Abstract
Mounting evidence suggest that epigenetic regulation of brain functions is important in the etiology of psychiatric disorders. These epigenetic regulatory mechanisms, such as DNA methylation and histone acetylation, are influenced by many pharmaceutical compounds including psychiatric drugs. It is therefore of interest to investigate how psychiatric drugs are of influence and what the potential is of new epigenetic drugs for psychiatric disorders. With this targeted review we summarize the current state of knowledge in order to provide insight in this developing field. Several traditional psychiatric drugs have been found to alter the epigenome and in a variety of animal studies, experimental compounds with epigenetic targets have been investigated as potential psychiatric drugs. After discussion of the most relevant epigenetic mechanisms we present the evidence for epigenetic effects for the most relevant classes of drugs.
Introduction
In the mid 1950s, C.H. Waddington revolutionized the field of developmental biology by merging the theories of genetics and development. Previously developmental genetics and its mechanisms were studied separately but Waddington focused on the interplay between genetic factors and embryological growth and differentiation. He coined the term “epigenetics” to describe this novel area.Citation1 Today, “epigenetics” is used to describe “the study of stable alterations in gene expression potential that arise during development and cell proliferation.” Epigenetic processes underlie embryonic cellular development and differentiation, but they are also important in mature mammals, either at random or under influence of the environment (for review seeCitation2). One area in which this is of special importance is the field of psychiatry as emerging evidence suggest that epigenetic mechanisms influence normal patterns of neurodevelopment and cerebral function (for review see ref. Citation3), as well as the aberrant neurodevelopment in psychiatric disorders.Citation4
The epigenetic machinery is extensive and complex and to date not fully understood. Central to our understanding is that epigenetic mechanisms influences transcription of DNA my modifying access to the DNA sequences. depicts the epigenetic mechanisms that are important for this review. Within the nucleus of a cell, the DNA sequence lies wrapped around histone proteins. The complex of DNA, histones and non-histone proteins, such as scaffold and polycomb proteins, forms a highly condensed structure called chromatin. The basic unit of chromatin is the nucleosome. Each nucleosome is connected to the next by a short segment of linker DNA to which a histone H1 protein binds. The string of nucleosomes is folded into a strongly condensed fiber bringing about a strong compaction. Several details on the folding of the nucleosome string are still elusive, and in particular the higher order structure of chromatin is not fully understood. Within the chromatin structure, the DNA sequence is methylated at certain nucleotides. The methylation pattern of DNA across the genome together with the chromatin structure is referred to as the epigenome. As in other cells, epigenetic mechanisms are essential to the development of the nervous system. The epigenetic machinery drives both embryonic and postnatal neural development. It is involved in neurogenesis,Citation5,Citation6 neuronal differentiation, cell fate specificationCitation7 and development of dendrites.Citation8 Interestingly, the course of development of the epigenetic profile is influenced by environmental factors in utero. In different species environmental factors such as temperature or the presence of predators, have been shown to affect the phenotype of the offspring. In humans and mice, the physiology of the baby is affected by the nutritional state of the motherCitation9 and maternal stress in rats also alters the phenotype of their offspring.Citation10 Such epigenetic developmental plasticity may involve preparing the offspring for the type of environment in which they are likely to liveCitation11 but aberrant epigenetic regulation may also lead to disease. Emerging evidence now suggests that vulnerability to psychiatric disorders such as depression and schizophrenia is modulated by epigenetic processes (for review see refs. Citation12 and Citation13).
Figure 1. AUTHOR: Please provide figure legend.
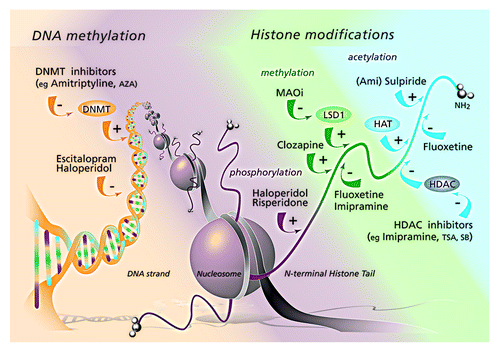
It was previously thought that the epigenetic marks obtained in utero remain the same over time. However, it is now clear that these mechanisms are dynamically regulated. Epigenetic remodeling takes place throughout adult life, under the influence of environmental factors such as nutrition, drugs, and chemical, physical and psychosocial factors (for review see ref. Citation14). The reversible nature of epigenetic marks and their susceptibility to environmental influences makes them potential targets for therapeutic interventions. Indeed, in the past decade many drugs including known psychotropic drugs were found to exert an influence on the epigenome. Therefore, the aim of this review is to summarize the current state of knowledge regarding drug-induced alterations of the epigenome in order to provide insight in this developing field.
