Abstract
Chromatin-modifying complexes typically contain signature domains that either have catalytic activity or recognize and bind to specific histone modifications such as acetylation, methylation, and phosphorylation. Despite tremendous progress in this area, much remains to be learned in particular about the mechanistic functions of less well characterized signature domains. One such module is the evolutionary conserved YEATS domain, found in a variety of chromatin-modifying and transcription complexes from yeast to human. Three yeast proteins contain a YEATS domain, including Yaf9, a subunit of both the histone variant H2A.Z deposition complex SWR1-C and the histone acetyltransferase complex NuA4. The three-dimensional structure of the YEATS domain from Yaf9 was solved recently, revealing the existence of three distinct structural regions. One region is characterized by a shallow groove that might constitute a potential acetyl-lysine binding pocket, raising questions about potential protein interaction partners of the Yaf9 YEATS domain.
Introduction
In eukaryotic cells, DNA is packaged together with histone proteins into repeating units of nucleosomes, which constitute the basic building block of chromatin. Chromatin is the principal substrate for many cellular processes including transcription, DNA replication, recombination and repair as the constituent enzymes and associated factors need to access the DNA template in the context of chromatin.Citation42 Histone variants, posttranslational histone modifications and ATP-dependent chromatin remodeling events all contribute to creating distinct structural and functional chromatin “neighborhoods.” The dynamic addition and removal of posttranslational modifications to histones is one important mechanism by which accessibility to chromatin and ultimately DNA is regulated.Citation43 Chemical groups attached to histones include acetylation, methylation, phosphorylation, ubiquitylation, sumoylation and others.Citation1 Rather than occurring and functioning independently, histone modifications often act in a spatially and temporarily coordinated manner and intersect closely with multisubunit complexes that provide “writer,” “eraser” and “reader” functions.Citation2,Citation3 The writer and eraser functions reside in subunits containing catalytic activity that add or remove the mark on histones, while the reader function frequently requires signature domains that specifically recognize and bind to these marks.Citation3 Chromatin-regulating complexes can act in step-wise and/or combinatorial fashion and engage in extensive crosstalk in order to regulate chromatin structure and chromatin accessibility for transcription factors and associated proteins.Citation3,Citation4 Improper target recognition by chromatin readers can have severe consequences, potentially leading to human disease, making these proteins attractive targets for therapeutic intervention.Citation5 The better characterized families of reader pockets include bromodomains, chromodomains, PhD fingers and PWWP domains, to name a few.Citation6 Although these domains clearly provide important chromatin-binding functions through their interaction with histone marks, several intriguing protein domains present in chromatin-modifying complexes have been less explored and largely lack structural and functional information. One of these lesser-known protein domains is the YEATS domain, named after the first proteins recognized to contain this module (Yaf9, ENL, AF9, Taf14, Sas5).Citation7 The primary amino acid sequence of the YEATS domain is evolutionarily conserved from yeast to human and is found in proteins belonging to a variety of chromatin-modifying complexes and transcription factors. In Saccharomyces cerevisiae, three proteins (Yaf9, Taf14 and Sas5) contain a YEATS domain, whereas the most prominent YEATS-proteins in humans, GAS41, ENL and AF9, are all linked to human cancers.Citation7 Here we discuss the recently solved structure of the Yaf9 YEATS domain and its potential functions in light of its similarity to histone chaperone Asf1 and its ability to bind histones.
