (symbiosis) between the commensal microflora and the host also carries the risk of household dispute. Dysregulated interaction between commensal and their host is a critical driving factor in the development of chronic inflammation, metabolic disorder, cardiovascular disease, and cancer.Citation2,Citation6-Citation8
The link between chronic inflammation and cancer has long been suspected since Rudolf Virchow discovered immune cell infiltration in tumors in the mid-1800s.Citation9 It is estimated that 20% of human cancers are linked to chronic inflammation and persistent infection. Examples of such association include Helicobacter pylori infection with gastric cancer, HBV and HCV infections with hepatocellular carcinoma (HCC), and chronic inflammatory bowel diseases (IBD) with colorectal cancer (CRC).Citation10-Citation12 Yet the majority of cancers that are not associated with preceding inflammation contain inflammatory infiltrates. The etiology of such “tumor-elicited inflammation,” and its role in cancer development, had remained largely elusive.
We recently reported a novel mechanism for the induction of tumor-elicited inflammation. Using a mouse model of sporadic CRC, we observed a striking loss of barrier function in transformed epithelial cells of colonic adenomas.Citation13 Loss of epithelial barrier function results in infiltration of bacterial products that activate tumor-associated macrophages to induce the production of inflammatory cytokines including IL-23 and promote tumor growth and progression. Immune and myeloid cells in the tumor stoma that produce IL-17, IL-6, IL-22, and IL-23 play key roles in this process ().Citation13
Figure 1. IL-23/IL-17 axis in tumor-elicited inflammation induced by barrier loss. Normal intestinal epithelium is covered by a mucus layer produced by goblet cells. Enterocytes also form tight junctions that control paracellular translocation of ions and molecules, and prevent translocation of commensal bacteria and their products into the lamina propria. Loss of Apc and activation of β-catenin induce adenoma formation in the intestine. Adenoma cells fail to produce mucus and form an effective intercellular junctional structure. As a consequence, gut bacteria and/or their products translocate into tumor stroma and activate tumor associated macrophages to produce IL-23, which in turn signals to Th17 cells and other IL-17 producing cells. IL-17 signals on adenoma cells and activates STAT3 indirectly to promote their proliferation.
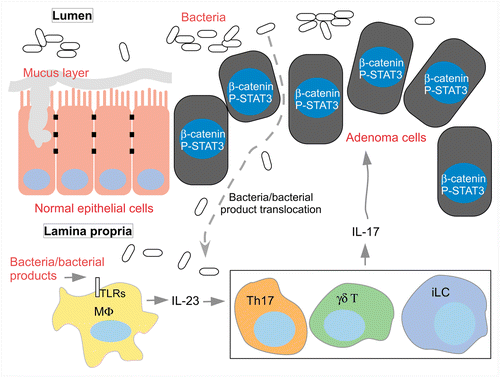
Mucosal layers and epithelial junctional structures serve as strong barriers that prevent translocation of bacteria and their products into intestinal tissue. Goblet cells produce mucus and secrete it into the lumen of intestinal tract. Highly glycosylated MUC2 protein forms a net-like polymer that spreads into layers above epithelial lining, and prevents close contact between the epithelial cells and commensal bacteria.Citation14 Mice lacking MUC2 develop spontaneous colitis and colitis-associated CRC (CAC).Citation15,Citation16 Ablation of MUC2 in ApcMin mice resulted in significantly increased intestinal tumorigenesis, especially in the colon.Citation17 Mice lacking the enzyme for mucin glycosylation also showed increased colonic permeability, and are highly susceptible to induction of colitis and CAC.Citation18 These data indicate that the mucus layer protects the host from colitis and CAC by keeping commensal microflora at a distance from the intestinal epithelium. Using a model of spontaneous CRC, we found that colonic adenomas are devoid of goblet cells that produce and secrete mucus.Citation13 MUC2 mRNA levels are significantly downregulated in both mouse and human colorectal adenomas, indicating defective mucus production and/or goblet cell differentiation already in early tumorigenesis.Citation13 The relationship between the mucus layer and CRC therefore becomes significant already at early stages of CRC development. Loss of mucin layer either before or early during CRC development serves as a driver of colonic tumorigenesis by augmenting tumor-promoting inflammation.