Method and scope
Two major types of epigenetic mechanisms that regulate gene expression in the nervous system have been studied to date; (i). Posttranslational histone modification and (ii) DNA methylation. Other types of epigenetic mechanisms such as non-coding RNA’s that can regulate gene expression, and chromatin remodeling mediated by polycomb proteins (for review see ref. Citation15) are likely to also be major mediators of neurodevelopmental plasticity. However, the central mechanisms of these latter processes are largely unknown and will not be discussed in this review. We therefore conducted a systematic literature search from 1996 to July 2011 using Medline from with search terms: (antidepressant OR antipsychotic) AND (epigenetic OR DNA methylation OR histone OR chromatin) ; (HDAC inhibitor OR DNMT inhibitor) AND (depressive disorder OR anxiety OR psychosis OR schizophrenia OR bipolar disorder) ; lithium AND (depressive disorder OR anxiety OR psychosis OR schizophrenia OR bipolar disorder) AND (epigenetic OR methylation OR histone OR chromatin). Cross references were included where relevant.
Posttranslational histone modification
Posttranslational histone modifications take place at the N-terminal histone tail of the nucleosome. depicts the most common modifications. These include small covalent including acetylation, methylation and phosphorylation. Less frequent modifications are ubiquitination, sumoylation, ADP ribosylation and deimination. Their effects on transcriptional activity are not yet well understood and are beyond the scope of this review.
Histone acetylation
Histone acetylation occurs most frequently on the lysine residues at H3 and H4 of the NH2-terminal, though it can also take place at other histones and in the globular domains. This dynamic process is controlled by specific enzymes that either add or remove the acetyl group. Histone acetyl transferases (HATs) catalyze the addition of acetyl groups. Over a dozen HATs have been identified, of which some can also acetylate non-histone proteins such as transcription factors. Several transcription factors even contain intrinsic HAT activity to activate gene expression. Histone deacetylases (HDACs) catalyze the removal of acetyl groups from histone proteins, as well as non-histone proteins among which p53, Sp1 and CREB (for review see ref. Citation16). There are four classes of HDACs, of which class I and II are the most relevant to this review, as the first is expressed throughout brain and body, and the second primarily in the brain, heart and muscle. Histone hyperacetylation is associated with decondensation of chromatin and an increase in gene expression, whereas hypoacetylation correlates with repression of chromatin and a decrease in gene expression. The balance between the opposing activity is important for regulating transcription and disbalance can lead to neurodegenerative diseases (for review see ref. Citation17).
Histone methylation
Histone methylation occurs on lysine residues of the histone tail. This modification can exist in a mono-, di-, or trimethylated state, each with a different effect on transcriptional activity through distinct coregulators. As with acetylation, methylation is regulated by specific enzymes that can exert effects on histone as well as on non-histone proteins. Histone methyltransferases (HMTs) add methyl groups to lysine residues and histone demethylases (HMDs) remove these methyl groups. There are distinct HMTs and HDMs for various lysine residues, each with specific abilities to catalyze mono-, di-, or trimethylated states (for reviews seeCitation18,Citation19). Interestingly, methylation of different lysine residues can achieve opposite effects on gene expression; it can cause both repression and activation depending on which lysine residue of the histone tail is methylated. In psychiatric epigenetics, methylation of histone H3 has gained attention. Trimethylation of histone H3 at lysine 4 (H3K4) is important for transcriptional activation, while the same methylation state is repressive at H3K9 and H3K27, and dimethylation of H3K9 and H3K27 are repressive as well.Citation20
Overall the methylation process gives rise to many unique possibilities to influence transcriptional activity, by combining a variety of enzymes with different lysine residues in different states.
Histone phosphorylation
Although histone phosphorylation is somewhat less well understood, several nuclear protein kinases and protein phosphatases are known that add or remove phosphate groups from the histone tail. The protein kinase MSK 1 and the protein phosphatase inhibitor DARRP‑32 have been shown to regulate phosphorylation in the brain. Phosphorylation of histones is associated with the promotion of transcriptional activity. It is the best characterized at serine 10 on histone H3, where it recruits a HAT to halt repressive methylation on lysine 9. This HAT in turn acetylates the lysine residue in a process called phosphoacetylation, which further increases transcriptional activity (for review see ref. Citation21).
