Abstract
[3H]CI-994, a radioactive isotopologue of the benzamide CI-994, a class I histone deacetylase inhibitor (HDACi), was evaluated as an autoradiography probe for ex vivo labeling and localizing of class I HDAC (isoforms 1–3) in the rodent brain. After protocol optimization, up to 80% of total binding was attributed to specific binding. Notably, like other benzamide HDACi, [3H]CI-994 exhibits slow binding kinetics when measured in vitro with isolated enzymes and ex vivo when used for autoradiographic mapping of HDAC1–3 density. The regional distribution and density of HDAC1–3 was determined through a series of saturation and kinetics experiments. The binding properties of [3H]CI-994 to HDAC1–3 were characterized and the data were used to determine the regional Bmax of the target proteins. Kd values, determined from slice autoradiography, were between 9.17 and 15.6 nM. The HDAC1–3 density (Bmax), averaged over whole brain sections, was of 12.9 picomol · mg−1 protein. The highest HDAC1–3 density was found in the cerebellum, followed by hippocampus and cortex. Moderate to low receptor density was found in striatum, hypothalamus and thalamus. These data were correlated with semi-quantitative measures of each HDAC isoform using western blot analysis and it was determined that autoradiographic images most likely represent the sum of HDAC1, HDAC2, and HDAC3 protein density. In competition experiments, [3H]CI-994 binding can be dose-dependently blocked with other HDAC inhibitors, including suberoylanilide hydroxamic acid (SAHA). In summary, we have developed the first known autoradiography tool for imaging class I HDAC enzymes. Although validated in the CNS, [3H]CI-994 will be applicable and beneficial to other target tissues and can be used to evaluate HDAC inhibition in tissues for novel therapies being developed. [3H]CI-994 is now an enabling imaging tool to study the relationship between diseases and epigenetic regulation.
Introduction
Several enzymes control the structure of chromatin and the accessibility of its associated DNA for transcription.Citation1 Of these, two families of enzymes modify the charge state of histone tails through acetylation or deacetylation of lysine residues. Although these enzymes can be promiscuous and are known to react with many non-histone proteins,Citation2 many of their primary biological functions have been ascribed to chromatin remodeling. Histone acetyltransferases (HATs) function by transferring the acetyl group of acetyl coenzyme A to the ε-amino group of certain lysines, which results in neutralization of the normally cationic amino acid. This in turn facilitates chromatin relaxation and can provide DNA access to the transcriptional machinery, as well as lead to the recruitment of acetyl-lysine binding proteins. HAT activity is opposed by histone deacetylases (HDACs), which trigger chromatin condensation and transcriptional repression.Citation3 Dysfunctional regulation of histone acetylation has been implicated in the pathophysiology of several diseases including cancer, heart failure, inflammation and neurodegeneration.Citation4 In some cases, the acetylation status can be ‘rescued’ through chemical inhibition of certain HDACs, a strategy that has been successful in cancer therapy.Citation5-Citation7 The utility of HDACi in treating central nervous system (CNS) disorders is less clear;Citation8 however, abnormal expression levels of class-I HDACs have been correlated with several CNS disorders, including Huntington’s disease,Citation9 amyotrophic lateral sclerosis (ALS),Citation10-Citation12 psychiatric disorders,Citation13 and Alzheimer disease.Citation14
Class I HDACs, consisting of HDAC isoforms 1, 2, 3 and 8, appear to have critical functions in brain development and CNS maintenance.Citation15 For example, HDAC2 silences progenitor transcripts during neuronal differentiation of adult generated neuronsCitation16,Citation17 and negatively regulates memory formation and synaptic plasticity. Class I HDAC inhibition can protect against epigenetic dysfunctions, such as contextual memory defects.Citation18,Citation19 HDAC2 and 3 enzymes have been implicated in neurotoxicity and HDAC inhibitors have potential as neuroprotective drugs.Citation20 HDAC1 and HDAC2 are indispensable for diverse aspects of neuronal development and control the progression of neural precursors to neurons during brain development.Citation21
Although class I HDACs are known to be expressed ubiquitously in the CNS and peripheral organs, information about distribution and density is limited to the relative abundance of its associated mRNA and relative HDAC protein density, as determined by techniques such as western blot and immunoprecipitation.Citation22-Citation26 Absolute protein densities and distributions of HDAC have not been determined, although these data would enlighten our understanding of HDAC function in both healthy and diseased tissue. Moreover, these data would provide a foundation for the development of in vivo imaging agents that could be used to characterize HDAC density, distribution, and ultimately function in humans.Citation27,Citation28 As a stepping stone to these goals, we are developing autoradiography probes that can be used for ex vivo tissue analyses. Development of an autoradiography probe for HDAC has certain advantages and provides complementary information when compared with other ‘visualization techniques’ for HDAC. For example, binding is stoichiometric and quantitative (in an absolute molarity sense); binding is displaceable and can be used to probe small-molecule interactions with HDAC; binding (for the ligand described herein) occurs in the active site and thus the images have direct relevance to the availability of the enzymes for catalysis, which can be important given that perhaps not all complex forms of HDAC can ‘receive’ a ligand or perform catalysis. Herein, we describe the development and validation of an autoradiography ligand, [3H]CI-994, that can be used to characterize class I HDAC enzymes (specifically the sum of HDAC isoforms 1–3) ex vivo.
CI-994, (acetylamino)-N-(2-aminophenyl) benzamide, has demonstrated antitumor activity in vitro and in vivo against a broad spectrum of murine and human tumor models.Citation29-Citation35 CI-994 has also been pursued as a treatment for neurodegenerative diseases and for diseases outside of the brainCitation36,Citation37 For example, it was recently reported that CI-994 inhibition of HDACs may have potential as for the treatment of memory/cognition and anxiety disorders (Tsai, et al. submitted) CI-994 is a class I HDACs inhibitor, selective for HDAC1, 2 and 3 with varying potency.Citation39 Thus, a tritium-labeled version could provide a map of HDAC1–3 and be used to associate this class with disease, to develop class or isoform selective inhibitors, and to monitor enzyme density changes as a function of natural variable (e.g., aging and sex) or disease and treatment. Herein, we describe the development and evaluation of an autoradiography probe, [3H]CI-994, that can be used to measure HDAC1–3 density and distribution in the rodent brain. Furthermore, we demonstrate that [3H]CI-994 can be used in competition experiments to evaluate and explore the affinity and localization of binding for other HDAC inhibitors.
