Abstract
The aging field is replete with theories. Over the past years, many distinct, yet overlapping mechanisms have been proposed to explain organismal aging. These include free radicals, loss of heterochromatin, genetically programmed senescence, telomere shortening, genomic instability, nutritional intake and growth signaling, to name a few. The objective of this Point-of-View is to highlight recent progress on the “loss of heterochromatin” model of aging and to propose that epigenetic changes contributing to global heterochromatin loss may underlie the various cellular processes associated with aging.
Epigenetic Changes During Aging and the “Loss of Heterochromatin” Model
The eukaryotic genome is organized in a highly ordered chromatin structure. The nucleosome is the most fundamental unit of chromatin and consists of 146–7 base pairs of DNA wrapped around an octomer of the major core histones H2A, H2B, H3 and H4.Citation1,Citation2 Histones are subject to a multitude of post-translational covalent modifications such as methylation, acetylation, phosphorylation, ubiquitination and sumoylation at numerous residues that contribute to changes in chromatin conformation and may serve as recognition sites for chromatin regulators with chromo, tudor, WD40 repeat or Plant Homeodomain (PHD) finger domains.Citation3,Citation4 It has been established that different regions of the eukaryotic genome exist in either the relatively loose euchromatin or tightly packaged heterochromatin state. Euchromatin regions are transcriptionally active domains in contrast to the transcriptionally silent heterochromatin regions, although there are exceptions.Citation5,Citation6 Histone hypoacetylation, H3K9 hypermethylation and Heterochromatin Protein 1 (HP1) recruitment are hallmark features of heterochromatin formation.Citation7,Citation8 Heterochromatin is further divided into constitutive heterochromatin, such as in centromeric and telomeric segments that adopt the silent formation in all cell types and facultative heterochromatin as found in the inactivated X chromosome, present in only a subset of cells.Citation9-Citation11
The “loss of heterochromatin” model of aging was first put forth by Villeponteau (1997), who proposed that heterochromatin domains established early in embryogenesis are broken down during the aging process, contributing to the derepression of silenced genes and leading to aberrant gene expression patterns.Citation12 Consistent with this proposal, global heterochromatin loss has been associated with aging in humans and animals. For example, in humans, germline mutations in lamins cause Hutchinson-Gilford Progeria Syndrome (HGPS) and Atypical Werner Syndrome, diseases that mimic premature aging.Citation13 Cultured cells obtained from a female HGPS patient and expressing progerin, a truncated, mutant form of Lamin A, exhibit abnormal nuclear morphology indicative of a loss of heterochromatin.Citation13 The cells also show a decrease in the heterochromatin marker H3K9me3 and a delocalization of HP1α and the CREST antigen from constitutive heterochromatin. Consistent with the loss of pericentric heterochromatin, pericentric satellite III repeat transcription is derepressed. With regard to facultative heterochromatin, downregulation of the EZH2 methyltransferse and concomitant loss of H3K27me3 occur at the inactivated X-chromosome. Although an increase in the constitutive heterochromatic marker H4K20me3 was also observed, these results overall point to the disruption of heterochromatin in aging-related phenotypes.
These findings were substantiated in studies of natural aging in C.elegans, Drosophila, and humans. A similar change in nuclear architecture and a loss of peripheral heterochromatin were detected in nonneuronal cells during aging of C.elegans.Citation14 In Drosophila, a gradual loss of heterochromatin correlates with age and lamin proteins were found to have a role in aberrant nuclear morphology and reduced lifespan.Citation15,Citation16 In humans, skin fibroblast cell lines established from old individuals showed nuclear defects similar to those of an HGPS patient’s cells, including changes in histone modifications and increased DNA damage.Citation17 Interestingly, in cells cultured from healthy individuals, inhibiting the sporadic use of a cryptic splice site in lamin A that produces progerin reverses the nuclear defects associated with aging, further demonstrating that the global loss of heterochromatin markers and dissociation of heterochromatin proteins seen in HGPS also occur during physiological aging.Citation17
One may argue that the “loss of heterochromatin” model of aging is perhaps an over-simplification, since simultaneously with the global decrease in constitutive heterochromatin observed with aging, an increase in localized facultative heterochromatin occurs in the form of Senescence-Associated Heterochromatin Foci (SAHFs). SAHF formation is a hallmark of cellular senescence, which is associated with molecular mechanisms that contribute to irreversible cell cycle arrest, such as telomere shortening, downregulation of the canonical Wnt signaling pathway, upregulation of the cyclin dependent kinase inhibitor p16INK4a, activation of ataxia-telangiectasia mutated (ATM) kinase and p53, to name just a few key events found in late passage cultured cells and in aged animal cells, including those of primates.Citation18-Citation22 Studies have shown that SAHF formation occurs in a sequential order, in which chromosomes first decondense, followed by H3K9 methylation, binding of HP1 proteins and finally, inclusion of macroH2A; this process requires key players in heterochromatin formation including histone chaperones HIRA and Asf1, HMGA proteins, H3K9 methyl-transferases Suv39h1 and h2, and heterochromatin proteins HP1α, β and γ.Citation21-Citation26 Sites of SAHF formation include E2F target genes at which recruitment of heterochromatin-inducing and silencing proteins depends on the Rb pathway.Citation27 Moreover, the establishment of oncogene-induced SAHF requires Ataxia-telangiectasia and Rad3-related (ATR) and the transcriptional repression of E2F target genes requires ATM and p53.Citation28,Citation29 Thus, while there is a decrease in “global” heterochromatin levels, aging is also associated with an increase in localized heterochromatin formation at specific loci. This process has been referred to as “heterochromatin redistribution.”Citation26
Further support for the global loss of heterochromatin model of aging comes from the observation that DNA methylation, which is associated with heterochromatin formation, also becomes depleted globally with aging, particularly at constitutively heterochromatic, repetitive DNA sequences.Citation30-Citation33 Similarly to heterochromatin redistribution, DNA methylation in senescent cells becomes enriched at gene-specific promoter sequences, particularly those that play important roles in senescence and cancer, such as ERα, IGF-II, P14ARF, p16INK4A, Ecadherin, c-fos, and collagen-α.Citation34-Citation42 The redistribution of global DNA methylation has been attributed to the differential regulation of the Dnmt1 DNA methyltransferase that maintains DNA methylation patterns by methylating hemimethylated sites, in contrast to Dnmt3a and Dnmt3b, which are responsible for de novo DNA methylation.Citation43 In aging and immortalized fibroblasts, Dnmt1 expression and activity decrease, concomitantly with an upregulation of Dnmt3b transcription.Citation44,Citation45 DNA methylation serves as a docking site for Methyl-CpG-binding proteins (MBPs) that recruit transcriptional co-repressors for gene silencing and chromatin remodeling. Furthermore, DNMTs interact with H3K9 methyltransferases, HP1 and histone deacetylases.Citation46-Citation57 At the rDNA locus, the N-CoR repressor complex, DNA methyltransferases and histone deacetylases assemble via protein-protein interaction in a sequential manner to mediate transcriptional repression of rRNA, illustrating the intimate link between DNA methylation, H3K9 methylation and histone deacetylation culminating in heterochromatin formation.Citation58,Citation59
Changes in histone modifications have also been associated with aging. Utilizing the brains of senescence-accelerated prone mouse 8 (SAMP8) as a model for aging, decreases were found in H4K20me and H3K36me3 in the brains of aged mice, while there were increases in H3K27me3, H3K79me and H3K79me2.Citation60 In the kidneys and livers of aged rats, however, H4K20me3, a marker for repression of gene transcription, increased with aging, while H4K20me1,2 levels did not change.Citation61 H3K27me3, a marker that serves as a recognition site for the polycomb repressive complex (PRC), transcriptionally silences the INK4/ARF locus in young cells, however in aging cells the H3K27 demethylase JMJD3 is upregulated and the locus is derepressed, consistent with a senescent phenotype.Citation62 These observations are in accord with those seen in SAHF of senescent cells, as described earlier, where histone methyltransferases, demethylases and the retinoblastoma family of proteins play integral roles in epigenetic regulation of cell-cycle gene promoters. Thus, changes in epigenetic signatures during the aging process require the differential expression of specific chromatin modifiers.Citation63-Citation65
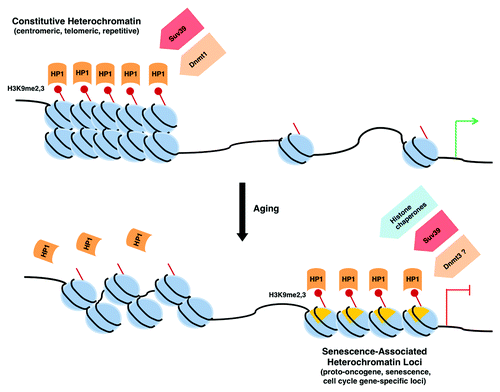
In summary, it appears that aging is associated with an overall net decrease in heterochromatin, but accompanied by an increase in heterochromatin at specific loci. The “loss of global heterochromatin” model for aging may, therefore, best be described as a net decrease in constitutive heterochromatin, with a partial redistribution of heterochromatin markers to facultative heterochromatin, resulting in the altered gene expression patterns associated with aging ().