DNA methylation
In cytosine or DNA methylation, a cytosine nucleotide is methylated by transfer of a methyl group from S-adenosyl methionine (SAM), resulting in 5-methyl-cytosine. It is catalyzed by DNA methyltransferases (DNMT) and occurs in approximately 3% of cytosines in human DNA. Recently a new, less frequent, form of DNA methylation has been discovered; 5-methylcytosineCitation22 that has the highest concentration in neuronal cellsCitation23 suggesting a role in neuronal function. ”Traditional” DNA methylation primarily takes place at the dinucleotide sequence CpG. The CpG sequence occurs in low frequency throughout the genome and in high frequency in so-called CpG islands (CGIs). CGIs occur in promoter, intragenic, 3′utr and intergenic regions.Citation24 DNA methylation is associated with transcriptional repression, is enhanced by methyl-binding proteins and also silences genes via blocking binding of certain transcription factors Citation25;Citation26. The cooperation between methyl-binding proteins and chromatin-remodeling enzymes illustrates that different epigenetic mechanisms act in synchrony to influence genetic transcription.
Psychopharmacological compounds with putative epigenetic effects
The recent interest for the role of epigenetics in the brain has led researchers to explore the possibility that drugs can modify epigenetic processes involved in psychiatric disorders. So far, several psychiatric drugs have been found to influence epigenetic mechanisms, suggesting that this may partially explain their mechanism of action. Moreover, the therapeutic potential of experimental compounds that specifically target epigenetic mechanisms is extensively investigated. We aim to give a broad overview of the current knowledge of drugs that modify epigenetic changes in psychiatric disorders.
Antidepressants
Imipramine
Imipramine is a tricyclic antidepressant that has been used since the 1950s. Its primary mechanism of action is to inhibit the reuptake of serotonin and norepinephrine, thus elevating the levels of these neurotransmitters in the brain. Recently, epigenetic effects of imipramine have been uncovered, providing a deeper understanding into the therapeutic effect of this drug. Since dysfunction of brain-derived neurotrophic factor BDNF is often implicated in the etiology of depression, Tsankova et al.Citation27 investigated the effectiveness of imipramine on epigenetic regulation of the Bdnf gene in the hippocampi of mice. Chronic social defeat led to repression of Bdnf and an increase in histone dimethylation of H3K27. Chronic treatment with imipramine could not reverse this, even though the depression-like behaviors disappeared. However chronic imipramine did lead to hyperacetylation of H3 at the Bdnf promoters, mediated by downregulation of Hdac5. Moreover, the efficacy of imipramine was blocked by overexpression of Hdac5, suggesting that downregulation of Hdac5 is essential to the efficacy of imipramine. The authors suggest that since histone H3 hypermethylation was not affected by imipramine, this remains a possible target for antidepressant therapy. Imipramine did however largely reverse repression of dimethylation of H3K9 and H3K27 and increased phospho-CREB binding in the nucleus accumbens induced by social defeat stress.Citation28 The global pattern of H3 dimethylation in the nucleus accumbens of mice which did not develop depression-like behaviors strongly resembled the dimethylation profile of mice that received chronic imipramine treatment after social defeat. This indicates that resilient animals somehow naturally overcome the effects of stress on the accumbens. The genes that show dimethylation in resilient mice, but not in mice treated with imipramine, may therefore provide novel targets for antidepressant medication.Citation28
Amitriptyline
Amitriptyline is a tricyclic antidepressant. In rat astrocytes, amitriptyline induced partially reversible global DNA hypomethylation without affecting histone acetylation and reduced enzymatic activity of DNA (cytosine-5)-methyltransferase 1 (DNMT1) without altering DNMT1 protein levels.Citation29 In vitro amitriptyline did increase H3 acetylation by inhibiting HDAC activity.Citation30
Fluoxetine
Fluoxetine is a widely used antidepressant, belonging to the class of selective serotonin reuptake inhibitors (SSRI). In the hippocampus, fluoxetine reversed decreased histone H3K9 trimethylation but not H3K4 trimethylation induced by chronic restraint stress.Citation31 However postnatal fluoxetine exposure did not changes the acetylation or trimethylation of histone H3 at the BDNF promoters.Citation32 Chronic fluoxetine treatment of healthy rats decreases acetylation of H3 in three serotonin projection areas; the caudate putamen, the frontal cortex and the dentate gyrus of the hippocampus. In addition, expression of the methyl-binding proteins MeCP2 and MBD1 was increased, accompanied by increased Hdac2 expression, further inhibiting transcriptional activity in these brain regions.Citation33 In mice that have been subjected to social defeat stress, reduced H3 acetylation in the hippocampus was reversed by fluoxetine in the same way as histone deacetylases (HDAC) inhibitors, suggesting a similar mechanism of action.Citation34
Escitalopram
The SSRI escitalopram was able to reverse hypermethylation at the P11 (S100A10) locus in a genetic rodent model of depression. Expression of P11 is associated with depression suggesting a putative epigenetic mechanism of escitalopram efficacy.Citation35
MAO inhibitors
Monoamine oxidase (MAO) inhibitors form a group of highly effective antidepressants. They act by inhibiting the MAO enzymes, thereby preventing the breakdown of the monoaminergic neurotransmitters serotonin, dopamine, epinephrine and norepinephrine. The MAO inhibitors phenelzine and tranylcypramine both inhibit demethylation of histone H3K4, resulting in a global increase in H3K4 methylation by breaking down lysine-specific demethylase 1 (LSD1), a histone demethylase that is structurally similar to MAO A and B.Citation36,Citation37 LSD1 specifically demethylates mono- and dimethylated H3K4 and H3K9, thus inhibiting transcription.Citation37 Interestingly, tranylcypromine was ten times more effective in inhibiting LSD1 than MAO A or B, suggesting that also other selective LSD1 inhibitors might be effective in treating depression.Citation38 Other experimental MAO inhibitors also increase H3K4 methylation,Citation36 but the effects of such drugs on H3K9 methylation have not been investigated to date.