Results and Discussion
Ligand selection and in vitro properties
Autoradiography methods provide a means to determine quantitative maps of protein density provided that a ligand can be developed, which is both selective and specific (saturable). Ideally, a radioligand should have the following desirable properties: high affinity for its target protein(s), high specific radioactivity (which is obviously isotope-dependent), high specific to non-specific binding ratio (determined through saturation), and high selectivity for its target(s). Our studies began by analyzing the structures and physiochemical properties of ligands known to bind HDAC enzymes with high affinity. We were particularly drawn to molecules known to be selective for class I HDACs due to the role of this class in brain disorders (but generally applicable in other tissues as well). We identified CI-994 as a tractable molecule for tritium labeling after first carefully analyzing its binding properties in vitro. This began with an analysis that we believed would be most predictive of non-specific binding, namely brain homogenate binding. CI-994 was compared with other benzamide-based HDAC inhibitors in this assay and found to be unique (). Only 12.9% of CI-994 was bound to mouse plasma protein when allowed to equilibrate with an equal volume of buffer; 14% was bound to mouse homogenate. Moreover, we note that the ligand has similarly good dissociation from human plasma proteins (23.7% bound at equilibrium). These data suggested to us that we would be able to wash away non-specifically bound CI-994 in autoradiography assays by using a series of washing steps.
Figure 1. Structure and properties of HDAC inhibitor CI-994. (A) CI-994 is a benzamide inhibitor with MW 269 Da, CLogP of 0.74, and tPSA of 84. (B) CI-994 inhibits recombinant class I HDAC enzymes at nanomolar concentrations with excellent selectivity over other HDAC isoforms. Of note, CI-994 has low plasma protein and brain tissue binding at equilibrium against buffer. (M = mouse; H = human).
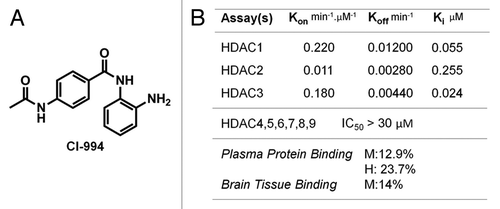
The affinity of CI-994 was determined against the HDAC isoforms 1–3 using a kinetic assay. This provided us with not only the relevant dissociation constant (measured as Ki to be 55, 250, 24 nM, respectively), but also informative information about autoradiography protocol design. These Ki values were similar to those calculated using a trypsin coupled assay with an H4K12Ac peptide substrate (50, 190, 500 nM, respectively, for HDAC1, 2, 3, with the greatest difference for HDAC3 (~20x). Through these binding kinetics experiments we determined that the association of CI-994 was likely slow (inhibition was slow and time-dependent) and, thus, autoradiography incubation would need to be significantly longer than is common. The long dissociation half-life was, accordingly, very long (58–250 min at room temperature) and provided us with an opportunity for long free and non-specifically bound ligand removal. The affinity range provided that we would be able to measure areas of tissue with reasonably low HDAC1–3 density. We also surmised that the development of an HDAC1–3 ligand could provide a means to map each of the three isoforms selectively (if paired appropriately with a selective inhibitor). For example, the use of CI-994 with an HDAC1–2 inhibitor could provide a map of HDAC3 density. Isoform-selective inhibitors within this class are indeed known.Citation40,Citation41
With the promising in vitro data in hand, we invested in the synthesis of [3H]CI-994, which was achieved through non-selective hydrogenation (proton exchange) using tritium-gas. The [3H]CI-994 was prepared in sufficiently high specific activity to proceed with initial autoradiography experiments. These experiments had the primary goal of optimization for specific vs. non-specific binding. Our major findings through these preliminary experiments were that: (1) incubation indeed needed to be long (hours to days), which necessitated tissue morphology stabilization; (2) specific binding was impacted by over-treatment with the fixative solution and morphology stabilization had to be rigorously controlled; (3) there was essentially no difference observed for samples incubated at room temperature vs. 37°C (room temperature was chosen because of increased tissue stability over long incubations); (4) washing needed to be accomplished at 4°C and was best when performed using multiple baths; (5) the identity, concentration and pH of the buffer for incubation and washing impacted the ratio of specific to non-specific binding; 6) DMSO must be used to increase the solubility of non-radioactive CI-994 in blocking experiments. In the end, using the optimized procedure outlined in the methods section, we were able to achieve up to 80% specific binding; however, we must note that there can be significant variation from experiment to experiment and thus non-specific binding must be determined in each tissue-sample pair to extract quantitative data.
Determination of specific binding, protein density, and equilibrium binding constant
Using our optimized protocol, we evaluated target saturation by observing total and non-specific binding as a function of [3H]CI-994 concentration (specific binding was calculated from these data). As seen in , regional distribution of [3H]CI-994 was heterogeneous and thus, we analyzed the saturation data both in terms of the whole brain () and in terms of individual brain regions (). We determined that the whole brain Bmax (assumed to represent HDAC1–3 density) was 12.9 picomol · mg−1 protein and the Kd calculated from saturation was 15.9 nM across the whole brain. In addition to curve fitting analysis, we formatted the data as a Scatchard plot, which was linear, as expected, and provided a Bmax of 13.6 picomol · mg−1 protein and Kd 9.6 nM. These data indicate that the affinity of CI-994 for HDAC in tissue is actually higher than for the isolated recombinant enzymes in vitro and may suggest that HDAC in association with its known binding partners (e.g., in the CoREST complex) may exhibit different binding affinities (and perhaps different selectivity). Moreover, these data support that HDAC class I enzymes are of sufficient concentration in the brain to warrant non-invasive PET imaging (ultimately, in humans). PET imaging of HDAC enzymes is a current area of research; however, to date, the Bmax was not known.
Figure 2. (A) Sagittal and coronal autoradiographic images of total binding and non-specific binding in the rat brain (10 nM [3H]CI-994, 16 h incubation); (B) mean intensity of non-specific binding and total binding of whole rat brain; (C) saturation binding curves for [3H]CI-994 in rat brain: Bmax is12.9 picomol · mg−1 of protein, Kd is calculated as 15.9 nM. and (D) Scatchard plots of [3H]CI-994 binding in rat brain, Bmax is 13.6 picomol · mg−1 of protein, Kd is calculated as 9.59 nM, (n = 6).
![Figure 2. (A) Sagittal and coronal autoradiographic images of total binding and non-specific binding in the rat brain (10 nM [3H]CI-994, 16 h incubation); (B) mean intensity of non-specific binding and total binding of whole rat brain; (C) saturation binding curves for [3H]CI-994 in rat brain: Bmax is12.9 picomol · mg−1 of protein, Kd is calculated as 15.9 nM. and (D) Scatchard plots of [3H]CI-994 binding in rat brain, Bmax is 13.6 picomol · mg−1 of protein, Kd is calculated as 9.59 nM, (n = 6).](/cms/asset/59abeff1-9f1f-430d-b918-84229606e444/kepi_a_10925202_f0002.gif)
Figure 3. (A) Bmax of different regions in the rat brain from [3H]CI-994 autoradiography (n = 6) averaged over each structure. (B) Correlation plot of ΣHDAC1–3 protein density obtained from western (n = 5) vs. HDACs Bmax obtained from [3H]CI-994 autoradiography (n = 6); R2 is 0.9796. (C) Finer detail within brain regions can be observed for example in the hippocampus where [3H]CI-994 binding is higher in CA3 and the dentate gyrus (DG), and in substructures of the cerebellum.