Figure 1. Heterochromatin redistribution during aging. Aging is associated with a loss of constitutive heterochromatin, as demonstrated by a decrease in H3K9 methylation and delocalization of HP1 proteins. Concurrently, increases in facultative heterochromatin occur at specific loci, particularly at Senescence-Associated Heterochromatin Foci (SAHFs) that depend on the activity of H3K9 methyltransferases, on recruitment of HP1 proteins and histone chaperones, and on the presence of macro H2A. The decrease in activity of the maintenance DNA methyltransferase Dnmt1 and the increase in de novo Dnmt3 methyltransferases during aging may also contribute to heterochromatin redistribution.
Linking the Various Models of Aging and Associated Molecular Mechanisms: The Central Role of Heterochromatin in Aging
Recently, it has been found that overexpressing Heterochromatin Protein 1a (HP1a) in Drosophila prolongs lifespan and slows the breakdown of muscle integrity that is characteristic of old animals.Citation16,Citation66 On the other hand, loss-of-function HP1a mutant animals have a shortened lifespan. A study of epigenetic signature changes during normal aging in Drosophila showed an overall decrease in H3 and the heterochromatin marker H3K9me2 as well as HP1a delocalization, similar to the findings in other organisms.Citation13,Citation14,Citation17,Citation67 These results support the global heterochromatin loss model of aging, since modulating HP1a levels affect aging and its associated phenotypes. The complex molecular interactions associated with aging have given rise to various aging models including ROS insult, genomic instability, genetically programmed senescence, telomere shortening, nutritional intake and growth signaling. All of these models could have heterochromatin alteration as an underlying theme.
Further molecular characterization demonstrated that with the loss-of-function HP1a genotype, or a JAK gain-of-function genotype, each of which perturbs heterochromatin,Citation68,Citation69 nucleolar morphology was disrupted and formation of extrachromosomal circular (ECC) DNA increased, reflecting recombination due to rDNA instability.Citation16 Conversely, overexpressing HP1a or a non-phosphorylatable STAT, which increases heterochromatin stability,Citation69,Citation70 extended life span.Citation16 These results provide evidence for aging mechanisms mediated by heterochromatin modulation, as discussed below.
Yeast replicative aging and Sir2
While the importance of heterochromatin in nucleolar stability has been demonstrated in flies,Citation71,Citation72 the role of sirtuin histone deacetylases in rDNA stability has been studied extensively in S.cerevisiae in the context of normal aging. Sir2 deletion shortens life span in yeast and is associated with increased rDNA recombination, while providing an extra genomic copy of Sir2 extends lifespan and enhances rDNA stability.Citation73-Citation76 Moreover, it has been observed that during aging, the Sir2-containing protein complex is redistributed from telomeres to the rDNA locus, where silencing occurs by means of heterochromatin formation mediated by Sir2 and the Sir2-associated proteins, Sir3 and Sir4.Citation74,Citation77,Citation78 In line with the idea of “heterochromatin redistribution” mentioned earlier, Oberdoerffer and Sinclair (2007) propose that the redistribution of heterochromatin-associated factors is a major mechanism leading to the changes in nuclear architecture and gene expression patterns associated with aging.Citation78
Sirtuins in other organisms
Anti-aging effects of Sir2 orthologs in various other organisms including Drosophila, C. elegans, and mammals have been reported, although direct involvement in rDNA silencing during the aging process has yet to be definitively determined.Citation79,Citation80 Pharmacological inhibition of Sir2 decreases lifespan in various organisms and the expression of SIRT1, the mammalian Sir2 ortholog, decreases with age in mitotic tissues of normal aging mice and in murine and human cell culture.Citation81,Citation82 SIRT1 levels have been found to correlate with proliferating cell nuclear antigen (PCNA) levels, while being inversely correlated with senescence-activated β-galactosidase activity and p63-mediated senescence.Citation81,Citation82 These findings reinforce the idea that this epigenetic silencer has a role in organismal aging.Citation83-Citation85 However, one recent study has challenged the validity of some previous studies showing anti-aging effects of Sir2 homologs in C. elegans and Drosophila.Citation86 Thus, further work is required to determine whether the role of Sir2 orthologs in promoting longevity is generally conserved.
Caloric restriction
The heterochromatin-inducing histone deacetylase, Sir2, has been extensively studied with regard to its involvement in caloric restriction (CR), a calorie-reducing dietary regimen that has been shown to extend lifespan in yeast, worms, flies, and mammals.Citation87 One of the major mechanisms by which CR extends lifespan appears to involve Sir2/SIRT1 activation, although other mechanisms, including decreased Tor signalingCitation88 and the generation of reactive oxygen species (ROS)Citation89 have been implicated as well.Citation84,Citation85,Citation90-Citation93 When nutrients are scarce, yeast changes metabolic activity leading to altered NAD/NADH ratio, thus increasing the availability of NAD, the required catalytic cofactor of Sir2.Citation91,Citation94-Citation96 Under CR conditions in yeast, Sir2 or its homolog, HST2 suppresses rDNA recombination although this idea has sparked controversy and is still under debate.Citation97-Citation100 It has been reported that lifespan in flies can be prolonged by the Sir2 activator resveratrol or by overexpression of Sir2, and under these conditions, lifespan is not further extended by CR tand therefore it is speculated that Sir2 activation may be the main downstream effector of CR.Citation84,Citation85 The direct involvement of Sir2 in maintaining rDNA stability and preventing ECC formation, however hasnot been demonstrated in flies. In mammals, SIRT1 and heterochromatin silence rDNA transcription in response to low intracellular energy levels.Citation101 Taken together with the observations in flies that loss-of-function of HP1a produces larger flies, while overepression of HP1a leads to smaller flies,Citation16 these studies suggest that enhanced rRNA transcription resulting from a decrease in heterochromatin at the rDNA locus may increase protein synthesis and growth to accelerate aging.
CR-induced SIRT1 and HDAC1 activation can also contribute to longevity by transcriptionally downregulating genes involved in senescence including p16INK4a, hTERT, SFRP1 and E-Cadherin, by deacetylating H4K16 at their genomic loci.Citation102,Citation103 Moreover, SIRT1 mediates functional cross-talk between histone deacetylation and the heterochromatin marker H3K9me3. While deacetylating H4K16, SIRT1 simultaneously upregulates activity of the SUV39H1 methyltransferase by deacetylating a residue in its catalytic SET domain.Citation104 CR also leads to altered DNA methylation patterns. SIRT1 deacetylates the DNA methyl transferase DNMT1 to regulate its function, indicating the existence of an intricate network of epigenetic cross-talk.Citation105 Studies in rats, mice, and a cultured human cell model of CR provide examples of regulation by DNA methylation. In the cultured human cell CR model, DNA hypermethylation occurs at the E2F-1 binding sequence within the p16INK4a promoter and leads to its downregulation.Citation106 In pancreatic acinar cells of rats, CR results in increased promoter DNA methylation of the Ras proto-oncogene.Citation107 In mice, CR is able to limit the increase in Dnmt3a levels that normally occurs in aging cells.Citation108,Citation109 Thus, nutrition and growth signaling both affect aging, mediated at least in part by heterochromatin redistribution. Regulation, of the expression of genes involved in metabolism, apoptosis and cellular senescence by increased local facultative heterochromatinization may be an important mechanism by which aging is regulated.