Antipsychotics
Haloperidol
Haloperidol is a widely used typical antipsychotic that preferentially antagonizes dopamine D2 receptor activity. In addition, some evidence indicates that haloperidol induces histone modifications to achieve its therapeutic effects. In the striatum of mice infusion of haloperidol led to phosphorylation of histone H3 at serine 10, acetylation of H3K14 as well as phospho-acetylation. These processes were mediated by the NMDA receptor, since administration of an NMDA receptor antagonist blocked phosphoacetylation.Citation39 Similar results on phosphorylation were seen following administration of the D2 receptor antagonist raclopride.Citation40 Two studies also reported an increase in mouse striatal H3 phosphorylation in response to haloperidol, although acetylation levels remained unchanged.Citation40,Citation41 One study reported that haloperidol decreased global DNA methylation levels in the brain of female rats.Citation42 However, haloperidol did not influence methylation of the reelin and GAD67 gene promoters in the frontal cortex or striatum.Citation43
Clozapine
The atypical antipsychotic clozapine binds dopamine D1, D2, and 5HT2 receptors. In the rat cerebral cortex it increased trimethylation of H3K4 by 3-fold and accompanied by increased expression of the H3K4-specific histone methyltransferase gene Mll1. Mll1 occupancy at the GAD67 promoter had doubled.Citation44 In the prefrontal cortex of humans who had been treated with clozapine, H3K4 trimethylation at the GAD67 gene had increased by 2-fold.Citation44 GAD is an enzyme that catalyzes the decarboxylation of glutamate to GABA and CO2. As the epigenetic modification at the GAD64 locus is associated with transcriptional activation, these findings suggest that the therapeutic action of clozapine relies at least in part on increased GABAergic activity mediated by histone methylation.Citation45 Further support for this hypothesis comes from the finding that methionine-induced reelin promoter hypermethylation was strongly decreased after administration of clozapine, together with an increase in promoter-associated H3K9 and H3K14 acetylation.Citation43 Clozapine reduced cytosine methylation at the Gadd45-β locus similar to the effects of brain-permeant mGlu2/3 receptor agonist LY379268 on hypermethylation.Citation46
Sulpiride and Amisulpiride
Two bezamine derivates with antipsychotic properties, sulpiride and amisulpiride, act by antagonizing the dopamine D2 receptor. A third benzamine derivate, MS-275 acts as HDAC inhibitor and is highly effective in increasing acetylation of H3 associated with Reelin and Gad67 gene promoters in the frontal cortices of mice. These drugs have the same effect in the hippocampus and the striatum, but much higher doses are needed, indicating there is different sensitivity to these drugs across different regions of the brain.Citation47 Indeed H3K9 and H3K14 acetylation was increased in the promoter of the Reelin gene after administration of sulpiride in the frontal cortex and striatum of mice that displayed methionine-induced hypermethylation of the reelin promoter, In addition sulpiride decrease methylation at this promotor.Citation43
Risperidone
Risperidone is another atypical antipsychotic that antagonizes D2 receptors, and binds to 5HT2 receptors. It induces global phospho-acetylation of H3 in the striatum, mediated by the NMDA receptor, suggesting that dopamine, serotonin and glutamate act in synchrony to influence chromatin regulation.Citation48
Mood stabilizers
Valproate
Valproic acid (VPA) is a short fatty-chain acid commonly used in the treatment of epilepsy and bipolar disorder. It increases GABAergic activity by inhibiting GABA transaminase, an enzyme that is involved in the synthesis and degradation of GABA. Moreover, VPA is the most extensively investigated compound in psychiatric epigenetics and a potent HDAC inhibitor of class I and II. It increases levels of acetylated histone H3 and H4 thus promoting gene expression. Much effort has been dedicated to elucidating the effects of VPA on neuronal processes in the brain. The therapeutic actions of VPA may be mediated through increase of tyrosine hydroxylase (TH) of which VPA increases mRNA and protein levels.Citation49 In term TH is involved in the biosynthesis of the catecholamines dopamine, norepinephrine and epinephrine, neurotransmitters long implicated in the mechanisms underlying psychiatric disorders. When VPA is administered to rat cortical neurons, mRNA levels and protein levels of exon 1–9-containing Bdnf are elevated, and in astrocytes, glial-derived neurotrophic factor is increasedCitation50-Citation52 accompanied by increased H3 and H4 acetylation.Citation50-Citation52 In a study on rat glioma cells, VPA failed to induce any change in levels of Bdnf mRNA per se, but it enhanced the stimulatory effect of serotonin on Bdnf gene