Yaf9 in Transcription and DNA Repair
Yaf9 is a subunit of both the ATP-dependent chromatin remodeling complex SWR1-C, which deposits histone variant H2A.Z into chromatin and the essential histone acetyltransferase complex NuA4, which acetylates H2A.Z among other substrates.Citation8–Citation13 H2A.Z has roles in a variety of processes, including transcriptional regulation and DNA damage response.Citation14,Citation15 Yaf9 is required for deposition of H2A.Z in vitro as well as in vivo,Citation16,Citation17 suggesting it facilitates the exchange of H2A with H2A.Z in some way. Moreover, the shared module of subunits between NuA4 and SWR1-C, including Yaf9, might participate in recruiting these two complexes to chromatin by binding nucleosomes.Citation18 The current model of extensive crosstalk between NuA4 and SWR1-C converging on H2A.Z suggests that in an initial step, acetylation of histone H4 by NuA4 is required for deposition of H2A.Z by SWR1-C into chromatin and subsequently, NuA4 acetylates H2A.Z itself.Citation9,Citation10,Citation19,Citation20 Through its ability to recognize and bind acetylated H4, the bromo-domain protein Bdf1 has an important role in the recruitment of SWR1-C.Citation20–Citation23 However, other domains in SWR1-C subunits likely also partake in this process, with Yaf9 being an attractive candidate due to its presence in the shared module and its ability to bind histones.Citation16,Citation18 Specifically, it might be the YEATS domain of Yaf9 that mediates this process, as it is required for H2A.Z deposition at specific promoters.Citation16
In addition to functioning in transcription, both NuA4 and SWR1-C have important roles during the DNA damage response.Citation44 The central signaling pathway activated in response to DNA damage caused by double strand breaks involves the phosphorylation of H2A.X at C-terminal serine residues.Citation24 Arp4, which, analagous to Yaf9, is present in the shared subunit module between NuA4 and SWR1-C, physically interacts with phosphorylated H2A.X, thereby recruiting NuA4 to acetylate H4 at double strand breaks.Citation25,Citation26 Subsequently, the SWR1-C and INO80 complexes are recruited to displace phosphorylated H2A.X.Citation27,Citation28 Therefore it is tempting to speculate that Yaf9, as a member of the shared module, may also participate in the recruitment of SWR1-C to DNA damage sites. Alternatively, Yaf9 might facilitate the spatial and temporal coordination of NuA4 and SWR1-C activities at the break site.
Structure and Function of the Yaf9 YEATS Domain
In light of the diverse roles of Yaf9, the recently determined three-dimensional structure of the Yaf9 YEATS domainCitation16 provides an important step forward in understanding the function of Yaf9 and specifically its YEATS domain in these processes. The YEATS domain consists of a beta-sandwich characteristic of the Immunoglobulin (Ig) fold family with 8 antiparallel β-strands capped on one end by 2 short α-helices, and contains three distinct structural features.Citation16 First, a highly conserved cleft is located on the end of the Ig fold opposite the two capping helices. Second, a relatively shallow groove near the N- and C-terminal termini of the YEATS domain is formed in part by the capping helices. Third, a patch rich in conserved charged residues lies between the cleft and the putative peptide-binding groove. Structure-function analysis of the most evolutionarily conserved amino acid residues located in these distinct regions revealed that the YEATS domain of Yaf9 is required for efficient deposition of the histone variant H2A.Z at specific promoters, global H2A.Z acetylation and resistance to genotoxic stressors.Citation16
Interestingly, the overall structure of the YEATS domain of Yaf9 is similar to that of the histone chaperone Asf1, and this structural similarity is congruent with Yaf9's ability to bind histones H3 and H4 in vitro,Citation16 a function that has been well established for Asf1.Citation29–Citation31 Yaf9's connection to Asf1 in yeast extends even further as simultaneous deletion of the two genes encoding them causes dramatic growth defects when compared to the two single deletions, suggesting that these two proteins are involved in a similar process. Yeast strains carrying an unacetylable form of H3K56ac, the histone modification linked to Asf1, have a similar synthetic growth defect in the absence of YAF9,Citation16 indicating that the genetic interaction between YAF9 and ASF1 involves acetylation on H3K56. Taken together, Yaf9's structural and genetic connection to Asf1 and its ability to bind histones H3 and H4 lend support to the hypothesis that the YEATS domain is a chromatin-binding module (i.e., reader) perhaps through interacting with nucleosomes. This might facilitate SWR1-C dependent deposition of H2A.Z during transcription and/or the DNA damage response, in both cases potentially involving coordination with NuA4.