In contrast to yeast Sir2, which appears to deacetylate only histones, mammalian SIRT1 can directly deactylate the lysines of multiple target proteins including PGC-1α, p53 and FOXO, to confer oxidative stress resistance, and the DNA repair protein, Ku70, leading to inactivation of Bax and inhibition of apoptosis.Citation73,Citation110-Citation112 These results suggest that apart from epigenetic modulation, SIRT1 can promote longevity in multiple ways, linking nutritional effects, genomic stability and heterochromatin protection.
Insulin signaling
In C. elegans, genetic analysis has demonstrated that sir-2 acts within the insulin-like receptor (IGF) pathway.Citation83 Insulin signaling and factors promoting growth and protein synthesis tend to accelerate aging, while inhibition of this pathway extends life span. Mutations in the Insulin/IGF-1-like receptor pathway have been found to extend lifespan in C. elegans, Drosophila and mice. Studies show that increased longevity resulting from decreased Insulin/IGF signaling is mediated to a great extent by FOXO and on the other hand, insulin/IGF pathway accelerates aging by signaling to TOR.Citation113-Citation118 The insulin-like receptor pathway has a large number of downstream transcriptional targets through which it may affect aging. These targets, which have been identified in various organisms, include antioxidant, chaperone, apolipoprotein, amino acid turnover and antibacterial genes, such as superoxide dismutase, metallothionine, catalase, and glutathionine S-transferase,Citation117,Citation119-Citation124 and collectively suggest that nutritional intake and growth signaling are related to ROS and DNA damage.
The insulin/IGF-1 signaling pathway is also directly linked to heterochromatin regulation during organismal aging in C.elegans, where the abnormal nuclear morphology phenotype associated with heterochromatin disruption and aging are delayed by insulin/IGF-1 signaling mutations.Citation14 Furthermore, in normally aging human cells, inhibiting the sporadic use of a splice site producing progerin, the truncated form of lamin A causing HGPS, leads to downregulation of p21, IGFBP3 and GADD45B.Citation17 Taken together, these observations demonstrate that lifespan extension by reduced growth signals is tightly coupled to the maintenance of heterochromatin domains.
Free radicals
Perhaps one of the most prominent theories of aging is the “free radical theory,” proposed by Harman in the 1950s. Free radicals can be intrinsic ROS that are normal byproducts of metabolism, or extrinsic ROS, such as those generated by radiation. ROS are sources of damage to macromolecules (they cause lipid peroxidation, protein damage, and single- and double-strand breaks, adducts, and crosslinking of DNA), and their accumulation contributes to the aging process.Citation90,Citation125 Consistently with this idea, 8-oxoguanine (oxo8dG), a major product of oxidative damage to DNA, accumulates with age, and many longevity-promoting mutations in model organisms increase resistance to oxidative stress.Citation126,Citation127 A study using human brain tissue and cultured neurons suggests that an accumulation of oxidative DNA damage at the promoters of genes involved in neural function may account for age-related cognitive decline.Citation128 However, the relationship between ROS and aging appears to be more complex than the free radical theory suggests. Reactive oxygen and nitrogen species have been found to function in normal cell signaling and in some studies on CR lifespan extension in yeast, flies, and worms, increased oxidative stress was detected in longer-lived animals.Citation90 Additionally, mice heterozygous for Sod2 have lifespans comparable to wildtype despite increased levels of nuclear and mitochondrial 8oxodG with age.Citation129 Indeed, the hormesis theory of aging argues that low-level stimulation by stress signals increases lifespan.Citation3 Nevertheless, it is evident that a sufficient accumulation of damage to macromolecules accelerates aging and the onset of aging-related pathologies. There is emerging evidence suggesting a protective role of heterochromatin in counteracting ROS-induced DNA damage, as further discussed below.
Genome instability
Genomic damage that results in genomic instability is a major inducer of the DNA damage response and p53-mediated senescence, as described earlier. Defects in the DNA repair machinery as a consequence of germline mutations in ATM and ATR, for instance, can cause phenotypes resembling premature aging.Citation130-Citation133 As with damage caused by factors such as ROS, DNA damage caused by replication errors and by spontaneous chemical changes to the DNA can also lead to genomic instability and senescence. In Drosophila, it has been demonstrated that increased levels of HP1a or unphosphorylated STAT, which augment global heterochromatin levels, provide protection against double-stranded DNA breaks induced by ionizing radiation and enhance survival.Citation134 Conversely, reducing heterochromatin levels increases sensitivity to DNA damage by ionizing radiation and increases the incidence of radiation-induced cell-cycle arrest.Citation134 It is therefore plausible to speculate that sustaining the global level of heterochromatin at relatively high levels may contribute to extended lifespan by protecting against DNA damage.
Telomere shortening
Telomere shortening is another cellular signal that induces senescence, by way of ATM, Chk2,and activation of p53.Citation135-Citation139 In S.cerevisiae, Sirtuins that are associated with telomeric heterochromatin are integral to telomere stability.Citation140-Citation142 At telomeres of telomerase deficient mice with an accelerated aging phenotype, or in late passage cultured cells,Citation143,Citation144 the heterochromatin markers H3K9me3, H4K20me3, and CBX3 diminish, while the euchromatin marker H3K9ac increases.Citation145,Citation146 On the other hand, levels of the telomere binding proteins TERF1 and TERF2 remain stable, demonstrating the specificity of epigenetic changes in this region. DNA methylation at telomeres also declines under these conditions, consistent with previous findings that mouse cells deficient for DNMTs have elongated telomeres that show increased recombination.Citation147 Thus loss of heterochromatin at telomeric regions and the resulting instability is another factor contributing to the aging process.
Cellular senescence
Senescent cells accumulate upon organismal aging and in the tissues of patients with pathologic conditions that mimic premature aging. Although senescence can occur in some cells without SAHF formation, in mouse lymphocytes, a lack of H3K9 methylation and Suv39h1 prevent Ras-induced senescence, suggesting that heterochromatin redistribution and SAHF formation may be upstream regulators of senescence.Citation65,Citation148,Citation149 Although cellular senescence and organismal aging are tightly associated, it remains to be established whether or not this link is causal. It is speculated that cellular senescence contributes to aging by limiting the numbers of progenitors and stem cells, thus restricting tissue renewal capacity, and that senescent cells secrete factors such as proteases that interfere with tissue function.Citation150-Citation152 As discussed previously, expression of progerin is associated with heterochromatin disruption and with HGPS aging phenotypes. Progerin expression also results in Notch signaling activity, which causes dysfunction of adult stem cells.Citation67 These findings suggest that maintenance of heterochromatin may promote longevity by sustaining the renewal capacity of stem cells.
Conclusions and Perspectives
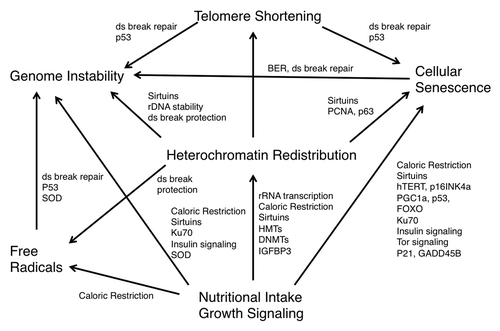
Increasing evidence suggests that the “loss of heterochromatin” model of aging proposed decades ago could perhaps be re-defined as the “global heterochromatin loss and redistribution” model, in which a total decline in constitutive heterochromatin occurs together with an increase in heterochromatin at specific loci, contributing to genomic instability and transcriptional changes. The epigenetic alterations resulting from global heterochromatin loss may be at the root of the various molecular events associated with aging and may tie together the various models of aging: the free radical theory, genetically programmed senescence, telomere shortening, genomic instability, nutritional intake, and growth signaling ().
Figure 2. Epigenetic changes resulting in a decrease and redistribution of global heterochromatin may underlie the various models of aging. The models of free radical accumulation, genetically programmed senescence, telomere shortening, genomic instability, nutritional intake and growth signaling are distinct, yet overlapping theories of aging that may all be linked by heterochromatin redistribution.
Acknowledgments
We thank Dr. Louise Silver-Morse and members of the Li laboratory for comments. This work was supported by grants from NIH (R01CA131326), the Leukemia and Lymphoma Society, and the Glenn Award for Research in Biological Mechanisms of Aging to W.X.L.