![Figure 3. (A) Bmax of different regions in the rat brain from [3H]CI-994 autoradiography (n = 6) averaged over each structure. (B) Correlation plot of ΣHDAC1–3 protein density obtained from western (n = 5) vs. HDACs Bmax obtained from [3H]CI-994 autoradiography (n = 6); R2 is 0.9796. (C) Finer detail within brain regions can be observed for example in the hippocampus where [3H]CI-994 binding is higher in CA3 and the dentate gyrus (DG), and in substructures of the cerebellum.](/cms/asset/d7fd8a41-f662-4277-b364-9677a8dab189/kepi_a_10925202_f0003.gif)
Validation of imaging data
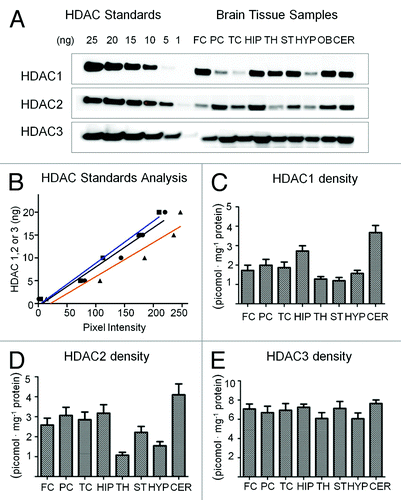
In order to provide proof that the autoradiography maps indeed relate to the HDAC1–3 density, we measured the HDAC1, HDAC2, and HDAC3 density in rodent brain regions using ‘semi-quantitative’ western blots. Brains from five different mice were dissected into nine major regions by the method of Glowinski and Iverson (1966)Citation42 and processed separately as described under experimental procedures. Brain homogenate with known protein content was separated by electrophoresis and antibody binding to tissue sample and reference standards of known concentration was used to determine Bmax ex vivo, . A linear calibration was used to approximate the mass (ng) of HDAC in each brain sample. Using these data, we generated regional distribution plots of each isoform separately (). Qualitatively, our data are consistent with the distribution of mRNA for each isoform provided by the Allen Brain Atlas.Citation43 Of note, HDAC3 protein density is uniform across the brain with a density typically twice that of HDAC1 or HDAC2. These data are in effect ‘average’ values within micro-dissected regions and thus variation that is observed as higher resolution is not detected. Also of note are the high densities of HDAC1 and HDAC2 in the cerebellum and the hippocampus.
Figure 4. HDAC density assessment in mouse brain using western blot analysis (A) western blot analysis of HDAC expression in different regions of mouse brain. Key: Frontal Cortex (FC), Parietal Cortex (PC), Temporal Cortex (TC), Hippocampus (HIP), Thalamus (TH), Striatum (ST), Hypothalamus (HYP), Olfactory Bulb (OB) and Cerebellum (CER). (B) Linearity of HDAC standard analysis by optical densitometry using imageJ analyzer; blue = HDAC1, black = HDAC2, orange = HDAC3 (C) HDAC1 protein density in regions of mouse brain (D) HDAC2 protein density in regions of mouse brain (E) HDAC3 protein density in different regions of mouse brains (n = 5).
Using the western-derived densities and the region-specific binding of [3H]CI-994, we plotted correlation plots of individual isoforms vs. [3H]CI-994, as well as all binary combinations. The best correlation, however, was observed by plotting the sum of isoform density by western () vs. [3H]CI-994-derived Bmax. These were remarkably well correlated and linear with respect to one-another (R2 = 0.98). Bmax values determined by [3H]CI-994 were slightly higher, on average, than those from western-derived values, but this was expected given the extreme differences in the protocols and analyses. From these data, we believe that [3H]CI-994 represents the linear sum of the isoform densities and that the difference in Kd for each isoform (determined in vitro) does not contribute to the selectivity between isoforms (i.e., the lower Ki of HDAC2 in vitro seems to be inconsequential). Correlations alone cannot precisely determine the ratio of HDAC1, HDAC2 and HDAC3 binding by [3H]CI-994. We are currently working to block individual isoforms with selective inhibitors to ascribe the precise ratio of isoform binding. For our preliminary analysis, we evaluated the density of ligand binding averaged across large brain regions. However, we note (as seen in ) that substructure can be observed. For detailed, high resolution assessment of subregions of the hippocampus, long film/phosphor screen exposure times (> 2 mo) will be required; however, with our 7 d exposure data, heterogeneous distribution is noted, for example, in portions of the dentate gyrus.
Determination of ligand binding properties in tissue sections
To rigorously determine the binding parameters of [3H]CI-994 in tissue, we performed kinetic binding experiments. This allowed us to compare Kd determined in our assay (assumed to be at equilibrium) to Kd derived from the on- and off-rates. It also provided a means to analyze whether regional uptake was dominated by rate differences, which could be indicative of different HDAC forms (e.g., isoform or complex density differences). provides a subset of these data: association curves for the hippocampus and thalamus, which respectively had high and low Bmax. The association rates were different in these two regions; however, we also note a difference in the dissociation analysis (). Of particular note is the remarkably slow kinetics of the benzamide ligand family, which dictates long incubation times to reach equilibrium binding (several hours, as opposed to only minutes for most ligand receptor interactions). The regional Kd calculated from these regions are different (9.45 vs. 10.2 nM) and suggest to us that there may be a regional variation in isoform ratio (or perhaps HDAC-complex identity). Additional studies will need to be performed to fully understand the regional variation in [3H]CI-994 affinity.
Figure 5. [3H]CI-994 Binding Kinetics. The observed on and off rates were calculated from association (A) and dissociation (B) curves using the one-phase exponential association and decay equations. Saturation occurred after 24 h. Half saturation was reached around 3 h. Dissociation was slow in both high and low binding regions with a t1/2 around 8 h. Kd can be calculated from the on and off rates: Kd = Koff/Kon. Kd (HIP) is 9.45 nM; and Kd (TH) is 10.2 nM, (n = 6), which is consistent with the Kd obtained from saturation binding value.