Potential Binding Targets of the Yaf9 YEATS Domain
If the Yaf9 YEATS domain were to function as a reader in the context of recruiting its resident complexes to chromatin, one might speculate that it should have a typical “reader pocket” present in signature domains such as bromodomains or chromodomains.Citation3 In contrast to these typical domains, there currently is no evidence for Yaf9 or other YEATS-domain proteins having the ability to bind to any modified histones. However, based on a more in-depth analysis of the Yaf9 YEATS domain structure, we suggest that Yaf9 has the capability to bind acetylated histones, consistent with a bonafide reader function. One of the distinguishing structural features in the YEATS domain is a shallow groove with a deep hydrophobic pocket that might function as a peptide-binding region.Citation16 Comparative structure analysis of Yaf9 and Asf1 provides an intriguing hint for the function of this pocket and its potential role in YEATS-domain target recognition. Superposition of the two folds, Yaf9 YEATS domain and Asf1, correctly co-aligned the topology of the β-strands based on their switched-Ig fold configuration (). Moreover, Asf1 engages the C-terminal tail of histone H4 in the same orientation and position as the hydrophobic groove of Yaf9 bound to the N-terminal segment of one of its three partner protomers ().Citation29–Citation31 In the crystal, the deep pocket within the groove bound a peptide such that an isoleucine was placed directly over the entrance into the pocket (). Modeling showed that the pocket was deep enough and was of sufficient dimension to accommodate an acetyl-lysine ().
Therefore, Yaf9's putative peptide-binding pocket could function as a reader module that targets specific acetyl-lysines on histones to trigger subsequent chromatin changes. While this is an attractive model, the ability of the Yaf9 YEATS domain to bind acetyl-lysine is clearly speculative and requires detailed analyses with direct peptide-binding assays. However, based on its biological functions and genetic interactions, it is tempting to consider which acetyl-lysine Yaf9 may bind to. One hint comes from work describing a requirement of Asf1/H3K56ac for chromatin assembly and checkpoint recovery after DNA repair, leading to inactivation of the DNA damage checkpoint and cell survival.Citation32 Given the involvement of NuA4 and SWR1-C in the cellular response to DNA double strand breaks, Yaf9, through its YEATS domain, may recognize and bind to H3K56ac during the DNA repair process.
Another possibility is that Yaf9, analogous to Bdf1, binds to acetylated H4 to recruit SWR1-C and NuA4 during transcription to specific promoters to deposit and acetylate H2A.Z, respectively. Compromising Yaf9 YEATS domain function results in loss of H2A.Z at specific promoters,Citation16 consistent with the YEATS domain reading certain histone marks in these regions and thereby helping to recruit the SWR1-C to these regions.
Functional Conservation of the YEATS Domain
It is reasonable to extrapolate our hypothesis of the Yaf9 YEATS domain being chromatin readers in yeast to human YEATS domain-containing proteins as this domain is not only structurally but also functionally conserved through evolution. The closest relative of Yaf9 is the human protein GAS41, which is amplified in glioblastomas and astrocytomas.Citation7,Citation33,Citation34 GAS41 is a subunit of the human TIP60 and SRCAP complexes, which are the equivalents of NuA4 and SWR1-C in yeast.Citation35–Citation38 Interestingly, the GAS41 YEATS domain can substitute for the Yaf9 YEATS domain in yeast,Citation16 providing direct evidence for functional conservation. This makes it likely that the GAS41 YEATS domain also binds histones, although this has not yet been tested to our knowledge. It will be interesting to reconcile this model with GAS41 serving as a co-activator for mammalian transcription factors AP-2 and p53,Citation38,Citation39 specifically as it relates to YEATS-mediated chromatin recognition proposed here. Regardless, histone binding might be a broader and general function of YEATS domains. The human YEATS protein ENL, which is one of the most common fusion partners of the mixed lineage leukemia (MLL) protein in human leukemias, binds to histones H3 and H1 in vitro through its YEATS domain.Citation40
Conclusion
It is well established that chromatin-modifying complexes have writer, eraser and reader functions that reside in multiple signature domains. We hypothesize that YEATS domains, found in a variety of chromatin-modifying and transcription-related complexes, generally may act as a reader of histone marks to recruit its resident complexes to specific chromatin neighborhoods during cellular processes that the particular YEATS protein is involved in. Specifically, for Yaf9's YEATS domain in yeast, we speculate that it might bind acetylated-lysine residues on histones, thereby facilitating the recruitment of NuA4 and SWR1 complexes to specific chromatin neighborhoods during DNA repair or transcription (). Future work will determine the relevant biochemical target of the Yaf9 YEATS domain beyond our speculation, and clarify its role in the recruitment of ATP-dependent chromatin remodeling and histone acetyltransferase activities at specific promoters during transcription or at DNA repair sites. Although the YEATS domain may have one specific target, the combination of all reader, writer and eraser domains in a complex, along with other regulators such as transcription factors, probably is responsible for targeting it to specific chromatin neighborhoods. There is still much to learn about the YEATS domain and its function, but the first structure of it in yeast is a good step forward that may encourage future studies of YEATS domains in other proteins and organisms.