Related Research Data
References
- Hansen JC. Conformational dynamics of the chromatin fiber in solution: determinants, mechanisms, and functions. Annu Rev Biophys Biomol Struct 2002; 31:361 - 92; http://dx.doi.org/10.1146/annurev.biophys.31.101101.140858; PMID: 11988475
- Luger K, Mäder AW, Richmond RK, Sargent DF, Richmond TJ. Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature 1997; 389:251 - 60; http://dx.doi.org/10.1038/38444; PMID: 9305837
- Radak Z, Chung HY, Goto S. Exercise and hormesis: oxidative stress-related adaptation for successful aging. Biogerontology 2005; 6:71 - 5; http://dx.doi.org/10.1007/s10522-004-7386-7; PMID: 15834665
- Kouzarides T. Chromatin modifications and their function. Cell 2007; 128:693 - 705; http://dx.doi.org/10.1016/j.cell.2007.02.005; PMID: 17320507
- Wallrath LL. Unfolding the mysteries of heterochromatin. Curr Opin Genet Dev 1998; 8:147 - 53; http://dx.doi.org/10.1016/S0959-437X(98)80135-4; PMID: 9610404
- Grewal SI, Moazed D. Heterochromatin and epigenetic control of gene expression. Science 2003; 301:798 - 802; http://dx.doi.org/10.1126/science.1086887; PMID: 12907790
- Martin C, Zhang Y. The diverse functions of histone lysine methylation. Nat Rev Mol Cell Biol 2005; 6:838 - 49; http://dx.doi.org/10.1038/nrm1761; PMID: 16261189
- Eissenberg JC, Elgin SC. The HP1 protein family: getting a grip on chromatin. Curr Opin Genet Dev 2000; 10:204 - 10; http://dx.doi.org/10.1016/S0959-437X(00)00058-7; PMID: 10753776
- Karpen GH, Allshire RC. The case for epigenetic effects on centromere identity and function. Trends Genet 1997; 13:489 - 96; http://dx.doi.org/10.1016/S0168-9525(97)01298-5; PMID: 9433139
- Wutz A. Gene silencing in X-chromosome inactivation: advances in understanding facultative heterochromatin formation. Nat Rev Genet 2011; 12:542 - 53; http://dx.doi.org/10.1038/nrg3035; PMID: 21765457
- Trojer P, Reinberg D. Facultative heterochromatin: is there a distinctive molecular signature?. Mol Cell 2007; 28:1 - 13; http://dx.doi.org/10.1016/j.molcel.2007.09.011; PMID: 17936700
- Villeponteau B. The heterochromatin loss model of aging. Exp Gerontol 1997; 32:383 - 94; http://dx.doi.org/10.1016/S0531-5565(96)00155-6; PMID: 9315443
- Shumaker DK, Dechat T, Kohlmaier A, Adam SA, Bozovsky MR, Erdos MR, et al. Mutant nuclear lamin A leads to progressive alterations of epigenetic control in premature aging. Proc Natl Acad Sci U S A 2006; 103:8703 - 8; http://dx.doi.org/10.1073/pnas.0602569103; PMID: 16738054
- Haithcock E, Dayani Y, Neufeld E, Zahand AJ, Feinstein N, Mattout A, et al. Age-related changes of nuclear architecture in Caenorhabditis elegans. Proc Natl Acad Sci U S A 2005; 102:16690 - 5; http://dx.doi.org/10.1073/pnas.0506955102; PMID: 16269543
- Brandt A, Krohne G, Grosshans J. The farnesylated nuclear proteins KUGELKERN and LAMIN B promote aging-like phenotypes in Drosophila flies. Aging Cell 2008; 7:541 - 51; http://dx.doi.org/10.1111/j.1474-9726.2008.00406.x; PMID: 18494863
- Larson K, Yan SJ, Tsurumi A, Liu J, Zhou J, Gaur K, et al. Heterochromatin formation promotes longevity and represses ribosomal RNA synthesis. PLoS Genet 2012; 8:e1002473; http://dx.doi.org/10.1371/journal.pgen.1002473; PMID: 22291607
- Scaffidi P, Misteli T. Lamin A-dependent nuclear defects in human aging. Science 2006; 312:1059 - 63; http://dx.doi.org/10.1126/science.1127168; PMID: 16645051
- Collado M, Blasco MA, Serrano M. Cellular senescence in cancer and aging. Cell 2007; 130:223 - 33; http://dx.doi.org/10.1016/j.cell.2007.07.003; PMID: 17662938
- Herbig U, Ferreira M, Condel L, Carey D, Sedivy JM. Cellular senescence in aging primates. Science 2006; 311:1257; http://dx.doi.org/10.1126/science.1122446; PMID: 16456035
- Ye X, Zerlanko B, Kennedy A, Banumathy G, Zhang R, Adams PD. Downregulation of Wnt signaling is a trigger for formation of facultative heterochromatin and onset of cell senescence in primary human cells. Mol Cell 2007; 27:183 - 96; http://dx.doi.org/10.1016/j.molcel.2007.05.034; PMID: 17643369
- Adams PD. Remodeling of chromatin structure in senescent cells and its potential impact on tumor suppression and aging. Gene 2007; 397:84 - 93; http://dx.doi.org/10.1016/j.gene.2007.04.020; PMID: 17544228
- Narita M. Cellular senescence and chromatin organisation. Br J Cancer 2007; 96:686 - 91; http://dx.doi.org/10.1038/sj.bjc.6603636; PMID: 17311013
- Jeyapalan AS, Orellana RA, Suryawan A, O’Connor PM, Nguyen HV, Escobar J, et al. Glucose stimulates protein synthesis in skeletal muscle of neonatal pigs through an AMPK- and mTOR-independent process. Am J Physiol Endocrinol Metab 2007; 293:E595 - 603; http://dx.doi.org/10.1152/ajpendo.00121.2007; PMID: 17551002
- Zhang R, Chen W, Adams PD. Molecular dissection of formation of senescence-associated heterochromatin foci. Mol Cell Biol 2007; 27:2343 - 58; http://dx.doi.org/10.1128/MCB.02019-06; PMID: 17242207
- Zhang R, Poustovoitov MV, Ye X, Santos HA, Chen W, Daganzo SM, et al. Formation of MacroH2A-containing senescence-associated heterochromatin foci and senescence driven by ASF1a and HIRA. Dev Cell 2005; 8:19 - 30; http://dx.doi.org/10.1016/j.devcel.2004.10.019; PMID: 15621527
- Sedivy JM, Banumathy G, Adams PD. Aging by epigenetics--a consequence of chromatin damage?. Exp Cell Res 2008; 314:1909 - 17; http://dx.doi.org/10.1016/j.yexcr.2008.02.023; PMID: 18423606
- Narita M, Nũnez S, Heard E, Narita M, Lin AW, Hearn SA, et al. Rb-mediated heterochromatin formation and silencing of E2F target genes during cellular senescence. Cell 2003; 113:703 - 16; http://dx.doi.org/10.1016/S0092-8674(03)00401-X; PMID: 12809602
- Di Micco R, Sulli G, Dobreva M, Liontos M, Botrugno OA, Gargiulo G, et al. Interplay between oncogene-induced DNA damage response and heterochromatin in senescence and cancer. Nat Cell Biol 2011; 13:292 - 302; http://dx.doi.org/10.1038/ncb2170; PMID: 21336312
- Ivanov A, Adams PD. A damage limitation exercise. Nat Cell Biol 2011; 13:193 - 5; http://dx.doi.org/10.1038/ncb0311-193; PMID: 21364567
- Richardson B. Impact of aging on DNA methylation. Ageing Res Rev 2003; 2:245 - 61; http://dx.doi.org/10.1016/S1568-1637(03)00010-2; PMID: 12726774
- Bollati V, Schwartz J, Wright R, Litonjua A, Tarantini L, Suh H, et al. Decline in genomic DNA methylation through aging in a cohort of elderly subjects. Mech Ageing Dev 2009; 130:234 - 9; http://dx.doi.org/10.1016/j.mad.2008.12.003; PMID: 19150625