![Figure 5. [3H]CI-994 Binding Kinetics. The observed on and off rates were calculated from association (A) and dissociation (B) curves using the one-phase exponential association and decay equations. Saturation occurred after 24 h. Half saturation was reached around 3 h. Dissociation was slow in both high and low binding regions with a t1/2 around 8 h. Kd can be calculated from the on and off rates: Kd = Koff/Kon. Kd (HIP) is 9.45 nM; and Kd (TH) is 10.2 nM, (n = 6), which is consistent with the Kd obtained from saturation binding value.](/cms/asset/30b352de-b962-41f3-af1e-b55fdce65c52/kepi_a_10925202_f0005.gif)
Assessment of competitive binding
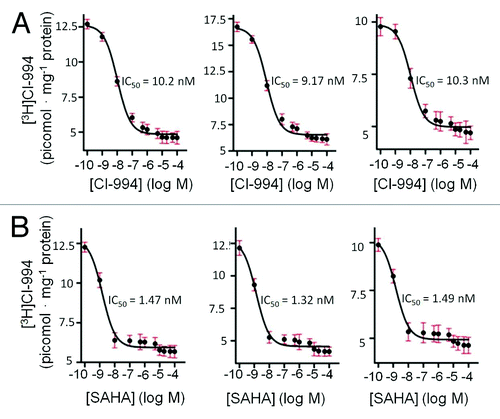
We envisioned that the development of an autoradiography imaging tool for HDAC would be valuable beyond the characterization of protein density and could be applied to the study of novel ligands using competition experiments. As a demonstration of this potential, we validated that the IC50 for CI-994 competition with [3H]CI-994 matched the Kd as anticipated, (). Interestingly, the IC50 showed a parallel trend to the kinetic parameters between various brain regions. For example, the IC50 difference in thalamus and hippocampus was consistent with the kinetically determined Kd in those regions. Regional IC50 differences using novel HDAC inhibitors could be used to improve our knowledge of the relationship between in vitro HDAC isoform affinity and in vivo binding (thus site of action) and efficacy. In order to characterize a well-studied and potent HDAC inhibitor, we determined the IC50 of SAHA using [3H]CI-994 (). The IC50 determined through our assay (1.3–1.5 nM) is consistent with the known potency of SAHA, which is relatively non-selective (as it also targets HDAC6, a class IIb HDAC), but yet highly potent at HDAC1, HDAC2 and HDAC3. We are currently using [3H]CI-994 in competition experiments with HDAC3-selective and HDAC1,2-selective benzamides to explore additional ways to use this autoradiographic tool.
Figure 6. (A) CI-994 self-competition curves (n = 6) in the whole brain slice, hippocampus and thalamus of the rat brain, (left to right); (B) SAHA competition curves in the corresponding rat brain tissues, (n = 6).
Conclusion and Outlook
[3H]CI-994 is now the first validated tool for imaging class I histone deacetylases by autoradiography and can now be used for probing regional variation in HDAC1–3 density in many experiment designs. For example, [3H]CI-994 will be valuable for studying the affinity of novel HDAC inhibitors in tissue sections and it will afford an option for measuring (at least ex vivo) the change in HDAC1–3 expression as a function of normal aging processes and biological diversity (e.g., sex). [3H]CI-994 can be used to characterize HDAC1–3 response to environmental inputs and will be valuable for exploring the relationships that exist between disease state and progression with respect to HDAC density. These studies can be accomplished on preserved human post mortem tissue provided that fixation of the tissue is mild to moderate. In preclinical settings, [3H]CI-994 may be also valuable for exploring HDACi therapy response and mechanism. In preliminary experiments, we have been successful in using [3H]CI-994 to study HDAC1–3 in other tissues (such as the heart) and found that the data provided herein for the brain are generally applicable to other organs.
Ultimately, we envision the development of probes to measure HDAC density in vivo and [3H]CI-994 will be important for prescreening compounds to prioritize for PET imaging. As an in vivo imaging ligand itself, we have used [11C]CI-994 and have determined that the in vivo blood-brain-barrier penetration of CI-994 is not sufficient for PET imaging in non-human primates; however, we are still characterizing its binding properties in other organs. We suspect that faster binding kinetic will be required for in vivo applications and are pursuing other classes of HDAC inhibitors.
Methods
Chemicals and supplies
CI-994 was provided by the Broad Institute, (Cambridge, MA). Synthesis of [3H]CI-994 (specific activity 47.45 Ci/mmol, radioactivity 1.0 mCi/mL, radiochemical purity > 97.7%) was synthesized by Perkin Elmer. Briefly, [3H]CI-994 was synthesized by a non-selective catalytic hydrogen-tritium exchange reaction using tritium gas tuner catalyzed by a hydrogen transfer catalyst (Pd / C) under a carrier-free tritium gas atmosphere at 40 psi. The purity and specific radioactivity were determined by HPLC, TLC and mass spectrometry.
For autoradiography, DMSO was obtained from Sigma. TRIS-HCl and PBS buffer were obtained from Fisher Scientific, Inc.. Paraformaldehyde (4% PFA) in phosphate-buffered saline was purchased from Boston Bioproducts. Scintillation cocktail, scintillation vials and filter paper were from Perkin Elmer. Slide incubation bath with lids, autoradiography cassette was from Fisher Scientific, Inc. For visualization, cyclone plus storage phosphor system, [3H] supersensitive screens were used (Perkin Elmer).
For western blot, reagent kits for protein characterization assay were obtained from Fisher Scientific, Inc.. Primary antibodies: rabbit polyclonal to anti-HDAC1 antibody (ab53091), anti-HDAC2-chip grade antibody (ab7029) and anti-HDAC3 antibody (ab16047), each of which bind to mouse, rat and human HDAC, were purchased from Abcam. The secondary antibody (goat anti-rabbit HRP-conjugate) and most of the western blot experimental supplies e.g. criterion 4–15% pre-cast gel, SDS loading, running and transferring buffers, PVDF membrane were purchased from Bio-Rad Laboratories, Inc.. Biotin serum albumin was from Fisher Scientific, Inc.. Non-fat dry milk was from Labscientific, Inc.. Blue loading buffer was from New England BioLabs, Inc.. Recombinant HDAC1, 2 and3 purified proteins were purchased from BPS Bioscience, Inc.. For imaging, Chemidoc TM XRS+ was used (Bio-Rad Laboratories, Inc.). Micro-dissection tools, forceps, needles, spatulas, razor blade for rats and mice were purchased from Fine Science Tools.