expression.Citation53 Changes in histone H3 and H4 acetylation were found after four weeks of VPA treatment in bipolar and schizophrenia patientsCitation54 and increased glutamic acid decarboxylases (GAD) (involved in GABA synthesis) in response to HDAC inhibitors was found neuronal progenitor cells and lymphoblast of healthy humans.Citation55,Citation56 Increased histone acetylation was associated with VPA-induced hippocampal neurogenesis in hippocampal extracts.Citation57 Further, in the frontal cortex of schizophrenic and bipolar disorder patients with psychosis, DNMT1 mRNA and protein expression was preferentially increased in layer I, II, and IV interneurons when patients received antipsychotic monotherapy, but not when they were treated with a combination of VPA and antipsychoticsCitation58 and only in schizophrenia not bipolar disorder.Citation59 The potential role of epigenetic modification of GABA regulation is recently reviewed by Guidotti.Citation45
Another gene that is targeted by VPA is RELN which has been studied as candidate gene for bipolar disorder and schizophrenia. In neural progenitor cells VPA increased acetylation of H3 and H4 and induced expression of reelin.Citation60,Citation61 This is most likely mediated by demethylation of the reelin promoter region, as treatment with VPA has been shown to stimulate promoter demethylation at the reelin and Gad67 genes.Citation61,Citation62 Additionally, Dnmt1 and 3A and B protein levels were downregulated in response to VPA further increasing gene expression.Citation55,Citation62 When VPA was administered together with methionine, it blocked the hypermethylation that is normally induced by methionine alone.Citation63,Citation64 In addition, VPA attenuated behavioral effects that were induced by methionine. It was suggested that this was mediated by VPA induced hyperacetylation of histone H3 at the reelin and GAD67 promoters, which facilitated a state of euchromatin and enhanced DNA demethylase activity. In mice that exhibit methionine-induced hypermethylation, VPA induced demethylation of the reelin promoter in the frontal cortex, and increased acetylation of H3K9 and H3K14 associated with the promoter. When VPA was administered in conjunction with clozapine or sulpiride, both demethylation and acetylation were further enhanced.Citation43
In an extensive study, Perisic and coworkers examined epigenetic effects of a variety of psychoactive compounds including VPA in hippocampal and cortical astrocytes. VPA was the only drug to exert effects on histone acetylation, inducing dose-dependent global hyperacetylation of histones H3 and H4, and decreased levels of inhibitory H3K9 dimethylation. Furthermore, the increased acetylation by VPA caused a 4-fold increase in glutamate transporter GLT-1 mRNA and reduced DNA methylation of the GLT‑1 gene, which supports the hypotheses of glutamate hypofunction in bipolar disorder. In addition, VPA induced global demethylation of CpG sites in astrocytes of the cortex and hippocampus. The effects on acetylation were transient, since they peaked during the first hour of treatment and then declined. Fourty-eight hours after VPA treatment acetylation levels had returned to baseline, suggesting that the effects of VPA are fully reversible.Citation29 In glioma cells VPA increased expression of HDAC 1, 2 and 3, and of MeCP2, suggesting VPA affects both histone acetylation and DNA methylation although the upregulation of HDACs and MeCP2 may constitute a compensatory mechanism to VPA-induced hyperacetylaton.Citation65 In these cells VPA increased melatonin MT1 receptor expression providing a possible mechanism of action of VPA in bipolar disorder. A final clue to an epigenetic mechanism of action is the finding that in rats, VPA heightened levels of histone H4 acetylation in the striatum, associated with the fosB promoter gene. Striatal fosB protein levels were also elevated, and since fosB is a ubiquitous transcription factor this indicates that VPA can enhance gene expression in the striatum.Citation66
HDAC inhibitors
A well-balanced regulation of HDACs and HATs is essential to gene transcription. Compounds that inhibit HDACs have been extensively studied in models of neurodegenerative disorders. HDAC inhibitors differ in their specificity for the separate classes of HDACs, although most target class I and II. HDAC inhibitors can be divided into categories based on their chemical structure and the most important are the hydroxamates, the short fatty-chain acids and the benzamides. Many more HDAC inhibitors exist that have been primarily investigated in the treatment of cancer, such as SAHA, trapoxin and LBH589 (for review see refs. Citation16 and Citation67). Unfortunately, HDAC properties of psychiatric drugs are largely unknown with the exception of valproate, imipramine and amitriptyline, but other HDAC inhibitors do show some potential.