- Bellizzi D, D’Aquila P, Montesanto A, Corsonello A, Mari V, Mazzei B, et al. Global DNA methylation in old subjects is correlated with frailty. Age (Dordr) 2012; 34:169 - 79; http://dx.doi.org/10.1007/s11357-011-9216-6; PMID: 21336567
- Wilson VL, Jones PA. DNA methylation decreases in aging but not in immortal cells. Science 1983; 220:1055 - 7; http://dx.doi.org/10.1126/science.6844925; PMID: 6844925
- Kim JY, Siegmund KD, Tavaré S, Shibata D. Age-related human small intestine methylation: evidence for stem cell niches. BMC Med 2005; 3:10; http://dx.doi.org/10.1186/1741-7015-3-10; PMID: 15975143
- Kim WY, Sharpless NE. The regulation of INK4/ARF in cancer and aging. Cell 2006; 127:265 - 75; http://dx.doi.org/10.1016/j.cell.2006.10.003; PMID: 17055429
- Issa JP, Ahuja N, Toyota M, Bronner MP, Brentnall TA. Accelerated age-related CpG island methylation in ulcerative colitis. Cancer Res 2001; 61:3573 - 7; PMID: 11325821
- Issa JP, Ottaviano YL, Celano P, Hamilton SR, Davidson NE, Baylin SB. Methylation of the oestrogen receptor CpG island links ageing and neoplasia in human colon. Nat Genet 1994; 7:536 - 40; http://dx.doi.org/10.1038/ng0894-536; PMID: 7951326
- Issa JP, Vertino PM, Boehm CD, Newsham IF, Baylin SB. Switch from monoallelic to biallelic human IGF2 promoter methylation during aging and carcinogenesis. Proc Natl Acad Sci U S A 1996; 93:11757 - 62; http://dx.doi.org/10.1073/pnas.93.21.11757; PMID: 8876210
- Singhal RP, Mays-Hoopes LL, Eichhorn GL. DNA methylation in aging of mice. Mech Ageing Dev 1987; 41:199 - 210; http://dx.doi.org/10.1016/0047-6374(87)90040-6; PMID: 3431172
- So K, Tamura G, Honda T, Homma N, Waki T, Togawa N, et al. Multiple tumor suppressor genes are increasingly methylated with age in non-neoplastic gastric epithelia. Cancer Sci 2006; 97:1155 - 8; http://dx.doi.org/10.1111/j.1349-7006.2006.00302.x; PMID: 16952303
- Waki T, Tamura G, Sato M, Motoyama T. Age-related methylation of tumor suppressor and tumor-related genes: an analysis of autopsy samples. Oncogene 2003; 22:4128 - 33; http://dx.doi.org/10.1038/sj.onc.1206651; PMID: 12821947
- Romanov GA, Vanyushin BF. Methylation of reiterated sequences in mammalian DNAs. Effects of the tissue type, age, malignancy and hormonal induction. Biochim Biophys Acta 1981; 653:204 - 18; PMID: 7225396
- Okano M, Bell DW, Haber DA, Li E. DNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian development. Cell 1999; 99:247 - 57; http://dx.doi.org/10.1016/S0092-8674(00)81656-6; PMID: 10555141
- Lopatina N, Haskell JF, Andrews LG, Poole JC, Saldanha S, Tollefsbol T. Differential maintenance and de novo methylating activity by three DNA methyltransferases in aging and immortalized fibroblasts. J Cell Biochem 2002; 84:324 - 34; http://dx.doi.org/10.1002/jcb.10015; PMID: 11787061
- Casillas MA Jr., Lopatina N, Andrews LG, Tollefsbol TO. Transcriptional control of the DNA methyltransferases is altered in aging and neoplastically-transformed human fibroblasts. Mol Cell Biochem 2003; 252:33 - 43; http://dx.doi.org/10.1023/A:1025548623524; PMID: 14577574
- Jones PL, Veenstra GJ, Wade PA, Vermaak D, Kass SU, Landsberger N, et al. Methylated DNA and MeCP2 recruit histone deacetylase to repress transcription. Nat Genet 1998; 19:187 - 91; http://dx.doi.org/10.1038/561; PMID: 9620779
- Nan X, Ng HH, Johnson CA, Laherty CD, Turner BM, Eisenman RN, et al. Transcriptional repression by the methyl-CpG-binding protein MeCP2 involves a histone deacetylase complex. Nature 1998; 393:386 - 9; http://dx.doi.org/10.1038/30764; PMID: 9620804
- Ng HH, Zhang Y, Hendrich B, Johnson CA, Turner BM, Erdjument-Bromage H, et al. MBD2 is a transcriptional repressor belonging to the MeCP1 histone deacetylase complex. Nat Genet 1999; 23:58 - 61; http://dx.doi.org/10.1038/12659; PMID: 10471499
- Sarraf SA, Stancheva I. Methyl-CpG binding protein MBD1 couples histone H3 methylation at lysine 9 by SETDB1 to DNA replication and chromatin assembly. Mol Cell 2004; 15:595 - 605; http://dx.doi.org/10.1016/j.molcel.2004.06.043; PMID: 15327775
- Wade PA, Gegonne A, Jones PL, Ballestar E, Aubry F, Wolffe AP. Mi-2 complex couples DNA methylation to chromatin remodelling and histone deacetylation. Nat Genet 1999; 23:62 - 6; http://dx.doi.org/10.1038/12664; PMID: 10471500
- Zhang Y, Ng HH, Erdjument-Bromage H, Tempst P, Bird A, Reinberg D. Analysis of the NuRD subunits reveals a histone deacetylase core complex and a connection with DNA methylation. Genes Dev 1999; 13:1924 - 35; http://dx.doi.org/10.1101/gad.13.15.1924; PMID: 10444591
- Fuks F, Burgers WA, Brehm A, Hughes-Davies L, Kouzarides T. DNA methyltransferase Dnmt1 associates with histone deacetylase activity. Nat Genet 2000; 24:88 - 91; http://dx.doi.org/10.1038/71750; PMID: 10615135
- Fuks F, Burgers WA, Godin N, Kasai M, Kouzarides T. Dnmt3a binds deacetylases and is recruited by a sequence-specific repressor to silence transcription. EMBO J 2001; 20:2536 - 44; http://dx.doi.org/10.1093/emboj/20.10.2536; PMID: 11350943
- Fuks F, Hurd PJ, Deplus R, Kouzarides T. The DNA methyltransferases associate with HP1 and the SUV39H1 histone methyltransferase. Nucleic Acids Res 2003; 31:2305 - 12; http://dx.doi.org/10.1093/nar/gkg332; PMID: 12711675
- Geiman TM, Sankpal UT, Robertson AK, Zhao Y, Zhao Y, Robertson KD. DNMT3B interacts with hSNF2H chromatin remodeling enzyme, HDACs 1 and 2, and components of the histone methylation system. Biochem Biophys Res Commun 2004; 318:544 - 55; http://dx.doi.org/10.1016/j.bbrc.2004.04.058; PMID: 15120635
- Rountree MR, Bachman KE, Baylin SB. DNMT1 binds HDAC2 and a new co-repressor, DMAP1, to form a complex at replication foci. Nat Genet 2000; 25:269 - 77; http://dx.doi.org/10.1038/77023; PMID: 10888872
- Robertson KD, Ait-Si-Ali S, Yokochi T, Wade PA, Jones PL, Wolffe AP. DNMT1 forms a complex with Rb, E2F1 and HDAC1 and represses transcription from E2F-responsive promoters. Nat Genet 2000; 25:338 - 42; http://dx.doi.org/10.1038/77124; PMID: 10888886
- Santoro R, Grummt I. Epigenetic mechanism of rRNA gene silencing: temporal order of NoRC-mediated histone modification, chromatin remodeling, and DNA methylation. Mol Cell Biol 2005; 25:2539 - 46; http://dx.doi.org/10.1128/MCB.25.7.2539-2546.2005; PMID: 15767661
- Santoro R, Li J, Grummt I. The nucleolar remodeling complex NoRC mediates heterochromatin formation and silencing of ribosomal gene transcription. Nat Genet 2002; 32:393 - 6; http://dx.doi.org/10.1038/ng1010; PMID: 12368916