In vitro rate and Kd determination
Slow, tight-binding kinetics of CI-994 with HDACs 1, 2, and 3 were evaluated by the progression curves in inhibition and dilution experiments.Citation44 Kinetic reactions were assembled in microtiter plate wells by adding 1.6 nM, 1.7 nM and 0.5 nM of recombinant, full length, human HDACs 1, 2, and 3 (BPS Biosciences, San Diego), respectively, into separate reaction mixtures containing 2 µM HDAC Caliper carboxyfluorescein (FAM)-labeled AcH4K12 peptide substrate (Broad Substrate A) and CI-994 at different concentrations. Plates were immediately placed into a LabChip EZ Reader II (Caliper/Perkin Elmer) and the wells were sampled continuously throughout a 3-h reaction period. The fluorescent product and substrate were separated based on charge difference between acetylated and deacetylated form, and monitored on the Caliper microfluidic instrument.Citation45 The substrate conversion was calculated with Caliper software. The progression curves at different inhibitor concentrations were analyzed with a nonlinear regression program in Origin 8.5 and fit to the integrated rate equation for slow-binding inhibitors. The off-rate of CI-994 was determined from a dilution experiment. HDAC1 (0.1 µM), HDAC2 (0.1 µM) or HDAC3 (0.05 µM) were incubated with CI-994 at 0.6 µM, 2.5 µM or 0.6 µM respectively for 1 h. The mixtures were then diluted 100-fold into buffer containing 2 µM Broad Substrate A. HDAC-diluted samples were measured continuously over a 4-h period. The recovery of HDAC activity following dilution of the enzyme-[CI-994]-complex is an indication of the reversibility of CI-994. CI-994 off-rates were calculated from dilution progression curve fitting analogously to above.
Western blots
In order to investigate the expression level of class-I HDACs in different brain regions, nine different regions of the mouse brain were micro-dissected. A western blot assay with a known amount of purified human HDACs as standard was used (the affinity of the antibody used should be the same between human and mouse mice HDACs) to calculate the specific amount of HDACs in each brain region. Eight-week old adult WT C57BL/6 mice from Charles River Laboratories International, Inc. (Wilmington, MA) were sacrificed by decapitation in accordance with the animal care and use policies from the Subcommittee on Research Animal Care (SRAC) at Massachusetts General Hospital (MGH). The brains were quickly removed and then washed in ice-cold 1X PBS buffer to remove the blood. Water on the surface of the brain was carefully wicked dry using a Kimwipe tissue. The brain was then transferred to a microscope-slide sitting on a bed of dry ice, loosely covered with aluminum foil and allowed to freeze for 10 min. Before sectioning the brain, it was rinsed with PBS buffer at 4°C. The brain was mounted on the microscope slide and cut in coronal slabs from olfactory bulb to cerebellum with thickness of 1 cm and the slabs were arranged in the Leica dissection microscope in sequence. Slabs were then micro-dissected in coronal orientation at room temperature as quickly as possible. Nine different regions of the mouse brain were dissected which included frontal cortex, parietal cortex, temporal cortex, hippocampus, thalamus, striatum, hypothalamus, olfactory bulb and cerebellum.Citation46 The dissected tissues were placed in Eppendorf tubes and quickly transferred to dry ice. For long-term storage they were later transferred to a -80°C freezer.
Protein preparation
After weighing the collected tissue sections, each tissue was homogenized by treating with 300 µL of lysing buffer (2 × RIPA) (25 mM TRIS-HCl, pH 7.6, 150 mM NaCl, 1% NP-40, 1% sodium deoxycholate, 0.1% SDS). A homogenization pestle was used followed by vortex, incubation and centrifugation to remove insoluble materials. The supernatant was collected and frozen at -20°C; the debris was discarded. The frozen pellets of mouse brain homogenate were thawed on the day of assay. HDAC standard (reference) proteins and the clarified protein supernatant concentration were colorimetrically determined (at 562 nM by BCA protein assay for total protein concentration by following the protocol provided by the Pierce BCA protein assay kit).Citation47 HDAC1, HDAC2 and HDAC3 levels in each of nine brain regions were analyzed by western blot in triplicate. Proteins (15 µg / lane) were separated by SDS-PAGE and then transferred onto PVDF membrane. The membrane was blocked with 5% of non-fat dry milk in 1X Tris-buffered saline containing 0.5% tween-20 for 1 h. The blots were incubated with their respective primary antibody (1:10000) for 16 h at 4°C. Blots were visualized after incubation with an anti-rabbit horseradish peroxidase-conjugated secondary antibody (1:50000) for 1 h, then developed with Bio-Rad Chem. luminescence. The amount of each HDAC in the different regions of mouse brains was determined and normalized to loaded protein mass. Quantitative analysis was performed densitometrically using an Image J analyzer (NIH, Bethesda, MD) and data are reported in pmol · mg−1 protein (not to be confused with pmol · mg−1 tissue).
Plasma and brain homogenate binding assays
Brain binding assays were performed by Sai Advantium Pharma Limited (Hinjewadi). Briefly, rapid equilibrium dialysis was performed with a rapid equilibrium dialysis (RED) device containing dialysis membrane with a molecular weight cut-off of 8,000 Daltons. Each dialysis insert contains two chambers. The red chamber is for plasma or brain homogenate while the white chamber is for buffer. A 200 μL aliquot of CI-994 in brain homogenate (triplicates) was added to the red chamber of dialysis inserts. A 350 μL aliquot of dialysis buffer was added to the buffer chamber of the inserts. Carbamazepine was used as positive control for brain tissue binding. After sealing the RED device with an adhesive film, dialysis was done at 37°C with shaking at 100 rpm for 4 h. A 50 μL aliquot of CI-994 and positive controls was separately added to 0.5 mL Eppendorf tubes. Two aliquots were frozen immediately (0 min sample). Two other aliquots were incubated at 37°C for 4 h along with the RED device. Following dialysis, an aliquot of 50 μL was removed from each well (brain homogenate and buffer) and diluted with equal volume of opposite matrix to nullify the matrix effect. Similarly, buffer was added to recovery and stability samples. An aliquot of 100 μL was submitted for LC-MS/MS analysis. A 25 μL aliquot of the carbamazepine and CI-994 were treated with 100 μL of MeCN containing internal standards (caffeine 200 ng/mL in MeCN) and vortexed for 5 min. The samples were centrifuged at 15000 rpm at 4°C for 10 min and 100 μL of supernatant was submitted for LC-MS/MS analysis. Samples were monitored for parent compound in MRM mode using LC-MS/MS.
Animal procedures and tissue preparation for autoradiography
The experimental procedures were performed according to a protocol approved by the Subcommittee on Research Animal Care (SRAC) at Massachusetts General Hospital (MGH). Brain tissue was obtained by decapitating 8-week adult male Sprague-Dawley rats (150–200 g), in accordance with MGH SRAC’s animal care and use policies. The animals were sacrificed by intraperitoneal injection of pentobarbital. The whole brains were removed and obtained rapidly, after a quick wash in cold 1XPBS; the brains were rapidly frozen by placing the brain on a microscope slide situated on a bed of dry ice. Once frozen, each brain was mounted for cryostat sectioning. Coronal and sagittal sections (20 µm) were consecutively cut and thaw mounted on gelatin-coated slides, which were from Fisher Scientific, Inc.. Slides were stored at -80°C, and based on our experience, should be used no more than two weeks after initial storage.