Sodium butyrate
Sodium butyrate (SB) is a short fatty-chain acid that is used in many experimental paradigms to inhibit histone deacetylase. It selectively inhibits HDAC class I and II, and it is applied in a range of studies to elucidate the involvement of HDACs in development and disease (for review see ref. Citation68). Similar to VPA, SB upregulates BDNF and GDNF mRNA levels in astrocytes, with marked increases in GDNF promoter activity and promoter-associated histone H3 acetylation.Citation51 As BDNF is involved in memory formation, SB might have beneficial effects on this process, as indeed was shown.Citation69 Treatment with SB also enhanced long-term memory for object recognition in mice, and improved memory formation in a fear conditioning task. Following fear conditioning, mice had elevated trimethylation levels of H3K4 and dimethylation of H3K9. Treatment with SB further elevated trimethylation while it diminished dimethylation, enhancing gene expression. This shows that while SB primarily is an HDAC inhibitor, it also affects histone methylation.Citation70 In a contextual fear model, fear extinction was highly accelerated by SB therapy, with fear decreasing eight times as fast as in the control condition, providing further support for learning and memory enhancement of SB.Citation71 More inconsistent results have been reported on the effectiveness of SB on depression-like behavior in animals. Despite increased histone H3 and H4 acetylation in the hippocampus and the frontal cortex following SB administration SB only improved depression-like behavior in three of seven of depression-like behavior.Citation72,Citation73 Further, SB induced a dose-dependent global increase in acetylation of histone H3 and H4.Citation29 In the central nucleus SB induces acetylation and phosphoacetylation of histone H3K14, accompanied by increased c-Fos expression.Citation74 Similar to VPA, SB increased TH mRNA and protein levelsCitation75 suggesting a role in catecholaminergic pathways and bearing promise for a potential therapeutic role in psychiatric disorders.
Trichostatin A
Originally an antifungal antibiotic, two decades ago Trichostatin A (TSA) was discovered to possess HDAC-inhibiting properties. It belongs to the hydroxymates, targeting both class I and class II HDACs. In the brain, tissue specific effects have been reported for TSA on BDNF expression. In the hippocampus TSA induces transcription of promoter exon 1 but not of exon 4 of the BDNF gene, associated with hyperacetylation at H3K9 and H3K14 and increased BDNF protein levels. At the same time an increase in HDAC mRNA and protein levels was also observed, suggesting a compensatory mechanism in response to HDAC inhibition.Citation76 In astrocytes TSA upregulates both BDNF and GDNF mRNA levels, with marked increases in GDNF promoter activity and promoter-associated histone H3 acetylation.Citation51 However, in glioma cells TSA does not induce any change in the basal levels of BDNF mRNA although TSA did strengthen the positive influence of serotonin on BDNF transcription in these cells,Citation53 suggesting that TSA may enhance BDNF expression in glia through serotonin-mediated pathways.
TSA also induced transcription of depression associated genes: the melatonin MT1 receptor gene in glioma cellsCitation65 and the Glucocorticoid Receptor (GR) gene.Citation77 In a study on the influence of maternal care on GR expression, Weaver and colleagues suggested that poor mothering lowers GR mRNA levels through methylation of the NGFI-A binding site. This interferes with NGFI-A binding to the GR exon 1 promoter, inhibiting expression.Citation77 In adulthood, TSA reversed the epigenetic effects of poor maternal care. It increased hippocampal GR expression by increasing H3K9 acetylation, DNA demethylation and binding of NGFI-A to the GR exon 1 promoter, to levels comparable to rats that received normal fostering and reversed the behavioral effects of poor fostering.Citation78 They found that over 900 genes were regulated by maternal fostering behavior. Of these, less than 2% were affected by TSA treatment but included genes that have been implicated in learning and memory, X-linked mental retardation and neurodegenerative disorders. TSA administration also accelerated fear extinction by 8-fold in a contextual fear paradigm.Citation71
Alteration of transcription of glutamate related genes may provide a functional link to psychosis. In astrocytes, TSA decreased DNA methylation of the GLT-1 gene leading to a 4-fold increase in GLT-1 mRNA.Citation29 Similar to VPA, TSA increases mRNA levels of the reelin and GAD67 genes in neural precursor and neural blastoma cells. By increasing acetylation of H3 and H4 and decreasing expression and enzymatic activity of DNMT1 and 3A and B, the reelin promoter is demethylated and more available for transcription.Citation55,Citation60,Citation61 The influence of TSA was also investigated in human lymphocytes. It induced an increase of nearly 4-fold in GAD67 mRNA levels, an almost 2-fold increase in acetylation of H3K9 and K14, and a 5-fold rise in attachment of acetylated H3K9 and K14 to the GAD67 promoter.Citation56