Figures and Tables
Figure 1 Interaction of the Yaf9 YEATS domain hydrophobic groove with its potential target. (A) Structural comparison of the Yaf9 YEATS domain and Asf1 core. Stereo superposition of the Yaf9 YEATS domain on Asf1. The ribbon diagram is colored gray for Yaf9 and green for Asf1. (B) Docking of the Asf1/histone H3–H4 complex onto Yaf9. Note that the N-terminal tail/hydrophobic groove interaction formed between symmetry-related Yaf9 protomers (cyan sticks) spatially and directionally overlaps with the interaction seen between the C-terminal tail of histone H4 (magenta ribbon) and Asf1. In both instances, the peptide from the binding partner docks into an inter-sheet groove and pairs by the last (“h”) strand of the Ig fold. (C) Close up of the Yaf9 YEATS domain hydrophobic groove (surface) and its interaction with the N-terminal segment of adjoining protomer (cyan sticks). An isoleucine in the peptide sits over a deep hole in the floor of the groove. (D) The hole in the hydrophobic groove (surface) of the Yaf9 YEATS domain is sufficiently wide and deep to accommodate a modeled acetyl-lysine residue (white sticks). The modified lysine was modeled using the preferred rotamer library available in PYMOL.Citation41
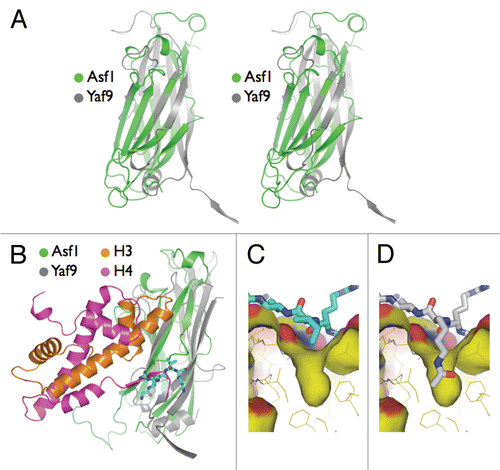
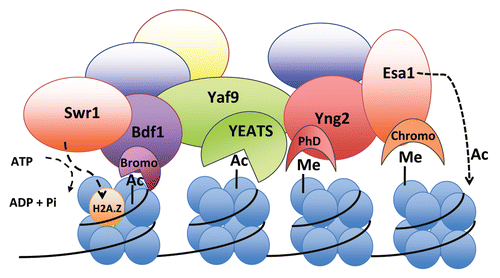
Figure 2 Model highlighting the SWR1-C and NuA4 complexes with their respective “reader” and “writer” subunits containing signature domains that bind to acetyl- or methyl-groups on histones. The SWR1-C subunit Bdf1 has bromodomains that bind to acetylated histones and NuA4 subunits Yng2 and Esa1 have a PhD finger and chromodomain, respectively, that bind to methylated histones.Citation37 Note that the YEATS domain of Yaf9 binding to acetyl groups on histones is speculative in nature and requires further experiments to provide support. Furthermore, it is possible that the subunits bind to the same nucleosome rather than multiple nucleosomes as shown.
Acknowledgements
We thank James M. Berger for discussions and help with the structural modelling and analysis as well as Jasper Rine for helpful discussions about YEATS domain biology. M.S.K.'s laboratory was supported by Canadian Institutes of Health Research (CIHR) Grant MOP-79442. J.M.S. was supported by a fellowship from Child and Family Research and A.Y.W. was supported by a fellowship from CIHR. M.S.K. is a Scholar of MSFHR and of the Canadian Institute for Advanced Research.