- Wang CM, Tsai SN, Yew TW, Kwan YW, Ngai SM. Identification of histone methylation multiplicities patterns in the brain of senescence-accelerated prone mouse 8. Biogerontology 2010; 11:87 - 102; http://dx.doi.org/10.1007/s10522-009-9231-5; PMID: 19434510
- Sarg B, Koutzamani E, Helliger W, Rundquist I, Lindner HH. Postsynthetic trimethylation of histone H4 at lysine 20 in mammalian tissues is associated with aging. J Biol Chem 2002; 277:39195 - 201; http://dx.doi.org/10.1074/jbc.M205166200; PMID: 12154089
- Agherbi H, Gaussmann-Wenger A, Verthuy C, Chasson L, Serrano M, Djabali M. Polycomb mediated epigenetic silencing and replication timing at the INK4a/ARF locus during senescence. PLoS One 2009; 4:e5622; http://dx.doi.org/10.1371/journal.pone.0005622; PMID: 19462008
- Fraga MF, Agrelo R, Esteller M. Cross-talk between aging and cancer: the epigenetic language. Ann N Y Acad Sci 2007; 1100:60 - 74; http://dx.doi.org/10.1196/annals.1395.005; PMID: 17460165
- Fraga MF, Esteller M. Epigenetics and aging: the targets and the marks. Trends Genet 2007; 23:413 - 8; http://dx.doi.org/10.1016/j.tig.2007.05.008; PMID: 17559965
- Dimauro T, David G. Chromatin modifications: the driving force of senescence and aging?. Aging (Albany NY) 2009; 1:182 - 90; PMID: 20157508
- Demontis F, Perrimon N. Integration of Insulin receptor/Foxo signaling and dMyc activity during muscle growth regulates body size in Drosophila. Development 2009; 136:983 - 93; http://dx.doi.org/10.1242/dev.027466; PMID: 19211682
- Scaffidi P, Misteli T. Lamin A-dependent misregulation of adult stem cells associated with accelerated ageing. Nat Cell Biol 2008; 10:452 - 9; http://dx.doi.org/10.1038/ncb1708; PMID: 18311132
- Shi S, Calhoun HC, Xia F, Li J, Le L, Li WX. JAK signaling globally counteracts heterochromatic gene silencing. Nat Genet 2006; 38:1071 - 6; http://dx.doi.org/10.1038/ng1860; PMID: 16892059
- Li WX. Canonical and non-canonical JAK-STAT signaling. Trends Cell Biol 2008; 18:545 - 51; http://dx.doi.org/10.1016/j.tcb.2008.08.008; PMID: 18848449
- Shi M, Lin TH, Appell KC, Berg LJ. Janus-kinase-3-dependent signals induce chromatin remodeling at the Ifng locus during T helper 1 cell differentiation. Immunity 2008; 28:763 - 73; http://dx.doi.org/10.1016/j.immuni.2008.04.016; PMID: 18549798
- Peng JC, Karpen GH. H3K9 methylation and RNA interference regulate nucleolar organization and repeated DNA stability. Nat Cell Biol 2007; 9:25 - 35; http://dx.doi.org/10.1038/ncb1514; PMID: 17159999
- Peng JC, Karpen GH. Heterochromatic genome stability requires regulators of histone H3 K9 methylation. PLoS Genet 2009; 5:e1000435; http://dx.doi.org/10.1371/journal.pgen.1000435; PMID: 19325889
- Blander G, Guarente L. The Sir2 family of protein deacetylases. Annu Rev Biochem 2004; 73:417 - 35; http://dx.doi.org/10.1146/annurev.biochem.73.011303.073651; PMID: 15189148
- Kennedy BK, Gotta M, Sinclair DA, Mills K, McNabb DS, Murthy M, et al. Redistribution of silencing proteins from telomeres to the nucleolus is associated with extension of life span in S. cerevisiae. Cell 1997; 89:381 - 91; http://dx.doi.org/10.1016/S0092-8674(00)80219-6; PMID: 9150138
- Sinclair DA, Guarente L. Extrachromosomal rDNA circles--a cause of aging in yeast. Cell 1997; 91:1033 - 42; http://dx.doi.org/10.1016/S0092-8674(00)80493-6; PMID: 9428525
- Sinclair DA, Mills K, Guarente L. Accelerated aging and nucleolar fragmentation in yeast sgs1 mutants. Science 1997; 277:1313 - 6; http://dx.doi.org/10.1126/science.277.5330.1313; PMID: 9271578
- Kaeberlein M, McVey M, Guarente L. The SIR2/3/4 complex and SIR2 alone promote longevity in Saccharomyces cerevisiae by two different mechanisms. Genes Dev 1999; 13:2570 - 80; http://dx.doi.org/10.1101/gad.13.19.2570; PMID: 10521401
- Oberdoerffer P, Sinclair DA. The role of nuclear architecture in genomic instability and ageing. Nat Rev Mol Cell Biol 2007; 8:692 - 702; http://dx.doi.org/10.1038/nrm2238; PMID: 17700626
- Longo VD, Kennedy BK. Sirtuins in aging and age-related disease. Cell 2006; 126:257 - 68; http://dx.doi.org/10.1016/j.cell.2006.07.002; PMID: 16873059
- Haigis MC, Guarente LP. Mammalian sirtuins--emerging roles in physiology, aging, and calorie restriction. Genes Dev 2006; 20:2913 - 21; http://dx.doi.org/10.1101/gad.1467506; PMID: 17079682
- Sasaki T, Maier B, Bartke A, Scrable H. Progressive loss of SIRT1 with cell cycle withdrawal. Aging Cell 2006; 5:413 - 22; http://dx.doi.org/10.1111/j.1474-9726.2006.00235.x; PMID: 16939484
- Sommer M, Poliak N, Upadhyay S, Ratovitski E, Nelkin BD, Donehower LA, et al. DeltaNp63alpha overexpression induces downregulation of Sirt1 and an accelerated aging phenotype in the mouse. Cell Cycle 2006; 5:2005 - 11; http://dx.doi.org/10.4161/cc.5.17.3194; PMID: 16940753
- Tissenbaum HA, Guarente L. Increased dosage of a sir-2 gene extends lifespan in Caenorhabditis elegans. Nature 2001; 410:227 - 30; http://dx.doi.org/10.1038/35065638; PMID: 11242085
- Wood JG, Rogina B, Lavu S, Howitz K, Helfand SL, Tatar M, et al. Sirtuin activators mimic caloric restriction and delay ageing in metazoans. Nature 2004; 430:686 - 9; http://dx.doi.org/10.1038/nature02789; PMID: 15254550
- Rogina B, Helfand SL. Sir2 mediates longevity in the fly through a pathway related to calorie restriction. Proc Natl Acad Sci U S A 2004; 101:15998 - 6003; http://dx.doi.org/10.1073/pnas.0404184101; PMID: 15520384
- Burnett C, Valentini S, Cabreiro F, Goss M, Somogyvári M, Piper MD, et al. Absence of effects of Sir2 overexpression on lifespan in C. elegans and Drosophila. Nature 2011; 477:482 - 5; http://dx.doi.org/10.1038/nature10296; PMID: 21938067
- Finkel T. Ageing: a toast to long life. Nature 2003; 425:132 - 3; http://dx.doi.org/10.1038/425132a; PMID: 12968159
- Kaeberlein M, Powers RW 3rd, Steffen KK, Westman EA, Hu D, Dang N, et al. Regulation of yeast replicative life span by TOR and Sch9 in response to nutrients. Science 2005; 310:1193 - 6; http://dx.doi.org/10.1126/science.1115535; PMID: 16293764
- Barja G. Free radicals and aging. Trends Neurosci 2004; 27:595 - 600; http://dx.doi.org/10.1016/j.tins.2004.07.005; PMID: 15374670
- Sohal RS, Weindruch R. Oxidative stress, caloric restriction, and aging. Science 1996; 273:59 - 63; http://dx.doi.org/10.1126/science.273.5271.59; PMID: 8658196
- Lin SJ, Kaeberlein M, Andalis AA, Sturtz LA, Defossez PA, Culotta VC, et al. Calorie restriction extends Saccharomyces cerevisiae lifespan by increasing respiration. Nature 2002; 418:344 - 8; http://dx.doi.org/10.1038/nature00829; PMID: 12124627