The intestinal epithelial barrier also depends on tight junction and adhesion junction proteins. These junctional structures define the apical and baso-lateral surface of polarized epithelial cells. Junctional structures also control paracellular traffic of ions and molecules, and prevent invasion of the epithelium by microorganisms.Citation19 Dysregulated junctional function and translocation of microbes is seen in multiple human diseases, including IBD, HIV infection, and alcohol abuse.Citation6 Ablation of JAM-A, a component of tight junction complex, resulted in enhanced colonic permeability and inflammation in mice.Citation20 Altered expression of tight junction proteins was also reported in human colorectal cancer tissues.Citation21 The change in junctional proteins was mostly speculated to be important in malignant progression of cancers. We found that the expression and localization of multiple junctional proteins, including claudins and junction–adhesion molecules (JAMs), are dysregulated in colonic adenomas compared with adjacent normal colon tissue.Citation13 Importantly, loss of barrier proteins was also observed in early human adenomas and in acutely induced aberrant foci in the mouse colon, suggesting that barrier defect is an early event during colon tumorigenesis.Citation13 Loss of the APC tumor suppressor gene and activation of β-catenin confers a proliferative state on enterocytes and blocks their differentiation.Citation22 Loss of barrier function may therefore come as a consequence of β-catenin activation and/or APC loss, in part due to loss of membrane localized, intact β-catenin, which controls cytoskeleton dynamics.Citation23
With the loss of barrier function at the adenoma surface during early CRC development, bacteria and/or their products come into close contact with transformed enterocytes and breach into the tumor stoma. Indeed, when injected into clamped mouse colon, fluorescent-labeled LPS was detected within tumors but not within adjacent normal tissue, and co-localized with the macrophage marker F4/80.Citation13 By using fluorescent in situ hybridization against bacterial 16S RNA, we also detected presence of bacteria in colorectal tumors and close to tumor epithelial cells of mouse and human early adenomas.Citation13 Infiltration of bacterial products serves as a driving force for tumor-elicited inflammation. The invading bacteria or bacterial products activate toll-like receptor (TLR)/MyD88 signaling in tumor associated macrophages, leading to the production of IL-23.
IL-23 belongs to the IL-12 family of heterodimeric cytokines that includes IL-12, IL-23, IL-27, and IL-35.Citation24,Citation25 IL-23 is composed of a unique p19 subunit, and a p40 subunit which it shares with IL-12.Citation24-Citation26 IL-23 is a major cytokine that promotes inflammation in a variety of autoimmune diseases. Ablation of the p19 subunit of IL-23 resulted in diminished experimental autoimmune encephalomyelitis (EAE) induction, similar to the effect of IL-12p40 deletion.Citation27 IL-23 was also found critical for other autoimmune diseases including rheumatoid arthritis,Citation28 psoriasis,Citation29 and IBD.Citation30-Citation32 IL-23 is upregulated in multiple human cancers and ablation of the Il23p19 gene resulted in reduced tumorigenesis in a mouse model of skin cancer.Citation33
In colonic adenomas, IL-23 signals to hematopoietic cells and upregulates the expression of other cytokines including IL-6, IL-17, and IL-22. By doing so IL-23 indirectly activates STAT3 in tumor cells and promotes tumor growth.Citation13 It is noteworthy that in human patients with stage I/II colorectal cancer, a high “Th17 signature” confers drastically reduced disease-free survival after resection of primary tumors.Citation34 Screening of early CRC cases for IL-17 expression and adjuvant treatment of patients showing a high “Th17-signature” with IL-23 or IL-17 antagonists may prove beneficial in extending disease-free survival in early stage CRC.
Over 1000 species of mostly unculturable bacteria, at a sum total of 100 trillion, reside in the human colon and form a normally symbiotic relationship with their host. However, a dysregulated relationship between the host and flora bacteria also causes multiple human diseases, including infections, obesity, IBD, and CRC.Citation1,Citation3,Citation35 Translocation of bacteria into mesenteric lymph nodes signals poor prognosis in human CRC patients.Citation36 Entertoxigenic Bacteroides fragilis (ETBF) causes inflammatory diarrhea in some humans. Colonization of the mouse intestine with ETBF triggered colitis and strongly increased tumorigenesis in the APCMin model of CRC, with a robust IL-17 response and STAT3 activation in mouse colon.Citation37 ETBF also promotes colonic tumorigenesis by inducing the production of reactive oxygen species and DNA damage.Citation38 Infection of APCMin mice with Citrobacter rodentium also resulted in elevated tumor formation in the colon.Citation39 In a model of CAC driven by loss of IL-10, chronic inflammation disturbs microbiota composition and enriches the percentage of E. coli in mice reconstituted from a germ-free facility.Citation40 Polyketide synthase (pks) genotoxic island from E. coli is more prevalent in patients with IBD and CRC, and promotes colonic tumorigenesis and malignant progression in mice.Citation40 On the contrary, delivery of Lactobacillus acidophilus that is deficient in the production of lipoteichoic acid (LTA) protected mice against colitis, and caused regression of colonic adenomas.Citation41 LTA is a component of the cell wall of several gram-positive bacteria and is recognized by TLR2 to stimulate cytokine production by dendritic cells and possibly other immune and myeloid cells.Citation42 Use of LTA-deficient L. acidophilus as probiotics ameliorated detrimental inflammation in mice and protected them from colonic tumorigenesis.Citation41 Such an effect remains to be demonstrated in humans.