MS-275
MS-275 is a benzamide-based HDAC inhibitor that selectively targets class I HDACs, but it is not as popular in scientific research as the previous compounds. In mouse models for depression, infusion of MS-275 into the nucleus accumbens delivered strong antidepressant-like effects. In these mice, chronic defeat stress led to a unique global pattern of gene transcription in the nucleus accumbens which could be reversed by MS-275 treatment. The effects of MS-275 on gene expression were compared with the effects of fluoxetine, in search for novel antidepressant targets. A number of genes were only regulated by MS-275, including genes involved in gap junction formation and in adrenergic receptor function.Citation79 In addition, MS-275 increased mRNA levels of reelin and GAD67 in neural precursor and neural blastoma cells. By decreasing expression and enzymatic activity of DNMTs, the reelin promoter is demethylated and more available for transcription.Citation55 Simonini and colleaguesCitation47 reported that MS-275 increased acetylation of histone H3 interacting with reelin and Gad67 gene promoters preferentially in the frontal cortices of mice. However, much higher dose of MS-275 were needed to have an effect in the hippocampus and the striatum, indicating different sensitivity to MS-275 across different regions of the brain.
Treatment with MS-275 stimulated promoter demethylation and histone H3 acetylation in reelin and GAD67 in the cortex and hippocampus of miceCitation62 and reversed defeat induced behavior.Citation79 In neural progenitor cells MS-275 dose-dependently decreased methylation at the reelin and GAD67 promoters as well, and it upregulated expression of reelin and GAD67, correlating with the extent of inhibition of HDAC activity.Citation55 MS-275 also downregulated DNMT1, DNMT3A and DNMT3B protein levels. These DNMTs, together with MeCP2 and HDAC1 appear to constitute gene-specific repressor complexes at the reelin and GAD67 promoters, as activation of these genes by MS-275 is associated with dissociation of these proteins from the promoter regions. In addition, MS-275 decreased expression of MeCP2 and HDAC1. The synchronic involvement of DNMTs, MeCP2 and HDAC1 again show that DNA methylation and histone acetylation are intertwined mechanisms, controlling gene expression in synchrony.Citation55
DNMT inhibitors
DNMTs inhibitors are predominantly used to investigate the functional characteristics of DNMTs butrecent studies suggest that DNMT activity is important in social behavior of mice.Citation80 The compounds 5-aza-dC (AZA), zebularine and doxorubicin inhibit DNMT1 and DNMT3 and decrease DNA methylation of the reelin promoter in neural progenitor cells. This dramatically increased reelin and GAD67 mRNA levels, showing that the expression of the reelin and GAD67 genes was mediated by DNMTs. In addition, these DNMT inhibitors reduced DNMT enzymatic activity, decreased DNMT1 protein levels and increased H3 acetylation in the promoter area.Citation60,Citation61,Citation81 Treating hippocampal neurons with AZA leads to increased BDNF exon 4 mRNA, indicating that this splice variant is methylated by DNMT1 or DNMT3.Citation82 These effects were replicated using systemic administration of AZA leading to dose-dependent antidepressant-like effect, followed by decreased DNA methylation and increased BDNF levels, in the hippocampus.Citation83
Limitations and recommendations
Current epigenetic research in psychiatry is primarily focused on a subset of mechanisms. In most studies, histone acetylation, methylation, phosphorylation and DNA methylation are investigated, as these mechanisms are best characterized and measurable with techniques such as chromatin immunoprecipitation (ChIP). Moreover these epigenetic mechanisms are generally studied in isolation despite mounting evidence that neither histone modifications nor DNA methylation act independently. It would therefore be desirable to measure these different levels of epigenetic modification simultaneously, to get better insight into relevant epigenetic interactions. The current state of evidence is further obscured by the fact that the effects several epigenetic mechanisms such as ubiquitination, sumoylation, RNA- and polycomb-based mechanisms are not fully understood. Despite that they have not been well characterized in the brain they are likely to be important in epigenetic regulation as well. Future understanding these mechanisms would improve epigenetic psychiatric research.