- Howitz KT, Bitterman KJ, Cohen HY, Lamming DW, Lavu S, Wood JG, et al. Small molecule activators of sirtuins extend Saccharomyces cerevisiae lifespan. Nature 2003; 425:191 - 6; http://dx.doi.org/10.1038/nature01960; PMID: 12939617
- Guarente L, Picard F. Calorie restriction--the SIR2 connection. Cell 2005; 120:473 - 82; http://dx.doi.org/10.1016/j.cell.2005.01.029; PMID: 15734680
- Anderson RM, Bitterman KJ, Wood JG, Medvedik O, Sinclair DA. Nicotinamide and PNC1 govern lifespan extension by calorie restriction in Saccharomyces cerevisiae. Nature 2003; 423:181 - 5; http://dx.doi.org/10.1038/nature01578; PMID: 12736687
- Imai S, Armstrong CM, Kaeberlein M, Guarente L. Transcriptional silencing and longevity protein Sir2 is an NAD-dependent histone deacetylase. Nature 2000; 403:795 - 800; http://dx.doi.org/10.1038/35001622; PMID: 10693811
- Lin SJ, Defossez PA, Guarente L. Requirement of NAD and SIR2 for life-span extension by calorie restriction in Saccharomyces cerevisiae. Science 2000; 289:2126 - 8; http://dx.doi.org/10.1126/science.289.5487.2126; PMID: 11000115
- Lamming DW, Latorre-Esteves M, Medvedik O, Wong SN, Tsang FA, Wang C, et al. HST2 mediates SIR2-independent life-span extension by calorie restriction. Science 2005; 309:1861 - 4; http://dx.doi.org/10.1126/science.1113611; PMID: 16051752
- Riesen M, Morgan A. Calorie restriction reduces rDNA recombination independently of rDNA silencing. Aging Cell 2009; 8:624 - 32; http://dx.doi.org/10.1111/j.1474-9726.2009.00514.x; PMID: 19732046
- Kaeberlein M, Steffen KK, Hu D, Dang N, Kerr EO, Tsuchiya M, et al. Comment on “HST2 mediates SIR2-independent life-span extension by calorie restriction”. [author reply] Science 2006; 312:1312 - , author reply 1312; http://dx.doi.org/10.1126/science.1124608; PMID: 16741098
- Kaeberlein M, Kirkland KT, Fields S, Kennedy BK. Sir2-independent life span extension by calorie restriction in yeast. PLoS Biol 2004; 2:E296; http://dx.doi.org/10.1371/journal.pbio.0020296; PMID: 15328540
- Murayama A, Ohmori K, Fujimura A, Minami H, Yasuzawa-Tanaka K, Kuroda T, et al. Epigenetic control of rDNA loci in response to intracellular energy status. Cell 2008; 133:627 - 39; http://dx.doi.org/10.1016/j.cell.2008.03.030; PMID: 18485871
- Li Y, Tollefsbol TO. p16(INK4a) suppression by glucose restriction contributes to human cellular lifespan extension through SIRT1-mediated epigenetic and genetic mechanisms. PLoS One 2011; 6:e17421; http://dx.doi.org/10.1371/journal.pone.0017421; PMID: 21390332
- Pruitt K, Zinn RL, Ohm JE, McGarvey KM, Kang SH, Watkins DN, et al. Inhibition of SIRT1 reactivates silenced cancer genes without loss of promoter DNA hypermethylation. PLoS Genet 2006; 2:e40; http://dx.doi.org/10.1371/journal.pgen.0020040; PMID: 16596166
- Vaquero A, Reinberg D. Calorie restriction and the exercise of chromatin. Genes Dev 2009; 23:1849 - 69; http://dx.doi.org/10.1101/gad.1807009; PMID: 19608767
- Peng L, Yuan Z, Ling H, Fukasawa K, Robertson K, Olashaw N, et al. SIRT1 deacetylates the DNA methyltransferase 1 (DNMT1) protein and alters its activities. Mol Cell Biol 2011; 31:4720 - 34; http://dx.doi.org/10.1128/MCB.06147-11; PMID: 21947282
- Li Y, Liu L, Tollefsbol TO. Glucose restriction can extend normal cell lifespan and impair precancerous cell growth through epigenetic control of hTERT and p16 expression. FASEB J 2010; 24:1442 - 53; http://dx.doi.org/10.1096/fj.09-149328; PMID: 20019239
- Hass BS, Hart RW, Lu MH, Lyn-Cook BD. Effects of caloric restriction in animals on cellular function, oncogene expression, and DNA methylation in vitro. Mutat Res 1993; 295:281 - 9; PMID: 7507563
- Muñoz-Najar U, Sedivy JM. Epigenetic control of aging. Antioxid Redox Signal 2011; 14:241 - 59; http://dx.doi.org/10.1089/ars.2010.3250; PMID: 20518699
- Chouliaras L, van den Hove DL, Kenis G, Dela Cruz J, Lemmens MA, van Os J, et al. Caloric restriction attenuates age-related changes of DNA methyltransferase 3a in mouse hippocampus. Brain Behav Immun 2011; 25:616 - 23; http://dx.doi.org/10.1016/j.bbi.2010.11.016; PMID: 21172419
- Sinclair DA. Toward a unified theory of caloric restriction and longevity regulation. Mech Ageing Dev 2005; 126:987 - 1002; http://dx.doi.org/10.1016/j.mad.2005.03.019; PMID: 15893363
- Jeong J, Juhn K, Lee H, Kim SH, Min BH, Lee KM, et al. SIRT1 promotes DNA repair activity and deacetylation of Ku70. Exp Mol Med 2007; 39:8 - 13; PMID: 17334224
- Cohen HY, Lavu S, Bitterman KJ, Hekking B, Imahiyerobo TA, Miller C, et al. Acetylation of the C terminus of Ku70 by CBP and PCAF controls Bax-mediated apoptosis. Mol Cell 2004; 13:627 - 38; http://dx.doi.org/10.1016/S1097-2765(04)00094-2; PMID: 15023334
- Chen D, Pan KZ, Palter JE, Kapahi P. Longevity determined by developmental arrest genes in Caenorhabditis elegans. Aging Cell 2007; 6:525 - 33; http://dx.doi.org/10.1111/j.1474-9726.2007.00305.x; PMID: 17521386
- Giannakou ME, Partridge L. Role of insulin-like signalling in Drosophila lifespan. Trends Biochem Sci 2007; 32:180 - 8; http://dx.doi.org/10.1016/j.tibs.2007.02.007; PMID: 17412594
- Blagosklonny MV. Increasing healthy lifespan by suppressing aging in our lifetime: preliminary proposal. Cell Cycle 2010; 9:4788 - 94; http://dx.doi.org/10.4161/cc.9.24.14360; PMID: 21150328
- Tavernarakis N. Ageing and the regulation of protein synthesis: a balancing act?. Trends Cell Biol 2008; 18:228 - 35; http://dx.doi.org/10.1016/j.tcb.2008.02.004; PMID: 18346894
- Kaletsky R, Murphy CT. The role of insulin/IGF-like signaling in C. elegans longevity and aging. Dis Model Mech 2010; 3:415 - 9; http://dx.doi.org/10.1242/dmm.001040; PMID: 20354111
- Wang MC, Bohmann D, Jasper H. JNK signaling confers tolerance to oxidative stress and extends lifespan in Drosophila. Dev Cell 2003; 5:811 - 6; http://dx.doi.org/10.1016/S1534-5807(03)00323-X; PMID: 14602080
- Sun J, Folk D, Bradley TJ, Tower J. Induced overexpression of mitochondrial Mn-superoxide dismutase extends the life span of adult Drosophila melanogaster. Genetics 2002; 161:661 - 72; PMID: 12072463
- Sun J, Molitor J, Tower J. Effects of simultaneous over-expression of Cu/ZnSOD and MnSOD on Drosophila melanogaster life span. Mech Ageing Dev 2004; 125:341 - 9; http://dx.doi.org/10.1016/j.mad.2004.01.009; PMID: 15130751
- Sun J, Tower J. FLP recombinase-mediated induction of Cu/Zn-superoxide dismutase transgene expression can extend the life span of adult Drosophila melanogaster flies. Mol Cell Biol 1999; 19:216 - 28; PMID: 9858546