The discovery of barrier breach during adenoma formation as a driving force behind “tumor-elicited inflammation” and cancer progression adds to our current knowledge of the involvement of bacteria in CRC. In this case the otherwise “benign” bacteria in our microbiota can turn into an oncogenic factor due to altered host defense mechanism, suggesting the uniqueness of the mucosal environment in prevention of cancer development. Similar mechanism may exist in cancers of other bacteria-rich epithelial surfaces, including lung, skin, and other sections of the gastrointestinal tract.
Acknowledgments
This work was supported by the Croucher Foundation and China Postdoctoral Science Foundation (20110490919) to K.W. and NIH (AI043477; DK035108) and American Association for Cancer Research (07-60-21-KARI) grants to M.K., who is an American Cancer Society Research Professor. The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIH.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
References
- Clemente JC, Ursell LK, Parfrey LW, Knight R. The impact of the gut microbiota on human health: an integrative view. Cell 2012; 148:1258 - 70; http://dx.doi.org/10.1016/j.cell.2012.01.035; PMID: 22424233
- Hill DA, Artis D. Intestinal bacteria and the regulation of immune cell homeostasis. Annu Rev Immunol 2010; 28:623 - 67; http://dx.doi.org/10.1146/annurev-immunol-030409-101330; PMID: 20192812
- Honda K, Littman DR. The microbiome in infectious disease and inflammation. Annu Rev Immunol 2012; 30:759 - 95; http://dx.doi.org/10.1146/annurev-immunol-020711-074937; PMID: 22224764
- Hooper LV, Littman DR, Macpherson AJ. Interactions between the microbiota and the immune system. Science 2012; 336:1268 - 73; http://dx.doi.org/10.1126/science.1223490; PMID: 22674334
- Littman DR, Pamer EG. Role of the commensal microbiota in normal and pathogenic host immune responses. Cell Host Microbe 2011; 10:311 - 23; http://dx.doi.org/10.1016/j.chom.2011.10.004; PMID: 22018232
- Brenchley JM, Douek DC. Microbial translocation across the GI tract. Annu Rev Immunol 2012; 30:149 - 73; http://dx.doi.org/10.1146/annurev-immunol-020711-075001; PMID: 22224779
- Ott SJ, El Mokhtari NE, Musfeldt M, Hellmig S, Freitag S, Rehman A, et al. Detection of diverse bacterial signatures in atherosclerotic lesions of patients with coronary heart disease. Circulation 2006; 113:929 - 37; http://dx.doi.org/10.1161/CIRCULATIONAHA.105.579979; PMID: 16490835
- Goldszmid RS, Trinchieri G. The price of immunity. Nat Immunol 2012; 13:932 - 8; http://dx.doi.org/10.1038/ni.2422; PMID: 22990891
- Balkwill F, Mantovani A. Inflammation and cancer: back to Virchow?. Lancet 2001; 357:539 - 45; http://dx.doi.org/10.1016/S0140-6736(00)04046-0; PMID: 11229684
- Grivennikov SI, Greten FR, Karin M. Immunity, inflammation, and cancer. Cell 2010; 140:883 - 99; http://dx.doi.org/10.1016/j.cell.2010.01.025; PMID: 20303878
- Coussens LM, Werb Z. Inflammation and cancer. Nature 2002; 420:860 - 7; http://dx.doi.org/10.1038/nature01322; PMID: 12490959
- Trinchieri G. Cancer and inflammation: an old intuition with rapidly evolving new concepts. Annu Rev Immunol 2012; 30:677 - 706; http://dx.doi.org/10.1146/annurev-immunol-020711-075008; PMID: 22224761
- Grivennikov SI, Wang K, Mucida D, Stewart CA, Schnabl B, Jauch D, et al. Adenoma-linked barrier defects and microbial products drive IL-23/IL-17-mediated tumour growth. Nature 2012; 491:254 - 8; http://dx.doi.org/10.1038/nature11465; PMID: 23034650