General experimental method
For most experiments, two parallel experiments were performed. One bath-series was used to measure total binding; and the other was used to determine non-specific binding. The slide-mounted consecutive tissue sections were first fixed with 4% of paraformaldehyde and 2% of ethanol in 1XPBS buffer for 1 h at 4°C to stabilize the morphology of the tissues. The tissue sections were then pre-incubated in 10 mM TRIS-HCl buffer, pH 7.5 for 10 min at room temperature in an attempt to dissociate endogenous ligand and remove any residue from cryostat slide mounting. A second bath-set (2 h at room temperature) was used to pre-saturate the ligand targets (for non-specific binding determination); a vehicle only bath was used for parity in the total binding experiments. Non-specific binding was determined using adjacent sections with 100 µM non-radioactive CI-994 in 5% DMSO and 10 mM TRIS-HCl buffer. (CI-994 should be first dissolved in DMSO then diluted to the concentration as needed by adding the TRIS-HCl buffer.) A third bath-set was used for the [3H]CI-994 incubation. The total binding bath consisted of 10 nM [3H]CI-994 in 5% DMSO and 10 mM TRIS-HCl buffer; the non-specific binding bath consisted of 10 nM [3H]CI-994 and 100 µM non-radioactive CI-994 in 5% DMSO and 10 mM TRIS-HCl buffer. Each incubation bath was covered with a lid to maintain the humidity and incubation was performed for 16 h at room temperature (unless otherwise noted). Following incubation, the slides were transferred into two sequential baths in an attempt to remove free and non-specifically bound [3H]CI-994 (10 mM Tris-Hcl buffer, pH 7.5 at 4°C): the first rinse was performed for 30 sec and the next for 16 h. After washing the sections in buffer, a deionized water bath (4°C) was used to remove buffer salts. The sections were then dried at reduced pressure (in a vacuum oven) at room temperature for 24 h. Once dry, the microscope slide were placed tissue-side down on tritium-sensitive phosphor screens, which were subsequently sealed in autoradiography cassettes and moved to a leaded storage box for 1 week, unless otherwise stated. A Cyclone Phosphor Imager (Perkin Elmer) was used to record and visualize the phosphor-screen data.
Binding optimization experiments
Consecutive tissue sections in coronal and sagittal orientation throughout the rat brain (mounted and fixed as described above) were sequentially numbered divided into two groups: odd numbered tissue sections were used for non-specific binding and even numbered tissue sections were used for total binding. Odd numbered tissue section were treated with 100 µM cold CI-994 plus 10 nM [3H]CI-994 in 5% DMSO and 10 mM TRIS-HCl buffer; even numbered tissue section were treated with 10 nM [3H]CI-994 in 5% DMSO and 10 mM TRIS-HCl buffer. Optimization of incubation time, temperature and the rinsing protocol was achieved systemically and led to standard set of conditions above.
Saturation binding assay
Alternating adjacent sections (as above) were incubated with increasing concentrations of radioligand [3H]CI-994 from 1 nM to 100 nM, either with or without excess 100 µM unlabeled CI-994. Saturation experiments were simultaneously performed on different anatomical regions of the rat brain: the frontal cortex, parietal cortex, temporal cortex, striatum, hippocampus, thalamus, hypothalamus and cerebellum in coronal and sagittal planes, respectively. Sections from each individual brain were exposed to the same phosphor screen film. A calibration standard was used to determine the efficiency difference between each of the phosphor screens used. Bmax and Kd were calculated by curve fitting and through a Scatchard plot analysis.
Dissociation and association kinetics experiments
Thaw mounted tissue sections in both coronal and sagittal orientations were treated with 10 nM [3H]CI-994 either with or without 100 µM unlabeled CI-994 according to the general method for 16 h. After transferring samples to the second washing bath (time zero), samples were removed from the baths at increasing time intervals from 1 to 48 h (according to ) to determine dissociation of both total and non-specifically bound [3H]CI-994. The rate of dissociation for specific bound [3H]CI-994 was calculated from each time-point pair of sample (total – non-specific). To determine association rate, the incubation time with [3H]CI-994 was varied from 1 to 48 h (washing was held constant at 16 h). Specific binding association rate was determined in an analogous manner. Curve-fitting in GraphPad Prism was used to extract koff (dissociation rate), which was used in the determination of kon (association rate). Rate-derived Kd was determined using these rate constants.
Self- and SAHA-competitive (IC50) binding experiments
Two parallel experiments were designed for competitive binding assay: CI-994 (self) and SAHA competition binding experiments. Thaw mounted tissue sections in both coronal and sagittal orientations were treated with unlabeled CI-994 or SAHA in a concentration range from 100 рM to100 µM (according to ) mixed with 10 nM [3H]CI-994 according to the general method. Competition curves were obtained by plotting total binding of [3H]CI-994 against the various (log) concentrations of the competing ligand (i.e., CI-994 or SAHA). IC50 was determined by curve fitting in GraphPad Prism.
Quantitative data analysis (calibration to protein density)
All autoradiography data were analyzed using densitometry using ImageJ Analyzer software (NIH). Specific binding measured in optical density (OD, non-normalized) was obtained by subtracting paired ROIs of non-specific binding and total binding. The non-specific and total binding ODs had been corrected for background signal using a local non-tissue region of the slide area analyzed. All pairs analyzed through subtraction in this way were recorded adjacently on a phosphor screen. Each screen (and data set) was calibrated from OD to nCi mg−1 plastic standard using a [3H]-radioactive standard from American Radiolabeled Chemical, Inc.. The standard samples were analogously treated in terms of ROI size and back subtraction and resulted in a calibration curve fit to a second order polynomial with R2 = 0.99 in Microsoft Excel. The polynomial equation was used for the conversion of OD to nCi mg−1 plastic standard. To determine the relationship between nCi mg−1 plastic standard and nCi · mg−1 protein, eight tissue samples of known size (2.1 cm x 1.0 cm) from cryostat sectioning were cut and weighed. These samples were homogenized as outlined in the western blot analysis details to quantify the protein content in the samples. Using the known tissue section volume, mass, and protein density values that were determined, nCi · mg−1 plastic standard values were converted to nCi mg−1 protein (for our analyses the conversion factor between these two values was 4.14). To convert from nCi to fmol, the specific radioactivity of [3H]CI-994 was used. Thus, the final data were represented in fmol of CI-994 specific binding · mg−1 total protein. Assuming that all specific binding occurs at HDAC1–3, this value represents the total density of HDAC1–3 in the tissue sections.
Acknowledgments
The authors would like to thank Drs. Jinsheng Yang and Wei Zhao for technical support in analysis of the autoradiography data, Dr. Chao Wei’s lab for mouse brain tissue, Drs. Changning Wang and Frederick 'Al' Schroeder for scientific discussion and the NIH for financial support (5R01DA030321, P30DA028800, and R01DA028301).