- Lee SS, Kennedy S, Tolonen AC, Ruvkun G. DAF-16 target genes that control C. elegans life-span and metabolism. Science 2003; 300:644 - 7; http://dx.doi.org/10.1126/science.1083614; PMID: 12690206
- Murphy CT, McCarroll SA, Bargmann CI, Fraser A, Kamath RS, Ahringer J, et al. Genes that act downstream of DAF-16 to influence the lifespan of Caenorhabditis elegans. Nature 2003; 424:277 - 83; http://dx.doi.org/10.1038/nature01789; PMID: 12845331
- Walker GA, Lithgow GJ. Lifespan extension in C. elegans by a molecular chaperone dependent upon insulin-like signals. Aging Cell 2003; 2:131 - 9; http://dx.doi.org/10.1046/j.1474-9728.2003.00045.x; PMID: 12882326
- Harman D. Aging: a theory based on free radical and radiation chemistry. J Gerontol 1956; 11:298 - 300; PMID: 13332224
- Finkel T, Holbrook NJ. Oxidants, oxidative stress and the biology of ageing. Nature 2000; 408:239 - 47; http://dx.doi.org/10.1038/35041687; PMID: 11089981
- Hamilton ML, Van Remmen H, Drake JA, Yang H, Guo ZM, Kewitt K, et al. Does oxidative damage to DNA increase with age?. Proc Natl Acad Sci U S A 2001; 98:10469 - 74; http://dx.doi.org/10.1073/pnas.171202698; PMID: 11517304
- Lu T, Pan Y, Kao SY, Li C, Kohane I, Chan J, et al. Gene regulation and DNA damage in the ageing human brain. Nature 2004; 429:883 - 91; http://dx.doi.org/10.1038/nature02661; PMID: 15190254
- Van Remmen H, Ikeno Y, Hamilton M, Pahlavani M, Wolf N, Thorpe SR, et al. Life-long reduction in MnSOD activity results in increased DNA damage and higher incidence of cancer but does not accelerate aging. Physiol Genomics 2003; 16:29 - 37; http://dx.doi.org/10.1152/physiolgenomics.00122.2003; PMID: 14679299
- Wong KK, Maser RS, Bachoo RM, Menon J, Carrasco DR, Gu Y, et al. Telomere dysfunction and Atm deficiency compromises organ homeostasis and accelerates ageing. Nature 2003; 421:643 - 8; http://dx.doi.org/10.1038/nature01385; PMID: 12540856
- Murga M, Bunting S, Montaña MF, Soria R, Mulero F, Cañamero M, et al. A mouse model of ATR-Seckel shows embryonic replicative stress and accelerated aging. Nat Genet 2009; 41:891 - 8; http://dx.doi.org/10.1038/ng.420; PMID: 19620979
- O’Driscoll M. Life can be stressful without ATR. Nat Genet 2009; 41:866 - 8; http://dx.doi.org/10.1038/ng0809-866; PMID: 19639025
- Ruzankina Y, Pinzon-Guzman C, Asare A, Ong T, Pontano L, Cotsarelis G, et al. Deletion of the developmentally essential gene ATR in adult mice leads to age-related phenotypes and stem cell loss. Cell Stem Cell 2007; 1:113 - 26; http://dx.doi.org/10.1016/j.stem.2007.03.002; PMID: 18371340
- Yan SJ, Lim SJ, Shi S, Dutta P, Li WX. Unphosphorylated STAT and heterochromatin protect genome stability. FASEB J 2011; 25:232 - 41; http://dx.doi.org/10.1096/fj.10-169367; PMID: 20847228
- Harley CB. Telomere loss: mitotic clock or genetic time bomb?. Mutat Res 1991; 256:271 - 82; PMID: 1722017
- Harley CB, Futcher AB, Greider CW. Telomeres shorten during ageing of human fibroblasts. Nature 1990; 345:458 - 60; http://dx.doi.org/10.1038/345458a0; PMID: 2342578
- Ben-Porath I, Weinberg RA. The signals and pathways activating cellular senescence. Int J Biochem Cell Biol 2005; 37:961 - 76; http://dx.doi.org/10.1016/j.biocel.2004.10.013; PMID: 15743671
- Herbig U, Sedivy JM. Regulation of growth arrest in senescence: telomere damage is not the end of the story. Mech Ageing Dev 2006; 127:16 - 24; http://dx.doi.org/10.1016/j.mad.2005.09.002; PMID: 16229875
- Olovnikov AM. Telomeres, telomerase, and aging: origin of the theory. Exp Gerontol 1996; 31:443 - 8; http://dx.doi.org/10.1016/0531-5565(96)00005-8; PMID: 9415101
- Lustig AJ. Mechanisms of silencing in Saccharomyces cerevisiae. Curr Opin Genet Dev 1998; 8:233 - 9; http://dx.doi.org/10.1016/S0959-437X(98)80146-9; PMID: 9610415
- Chien CT, Buck S, Sternglanz R, Shore D. Targeting of SIR1 protein establishes transcriptional silencing at HM loci and telomeres in yeast. Cell 1993; 75:531 - 41; http://dx.doi.org/10.1016/0092-8674(93)90387-6; PMID: 8221892
- Cockell M, Palladino F, Laroche T, Kyrion G, Liu C, Lustig AJ, et al. The carboxy termini of Sir4 and Rap1 affect Sir3 localization: evidence for a multicomponent complex required for yeast telomeric silencing. J Cell Biol 1995; 129:909 - 24; http://dx.doi.org/10.1083/jcb.129.4.909; PMID: 7744964
- Lee HW, Blasco MA, Gottlieb GJ, Horner JW 2nd, Greider CW, DePinho RA. Essential role of mouse telomerase in highly proliferative organs. Nature 1998; 392:569 - 74; http://dx.doi.org/10.1038/33345; PMID: 9560153
- Rudolph KL, Chang S, Lee HW, Blasco M, Gottlieb GJ, Greider C, et al. Longevity, stress response, and cancer in aging telomerase-deficient mice. Cell 1999; 96:701 - 12; http://dx.doi.org/10.1016/S0092-8674(00)80580-2; PMID: 10089885
- Benetti R, García-Cao M, Blasco MA. Telomere length regulates the epigenetic status of mammalian telomeres and subtelomeres. Nat Genet 2007; 39:243 - 50; http://dx.doi.org/10.1038/ng1952; PMID: 17237781
- Benetti R, Gonzalo S, Jaco I, Schotta G, Klatt P, Jenuwein T, et al. Suv4-20h deficiency results in telomere elongation and derepression of telomere recombination. J Cell Biol 2007; 178:925 - 36; http://dx.doi.org/10.1083/jcb.200703081; PMID: 17846168
- Gonzalo S, Jaco I, Fraga MF, Chen T, Li E, Esteller M, et al. DNA methyltransferases control telomere length and telomere recombination in mammalian cells. Nat Cell Biol 2006; 8:416 - 24; http://dx.doi.org/10.1038/ncb1386; PMID: 16565708
- Kosar M, Bartkova J, Hubackova S, Hodny Z, Lukas J, Bartek J. Senescence-associated heterochromatin foci are dispensable for cellular senescence, occur in a cell type- and insult-dependent manner and follow expression of p16(ink4a). Cell Cycle 2011; 10:457 - 68; http://dx.doi.org/10.4161/cc.10.3.14707; PMID: 21248468
- Braig M, Lee S, Loddenkemper C, Rudolph C, Peters AH, Schlegelberger B, et al. Oncogene-induced senescence as an initial barrier in lymphoma development. Nature 2005; 436:660 - 5; http://dx.doi.org/10.1038/nature03841; PMID: 16079837
- Krtolica A, Campisi J. Cancer and aging: a model for the cancer promoting effects of the aging stroma. Int J Biochem Cell Biol 2002; 34:1401 - 14; http://dx.doi.org/10.1016/S1357-2725(02)00053-5; PMID: 12200035
- Lombard DB, Chua KF, Mostoslavsky R, Franco S, Gostissa M, Alt FW. DNA repair, genome stability, and aging. Cell 2005; 120:497 - 512; http://dx.doi.org/10.1016/j.cell.2005.01.028; PMID: 15734682
- Pelicci PG. Do tumor-suppressive mechanisms contribute to organism aging by inducing stem cell senescence?. J Clin Invest 2004; 113:4 - 7; PMID: 14702099