- Johansson ME, Larsson JM, Hansson GC. The two mucus layers of colon are organized by the MUC2 mucin, whereas the outer layer is a legislator of host-microbial interactions. Proc Natl Acad Sci U S A 2011; 108:Suppl 1 4659 - 65; http://dx.doi.org/10.1073/pnas.1006451107; PMID: 20615996
- Van der Sluis M, De Koning BA, De Bruijn AC, Velcich A, Meijerink JP, Van Goudoever JB, et al. Muc2-deficient mice spontaneously develop colitis, indicating that MUC2 is critical for colonic protection. Gastroenterology 2006; 131:117 - 29; http://dx.doi.org/10.1053/j.gastro.2006.04.020; PMID: 16831596
- Velcich A, Yang W, Heyer J, Fragale A, Nicholas C, Viani S, et al. Colorectal cancer in mice genetically deficient in the mucin Muc2. Science 2002; 295:1726 - 9; http://dx.doi.org/10.1126/science.1069094; PMID: 11872843
- Yang K, Popova NV, Yang WC, Lozonschi I, Tadesse S, Kent S, et al. Interaction of Muc2 and Apc on Wnt signaling and in intestinal tumorigenesis: potential role of chronic inflammation. Cancer Res 2008; 68:7313 - 22; http://dx.doi.org/10.1158/0008-5472.CAN-08-0598; PMID: 18794118
- An G, Wei B, Xia B, McDaniel JM, Ju T, Cummings RD, et al. Increased susceptibility to colitis and colorectal tumors in mice lacking core 3-derived O-glycans. J Exp Med 2007; 204:1417 - 29; http://dx.doi.org/10.1084/jem.20061929; PMID: 17517967
- Marchiando AM, Graham WV, Turner JR. Epithelial barriers in homeostasis and disease. Annu Rev Pathol 2010; 5:119 - 44; http://dx.doi.org/10.1146/annurev.pathol.4.110807.092135; PMID: 20078218
- Laukoetter MG, Nava P, Lee WY, Severson EA, Capaldo CT, Babbin BA, et al. JAM-A regulates permeability and inflammation in the intestine in vivo. J Exp Med 2007; 204:3067 - 76; http://dx.doi.org/10.1084/jem.20071416; PMID: 18039951
- Wang X, Tully O, Ngo B, Zitin M, Mullin JM. Epithelial tight junctional changes in colorectal cancer tissues. ScientificWorldJournal 2011; 11:826 - 41; http://dx.doi.org/10.1100/tsw.2011.86; PMID: 21479352
- Barker N, Ridgway RA, van Es JH, van de Wetering M, Begthel H, van den Born M, et al. Crypt stem cells as the cells-of-origin of intestinal cancer. Nature 2009; 457:608 - 11; http://dx.doi.org/10.1038/nature07602; PMID: 19092804
- Näthke I. Cytoskeleton out of the cupboard: colon cancer and cytoskeletal changes induced by loss of APC. Nat Rev Cancer 2006; 6:967 - 74; http://dx.doi.org/10.1038/nrc2010; PMID: 17093505
- Vignali DA, Kuchroo VK. IL-12 family cytokines: immunological playmakers. Nat Immunol 2012; 13:722 - 8; http://dx.doi.org/10.1038/ni.2366; PMID: 22814351
- Trinchieri G, Pflanz S, Kastelein RA. The IL-12 family of heterodimeric cytokines: new players in the regulation of T cell responses. Immunity 2003; 19:641 - 4; http://dx.doi.org/10.1016/S1074-7613(03)00296-6; PMID: 14614851
- Oppmann B, Lesley R, Blom B, Timans JC, Xu Y, Hunte B, et al. Novel p19 protein engages IL-12p40 to form a cytokine, IL-23, with biological activities similar as well as distinct from IL-12. Immunity 2000; 13:715 - 25; http://dx.doi.org/10.1016/S1074-7613(00)00070-4; PMID: 11114383
- Cua DJ, Sherlock J, Chen Y, Murphy CA, Joyce B, Seymour B, et al. Interleukin-23 rather than interleukin-12 is the critical cytokine for autoimmune inflammation of the brain. Nature 2003; 421:744 - 8; http://dx.doi.org/10.1038/nature01355; PMID: 12610626
- Murphy CA, Langrish CL, Chen Y, Blumenschein W, McClanahan T, Kastelein RA, et al. Divergent pro- and antiinflammatory roles for IL-23 and IL-12 in joint autoimmune inflammation. J Exp Med 2003; 198:1951 - 7; http://dx.doi.org/10.1084/jem.20030896; PMID: 14662908