References
- Peterson CL, Laniel MA. Histones and histone modifications. Curr Biol 2004; 14:546 - 551
- Ruthenburg AJ, Li H, Patel DJ, Allis CD. Multivalent engagement of chromatin modifications by linked binding modules. Nat Rev Mol Cell Biol 2007; 8:983 - 994
- Taverna SD, Li H, Ruthenburg AJ, Allis CD, Patel DJ. How chromatin-binding modules interpret histone modifications: lessons from professional pocket pickers. Nat Struct Mol Biol 2007; 14:1025 - 1040
- Suganuma T, Workman JL. Crosstalk among histone modifications. Cell 2008; 135:604 - 607
- Baker LA, Allis CD, Wang GG. PHD fingers in human diseases: disorders arising from misinterpreting epigenetic marks. Mutat Res 2008; 647:3 - 12
- de la Cruz X, Lois S, Sanchez-Molina S, Martinez-Balbas MA. Do protein motifs read the histone code?. Bioessays 2005; 27:164 - 175
- Schulze JM, Wang AY, Kobor MS. YEATS domain proteins: a diverse family with many links to chromatin modification and transcription. Biochem Cell Biol 2009; 87:65 - 75
- Lu PY, Levesque N, Kobor MS. NuA4 and SWR1-C: two chromatin-modifying complexes with overlapping functions and components. Biochem Cell Biol 2009; 87:799 - 815
- Babiarz JE, Halley JE, Rine J. Telomeric heterochromatin boundaries require NuA4-dependent acetylation of histone variant H2A.Z in Saccharomyces cerevisiae. Genes Dev 2006; 20:700 - 710
- Keogh MC, Mennella TA, Sawa C, Berthelet S, Krogan NJ, Wolek A, et al. The Saccharomyces cerevisiae histone H2A variant Htz1 is acetylated by NuA4. Genes Dev 2006; 20:660 - 665
- Kobor MS, Venkatasubrahmanyam S, Meneghini MD, Gin JW, Jennings JL, Link AJ, et al. A protein complex containing the conserved Swi2/Snf2-related ATPase Swr1p deposits histone variant H2A.Z into euchromatin. PLoS Biol 2004; 2:131
- Krogan NJ, Keogh MC, Datta N, Sawa C, Ryan OW, Ding H, et al. A Snf2 family ATPase complex required for recruitment of the histone H2A variant Htz1. Mol Cell 2003; 12:1565 - 1576
- Mizuguchi G, Shen X, Landry J, Wu WH, Sen S, Wu C. ATP-driven exchange of histone H2AZ variant catalyzed by SWR1 chromatin remodeling complex. Science 2004; 303:343 - 348
- Draker R, Cheung P. Transcriptional and epigenetic functions of histone variant H2A.Z. Biochem Cell Biol 2009; 87:19 - 25
- Zlatanova J, Thakar A. H2A.Z: view from the top. Structure 2008; 16:166 - 179
- Wang AY, Schulze JM, Skordalakes E, Gin JW, Berger JM, Rine J, et al. Asf1-like structure of the conserved Yaf9 YEATS domain and role in H2A.Z deposition and acetylation. Proc Natl Acad Sci USA 2009; 106:21573 - 21578
- Wu WH, Alami S, Luk E, Wu CH, Sen S, Mizuguchi G, et al. Swc2 is a widely conserved H2AZ-binding module essential for ATP-dependent histone exchange. Nat Struct Mol Biol 2005; 12:1064 - 1071
- Wu WH, Wu CH, Ladurner A, Mizuguchi G, Wei D, Xiao H, et al. N terminus of Swr1 binds to histone H2AZ and provides a platform for subunit assembly in the chromatin remodeling complex. J Biol Chem 2009; 284:6200 - 6207
- Altaf M, Auger A, Monnet-Saksouk J, Brodeur J, Piquet S, Cramet M, et al. NuA4-dependent acetylation of nucleosomal histones H4 and H2A directly stimulates incorporation of H2A.Z by the SWR1 complex. J Biol Chem 2010; 285:15966 - 15977
- Durant M, Pugh BF. NuA4-directed chromatin transactions throughout the Saccharomyces cerevisiae genome. Mol Cell Biol 2007; 27:5327 - 5335
- Matangkasombut O, Buratowski S. Different sensitivities of bromodomain factors 1 and 2 to histone H4 acetylation. Mol Cell 2003; 11:353 - 363
- Boudreault AA, Cronier D, Selleck W, Lacoste N, Utley RT, Allard S, et al. Yeast enhancer of polycomb defines global Esa1-dependent acetylation of chromatin. Genes Dev 2003; 17:1415 - 1428