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
References
- Clayton AL, Hazzalin CA, Mahadevan LC. Enhanced histone acetylation and transcription: a dynamic perspective. Mol Cell 2006; 23:289 - 96; http://dx.doi.org/10.1016/j.molcel.2006.06.017; PMID: 16885019
- Cohen I, Poręba E, Kamieniarz K, Schneider R. Histone modifiers in cancer: friends or foes?. Genes Cancer 2011; 2:631 - 47; http://dx.doi.org/10.1177/1947601911417176; PMID: 21941619
- Spencer CA, Kruhlak MJ, Jenkins HL, Sun X, Bazett-Jones DP. Mitotic transcription repression in vivo in the absence of nucleosomal chromatin condensation. J Cell Biol 2000; 150:13 - 26; http://dx.doi.org/10.1083/jcb.150.1.13; PMID: 10893252
- Haberland M, Montgomery RL, Olson EN. The many roles of histone deacetylases in development and physiology: implications for disease and therapy. Nat Rev Genet 2009; 10:32 - 42; http://dx.doi.org/10.1038/nrg2485; PMID: 19065135
- Hendricks JA, Keliher EJ, Marinelli B, Reiner T, Weissleder R, Mazitschek R. In vivo PET imaging of histone deacetylases by 18F-suberoylanilide hydroxamic acid (18F-SAHA). J Med Chem 2011; 54:5576 - 82; http://dx.doi.org/10.1021/jm200620f; PMID: 21721525
- Wilting RH, Dannenberg JH. Epigenetic mechanisms in tumorigenesis, tumor cell heterogeneity and drug resistance. Drug Resist Updat 2012; 15:21 - 38; http://dx.doi.org/10.1016/j.drup.2012.01.008; PMID: 22356866
- Chavan AV, Somani RR. HDAC inhibitors - new generation of target specific treatment. Mini Rev Med Chem 2010; 10:1263 - 76; http://dx.doi.org/10.2174/13895575110091263; PMID: 20701588
- Shein NA, Shohami E. Histone deacetylase inhibitors as therapeutic agents for acute central nervous system injuries. Mol Med 2011; 17:448 - 56; http://dx.doi.org/10.2119/molmed.2011.00038; PMID: 21274503
- Jia H, Kast RJ, Steffan JS, Thomas EA. Selective histone deacetylase (HDAC) inhibition imparts beneficial effects in Huntington’s disease mice: implications for the ubiquitin-proteasomal and autophagy systems. Hum Mol Genet 2012; 21:5280 - 93; http://dx.doi.org/10.1093/hmg/dds379; PMID: 22965876
- Schmalbach S, Petri S. Histone deacetylation and motor neuron degeneration. CNS Neurol Disord Drug Targets 2010; 9:279 - 84; http://dx.doi.org/10.2174/187152710791292684; PMID: 20406183
- Echaniz-Laguna A, Bousiges O, Loeffler JP, Boutillier AL. Histone deacetylase inhibitors: therapeutic agents and research tools for deciphering motor neuron diseases. Curr Med Chem 2008; 15:1263 - 73; http://dx.doi.org/10.2174/092986708784534974; PMID: 18537606
- Tremolizzo L, Rodriguez-Menendez V, Sala G, Di Francesco JC, Ferrarese C. Valproate and HDAC inhibition: a new epigenetic strategy to mitigate phenotypic severity in ALS?. Amyotroph Lateral Scler Other Motor Neuron Disord 2005; 6:185 - 6; http://dx.doi.org/10.1080/14660820510033614; PMID: 16183561
- Grayson DR, Kundakovic M, Sharma RP. Is there a future for histone deacetylase inhibitors in the pharmacotherapy of psychiatric disorders?. Mol Pharmacol 2010; 77:126 - 35; http://dx.doi.org/10.1124/mol.109.061333; PMID: 19917878
- Kazantsev AG, Thompson LM. Therapeutic application of histone deacetylase inhibitors for central nervous system disorders. Nat Rev Drug Discov 2008; 7:854 - 68; http://dx.doi.org/10.1038/nrd2681; PMID: 18827828
- Fass DM, Schroeder FA, Perlis RH, Haggarty SJ. Epigenetic mechanisms in mood disorders: Targeting neuroplasticity. Neuroscience 2013; •••:S0306 - 4522; PMID: 23376737
- Jawerka M, Colak D, Dimou L, Spiller C, Lagger S, Montgomery RL, et al. The specific role of histone deacetylase 2 in adult neurogenesis. Neuron Glia Biol 2010; 6:93 - 107; http://dx.doi.org/10.1017/S1740925X10000049; PMID: 20388229
- Delcuve GP, Khan DH, Davie JR. Roles of histone deacetylases in epigenetic regulation: emerging paradigms from studies with inhibitors. Clin Epigenetics 2012; 4:5; http://dx.doi.org/10.1186/1868-7083-4-5; PMID: 22414492
- Gräff J, Kim D, Dobbin MM, Tsai LH. Epigenetic regulation of gene expression in physiological and pathological brain processes. Physiol Rev 2011; 91:603 - 49; http://dx.doi.org/10.1152/physrev.00012.2010; PMID: 21527733
- Guan JS, Haggarty SJ, Giacometti E, Dannenberg JH, Joseph N, Gao J, et al. HDAC2 negatively regulates memory formation and synaptic plasticity. Nature 2009; 459:55 - 60; http://dx.doi.org/10.1038/nature07925; PMID: 19424149
- Biermann J, Boyle J, Pielen A, Lagrèze WA. Histone deacetylase inhibitors sodium butyrate and valproic acid delay spontaneous cell death in purified rat retinal ganglion cells. Mol Vis 2011; 17:395 - 403; PMID: 21311741
- Montgomery RL, Hsieh J, Barbosa AC, Richardson JA, Olson EN. Histone deacetylases 1 and 2 control the progression of neural precursors to neurons during brain development. Proc Natl Acad Sci U S A 2009; 106:7876 - 81; http://dx.doi.org/10.1073/pnas.0902750106; PMID: 19380719
- Cardinale JP, Sriramula S, Pariaut R, Guggilam A, Mariappan N, Elks CM, et al. HDAC inhibition attenuates inflammatory, hypertrophic, and hypertensive responses in spontaneously hypertensive rats. Hypertension 2010; 56:437 - 44; http://dx.doi.org/10.1161/HYPERTENSIONAHA.110.154567; PMID: 20679181
- Chen Y, Wang H, Yoon SO, Xu X, Hottiger MO, Svaren J, et al. HDAC-mediated deacetylation of NF-κB is critical for Schwann cell myelination. Nat Neurosci 2011; 14:437 - 41; http://dx.doi.org/10.1038/nn.2780; PMID: 21423191