- Chan JR, Blumenschein W, Murphy E, Diveu C, Wiekowski M, Abbondanzo S, et al. IL-23 stimulates epidermal hyperplasia via TNF and IL-20R2-dependent mechanisms with implications for psoriasis pathogenesis. J Exp Med 2006; 203:2577 - 87; http://dx.doi.org/10.1084/jem.20060244; PMID: 17074928
- Hue S, Ahern P, Buonocore S, Kullberg MC, Cua DJ, McKenzie BS, et al. Interleukin-23 drives innate and T cell-mediated intestinal inflammation. J Exp Med 2006; 203:2473 - 83; http://dx.doi.org/10.1084/jem.20061099; PMID: 17030949
- Kullberg MC, Jankovic D, Feng CG, Hue S, Gorelick PL, McKenzie BS, et al. IL-23 plays a key role in Helicobacter hepaticus-induced T cell-dependent colitis. J Exp Med 2006; 203:2485 - 94; http://dx.doi.org/10.1084/jem.20061082; PMID: 17030948
- Becker C, Dornhoff H, Neufert C, Fantini MC, Wirtz S, Huebner S, et al. Cutting edge: IL-23 cross-regulates IL-12 production in T cell-dependent experimental colitis. J Immunol 2006; 177:2760 - 4; PMID: 16920909
- Langowski JL, Zhang X, Wu L, Mattson JD, Chen T, Smith K, et al. IL-23 promotes tumour incidence and growth. Nature 2006; 442:461 - 5; http://dx.doi.org/10.1038/nature04808; PMID: 16688182
- Tosolini M, Kirilovsky A, Mlecnik B, Fredriksen T, Mauger S, Bindea G, et al. Clinical impact of different classes of infiltrating T cytotoxic and helper cells (Th1, th2, treg, th17) in patients with colorectal cancer. Cancer Res 2011; 71:1263 - 71; http://dx.doi.org/10.1158/0008-5472.CAN-10-2907; PMID: 21303976
- Sonnenberg GF, Artis D. Innate lymphoid cell interactions with microbiota: implications for intestinal health and disease. Immunity 2012; 37:601 - 10; http://dx.doi.org/10.1016/j.immuni.2012.10.003; PMID: 23084357
- Chin KF, Kallam R, O’Boyle C, MacFie J. Bacterial translocation may influence the long-term survival in colorectal cancer patients. Dis Colon Rectum 2007; 50:323 - 30; http://dx.doi.org/10.1007/s10350-006-0827-4; PMID: 17237910
- Wu S, Rhee KJ, Albesiano E, Rabizadeh S, Wu X, Yen HR, et al. A human colonic commensal promotes colon tumorigenesis via activation of T helper type 17 T cell responses. Nat Med 2009; 15:1016 - 22; http://dx.doi.org/10.1038/nm.2015; PMID: 19701202
- Goodwin AC, Destefano Shields CE, Wu S, Huso DL, Wu X, Murray-Stewart TR, et al. Polyamine catabolism contributes to enterotoxigenic Bacteroides fragilis-induced colon tumorigenesis. Proc Natl Acad Sci U S A 2011; 108:15354 - 9; http://dx.doi.org/10.1073/pnas.1010203108; PMID: 21876161
- Newman JV, Kosaka T, Sheppard BJ, Fox JG, Schauer DB. Bacterial infection promotes colon tumorigenesis in Apc(Min/+) mice. J Infect Dis 2001; 184:227 - 30; http://dx.doi.org/10.1086/321998; PMID: 11424022
- Arthur JC, Perez-Chanona E, Mühlbauer M, Tomkovich S, Uronis JM, Fan TJ, et al. Intestinal inflammation targets cancer-inducing activity of the microbiota. Science 2012; 338:120 - 3; http://dx.doi.org/10.1126/science.1224820; PMID: 22903521
- Khazaie K, Zadeh M, Khan MW, Bere P, Gounari F, Dennis K, et al. Abating colon cancer polyposis by Lactobacillus acidophilus deficient in lipoteichoic acid. Proc Natl Acad Sci U S A 2012; 109:10462 - 7; http://dx.doi.org/10.1073/pnas.1207230109; PMID: 22689992
- Mayer ML, Phillips CM, Townsend RA, Halperin SA, Lee SF. Differential activation of dendritic cells by Toll-like receptor agonists isolated from the Gram-positive vaccine vector Streptococcus gordonii. Scand J Immunol 2009; 69:351 - 6; http://dx.doi.org/10.1111/j.1365-3083.2009.02232.x; PMID: 19284500