- Ladurner AG, Inouye C, Jain R, Tjian R. Bromodomains mediate an acetyl-histone encoded antisilencing function at heterochromatin boundaries. Mol Cell 2003; 11:365 - 376
- Shroff R, Arbel-Eden A, Pilch D, Ira G, Bonner WM, Petrini JH, et al. Distribution and dynamics of chromatin modification induced by a defined DNA double-strand break. Curr Biol 2004; 14:1703 - 1711
- Bird AW, Yu DY, Pray-Grant MG, Qiu Q, Harmon KE, Megee PC, et al. Acetylation of histone H4 by Esa1 is required for DNA double-strand break repair. Nature 2002; 419:411 - 415
- Downs JA, Allard S, Jobin-Robitaille O, Javaheri A, Auger A, Bouchard N, et al. Binding of chromatin-modifying activities to phosphorylated histone H2A at DNA damage sites. Mol Cell 2004; 16:979 - 990
- van Attikum H, Fritsch O, Gasser SM. Distinct roles for SWR1 and INO80 chromatin remodeling complexes at chromosomal double-strand breaks. EMBO J 2007; 26:4113 - 4125
- Papamichos-Chronakis M, Krebs JE, Peterson CL. Interplay between Ino80 and Swr1 chromatin remodeling enzymes regulates cell cycle checkpoint adaptation in response to DNA damage. Genes Dev 2006; 20:2437 - 2449
- English CM, Adkins MW, Carson JJ, Churchill ME, et al. Structural basis for the histone chaperone activity of Asf1. Cell 2006; 127:495 - 508
- Natsume R, Eitoku M, Akai Y, Sano N, Horikoshi M, Senda T. Structure and function of the histone chaperone CIA/ASF1 complexed with histones H3 and H4. Nature 2007; 446:338 - 341
- Antczak AJ, Tsubota T, Kaufman PD, Berger JM. Structure of the yeast histone H3-ASF1 interaction: implications for chaperone mechanism, species-specific interactions and epigenetics. BMC Struct Biol 2006; 6:26
- Downs JA. Histone H3 K56 acetylation, chromatin assembly and the DNA damage checkpoint. DNA Repair 2008; 7:2020 - 2024
- Fischer U, Heckel D, Michel A, Janka M, Hulsebos T, Meese E. Cloning of a novel transcription factor-like gene amplified in human glioma including astrocytoma grade I. Hum Mol Genet 1997; 6:1817 - 1822
- Fischer U, Meltzer P, Meese E. Twelve amplified and expressed genes localized in a single domain in glioma. Hum Genet 1996; 98:625 - 628
- Cai Y, Jin J, Florens L, Swanson SK, Kusch T, Li B, et al. The mammalian YL1 protein is a shared subunit of the TRRAP/TIP60 histone acetyltransferase and SRCAP complexes. J Biol Chem 2005; 280:13665 - 13670
- Cai Y, Jin J, Tomomori-Sato C, Sato S, Sorokina I, Parmely TJ, et al. Identification of new subunits of the multiprotein mammalian TRRAP/TIP60-containing histone acetyltransferase complex. J Biol Chem 2003; 278:42733 - 42736
- Doyon Y, Cote J. The highly conserved and multifunctional NuA4 HAT complex. Curr Opin Genet Dev 2004; 14:147 - 154
- Park JH, Roeder RG. GAS41 is required for repression of the p53 tumor suppressor pathway during normal cellular proliferation. Mol Cell Biol 2006; 26:4006 - 4016
- Ding X, Fan C, Zhou J, Zhong Y, Liu R, Ren K, et al. GAS41 interacts with transcription factor AP-2beta and stimulates AP-2beta-mediated transactivation. Nucleic Acids Res 2006; 34:2570 - 2578
- Zeisig DT, Bittner CB, Zeisig BB, Garcia-Cuellar MP, Hess JL, Slany RK. The eleven-nineteen-leukemia protein ENL connects nuclear MLL fusion partners with chromatin. Oncogene 2005; 24:5525 - 5532
- DeLano WL. The PyMOL Molecular Graphics System 2002; San Carlos CA, USA DeLano Scientific
- Ehrenhofer-Murray AE. Chromatin dynamics at DNA replication, transcription and repair. Eur J Biochem 2004; 271:2335 - 2349
- Kouzarides T. Chromatin modifications and their function. Cell 2007; 128:693 - 705
- van Attikum H, Gasser SM. Crosstalk between histone modifications during the DNA damage response. Trends Cell Biol 2009; 19:207 - 217