- Miller RH. Unwrapping HDAC1 and HDAC2 functions in Schwann cell myelination. Nat Neurosci 2011; 14:401 - 3; http://dx.doi.org/10.1038/nn.2788; PMID: 21445062
- MacDonald JL, Roskams AJ. Histone deacetylases 1 and 2 are expressed at distinct stages of neuro-glial development. Dev Dyn 2008; 237:2256 - 67; http://dx.doi.org/10.1002/dvdy.21626; PMID: 18651664
- Broide RS, Redwine JM, Aftahi N, Young W, Bloom FE, Winrow CJ. Distribution of histone deacetylases 1-11 in the rat brain. J Mol Neurosci 2007; 31:47 - 58; http://dx.doi.org/10.1007/BF02686117; PMID: 17416969
- Hooker JM, Kim SW, Alexoff D, Xu Y, Shea C, Reid A, et al. Histone deacetylase inhibitor, MS-275, exhibits poor brain penetration: PK studies of [C]MS-275 using Positron Emission Tomography. ACS Chem Neurosci 2010; 1:65 - 73; http://dx.doi.org/10.1021/cn9000268; PMID: 20657706
- Reid AE, Hooker J, Shumay E, Logan J, Shea C, Kim SW, et al. Evaluation of 6-([(18)F]fluoroacetamido)-1-hexanoicanilide for PET imaging of histone deacetylase in the baboon brain. Nucl Med Biol 2009; 36:247 - 58; http://dx.doi.org/10.1016/j.nucmedbio.2008.12.005; PMID: 19324270
- Undevia SD, Kindler HL, Janisch L, Olson SC, Schilsky RL, Vogelzang NJ, et al. A phase I study of the oral combination of CI-994, a putative histone deacetylase inhibitor, and capecitabine. Ann Oncol 2004; 15:1705 - 11; http://dx.doi.org/10.1093/annonc/mdh438; PMID: 15520075
- Pauer LR, Olivares J, Cunningham C, Williams A, Grove W, Kraker A, et al. Phase I study of oral CI-994 in combination with carboplatin and paclitaxel in the treatment of patients with advanced solid tumors. Cancer Invest 2004; 22:886 - 96; http://dx.doi.org/10.1081/CNV-200039852; PMID: 15641487
- Perabo FG, Müller SC. New agents for treatment of advanced transitional cell carcinoma. Ann Oncol 2007; 18:835 - 43; http://dx.doi.org/10.1093/annonc/mdl331; PMID: 17018703
- Gridelli C, Rossi A, Maione P. The potential role of histone deacetylase inhibitors in the treatment of non-small-cell lung cancer. Crit Rev Oncol Hematol 2008; 68:29 - 36; http://dx.doi.org/10.1016/j.critrevonc.2008.03.002; PMID: 18424067
- Ropero S, Esteller M. The role of histone deacetylases (HDACs) in human cancer. Mol Oncol 2007; 1:19 - 25; http://dx.doi.org/10.1016/j.molonc.2007.01.001; PMID: 19383284
- Hubeek I, Comijn EM, Van der Wilt CL, Merriman RL, Padron JM, Kaspers GJ, et al. CI-994 (N-acetyl-dinaline) in combination with conventional anti-cancer agents is effective against acute myeloid leukemia in vitro and in vivo. Oncol Rep 2008; 19:1517 - 23; PMID: 18497959
- Richards DA, Boehm KA, Waterhouse DM, Wagener DJ, Krishnamurthi SS, Rosemurgy A, et al. Gemcitabine plus CI-994 offers no advantage over gemcitabine alone in the treatment of patients with advanced pancreatic cancer: results of a phase II randomized, double-blind, placebo-controlled, multicenter study. Ann Oncol 2006; 17:1096 - 102; http://dx.doi.org/10.1093/annonc/mdl081; PMID: 16641168
- Chou DH, Holson EB, Wagner FF, Tang AJ, Maglathlin RL, Lewis TA, et al. Inhibition of histone deacetylase 3 protects beta cells from cytokine-induced apoptosis. Chem Biol 2012; 19:669 - 73; http://dx.doi.org/10.1016/j.chembiol.2012.05.010; PMID: 22726680
- Li-Huei Tsai J-SG, Stephen J. Haggarty, Edward Holson, Florence Wagner, Johannes Graeff, Use of CI-994 and dinaline for the treatment of memory/cognition and anxiety disorders. 2010.
- Bradner JE, Mak R, Tanguturi SK, Mazitschek R, Haggarty SJ, Ross K, et al. Chemical genetic strategy identifies histone deacetylase 1 (HDAC1) and HDAC2 as therapeutic targets in sickle cell disease. Proc Natl Acad Sci U S A 2010; 107:12617 - 22; http://dx.doi.org/10.1073/pnas.1006774107; PMID: 20616024
- Cavasin MA, Demos-Davies K, Horn TR, Walker LA, Lemon DD, Birdsey N, et al. Selective class I histone deacetylase inhibition suppresses hypoxia-induced cardiopulmonary remodeling through an antiproliferative mechanism. Circ Res 2012; 110:739 - 48; http://dx.doi.org/10.1161/CIRCRESAHA.111.258426; PMID: 22282194
- Hu E, Dul E, Sung CM, Chen Z, Kirkpatrick R, Zhang GF, et al. Identification of novel isoform-selective inhibitors within class I histone deacetylases. J Pharmacol Exp Ther 2003; 307:720 - 8; http://dx.doi.org/10.1124/jpet.103.055541; PMID: 12975486
- Glowinski J, Axelrod J. Effects of drugs on the disposition of H-3-norepinephrine in the rat brain. Pharmacol Rev 1966; 18:775 - 85; PMID: 5904189
- Allen Brain Atlas. [cited 2012; Available from: http://www.brain-map.org/.
- Zhang Y, Holson EB, Wagner FF. inventors. The Broad Institute, Inc., assignee. Fluorescent substrates for determining lysine modifying enzyme activity. United States patent US WO2013067391. 2013 Oct 05.
- Holson EW, Weïwer M, Zhang YL. inventors. The Broad Institute, Inc., assignee. Inhibitors of histone deacetylases. United States patent US WO2012149540. 2012 Jan 11.
- Li KW, ed. Neuroproteomics (Neuromethods). Dissection of Rodent Brain Regions, ed. S. Spijker. Vol. 57. 2011. 13-26.
- Tyllianakis PE, Kakabakos SE, Evangelatos GP, Ithakissios DS. Direct colorimetric determination of solid-supported functional groups and ligands using bicinchoninic acid. Anal Biochem 1994; 219:335 - 40; http://dx.doi.org/10.1006/abio.1994.1273; PMID: 8080090