Conclusion
gives an overview of the epigenetic mechanisms influenced by know and potential future psychopharmacological drug. The most prominent (but not all) epigenetic influences of psychiatric drugs are depicted in . Compelling evidence from animal and human research indicate that some antidepressants most notably imipramine as well as some antipsychotics particularly clozapine and the mood stabilizer valproic acid alter epigenetic regulation of a variety of genes. In addition new compounds such as sodium butyrate and trichostatin A show some promise for future development of epigenetic psychopharmacology. Considering that the first converging evidence of behavioral and pharmacological efficacy of epigenetic compounds is emerging, the expectation is that further combined behavioral and pharmacological studies can pioneer this promising field. Although our understanding of the complex epigenetic regulatory mechanism is in the early stages, the preliminary evidence as presented here suggest we could be standing at the dawn of the development of epigenetic therapy for psychiatric disorders.
Table 1. Known epigenetic mechanisms of current and potential psychotropic drugs
Acknowledgments
We thank Ingrid Janssen for drawing the figure. M.B. is supported by a NARSAD young investigator grant. R.A.O. gratefully acknowledges support of NIMH grant RO1 MG078075.
Related Research Data
References
- Van, Speybroeck L. From epigenesis to epigenetics: the case of C. H. Waddington. Ann N Y Acad Sci 2002; 981:61 - 81; PMID: 12547674
- Jaenisch R, Bird A. Epigenetic regulation of gene expression: how the genome integrates intrinsic and environmental signals. Nat Genet 2003; 33:Suppl 245 - 54; http://dx.doi.org/10.1038/ng1089; PMID: 12610534
- Mikaelsson MA, Miller CA. DNA methylation: a transcriptional mechanism co-opted by the developed mammalian brain?. Epigenetics 2011; 6:548 - 51; http://dx.doi.org/10.4161/epi.6.5.15679; PMID: 21527830
- Mill J, Tang T, Kaminsky Z, Khare T, Yazdanpanah S, Bouchard L, et al. Epigenomic profiling reveals DNA-methylation changes associated with major psychosis. Am J Hum Genet 2008; 82:696 - 711; http://dx.doi.org/10.1016/j.ajhg.2008.01.008; PMID: 18319075
- Kuwabara T, Hsieh J, Nakashima K, Taira K, Gage FH. A small modulatory dsRNA specifies the fate of adult neural stem cells. Cell 2004; 116:779 - 93; http://dx.doi.org/10.1016/S0092-8674(04)00248-X; PMID: 15035981
- Zhao X, Ueba T, Christie BR, Barkho B, McConnell MJ, Nakashima K, et al. Mice lacking methyl-CpG binding protein 1 have deficits in adult neurogenesis and hippocampal function. Proc Natl Acad Sci USA 2003; 100:6777 - 82; http://dx.doi.org/10.1073/pnas.1131928100; PMID: 12748381
- Fan G, Beard C, Chen RZ, Csankovszki G, Sun Y, Siniaia M, et al. DNA hypomethylation perturbs the function and survival of CNS neurons in postnatal animals. J Neurosci 2001; 21:788 - 97; PMID: 11157065
- Wu JI, Lessard J, Olave IA, Qiu Z, Ghosh A, Graef IA, et al. Regulation of dendritic development by neuron-specific chromatin remodeling complexes. Neuron 2007; 56:94 - 108; http://dx.doi.org/10.1016/j.neuron.2007.08.021; PMID: 17920018
- Heijmans BT, Tobi EW, Stein AD, Putter H, Blauw GJ, Susser ES, et al. Persistent epigenetic differences associated with prenatal exposure to famine in humans. Proc Natl Acad Sci USA 2008; 105:17046 - 9; http://dx.doi.org/10.1073/pnas.0806560105; PMID: 18955703
- Weaver IC, Cervoni N, Champagne FA, D'Alessio AC, Sharma S, Seckl JR, et al. Epigenetic programming by maternal behavior. Nat Neurosci 2004; 7:847 - 54; http://dx.doi.org/10.1038/nn1276; PMID: 15220929
- Bateson P, Barker D, Clutton-Brock T, Deb D, D'Udine B, Foley RA, et al. Developmental plasticity and human health. Nature 2004; 430:419 - 21; http://dx.doi.org/10.1038/nature02725; PMID: 15269759
- Rutten BP, Mill J. Epigenetic mediation of environmental influences in major psychotic disorders. Schizophr Bull 2009; 35:1045 - 56; http://dx.doi.org/10.1093/schbul/sbp104; PMID: 19783603
- Mill J, Petronis A. Molecular studies of major depressive disorder: the epigenetic perspective. Mol Psychiatry 2007; 12:799 - 814; http://dx.doi.org/10.1038/sj.mp.4001992; PMID: 17420765
- Dolinoy DC, Weidman JR, Jirtle RL. Epigenetic gene regulation: linking early developmental environment to adult disease. Reprod Toxicol 2007; 23:297 - 307; http://dx.doi.org/10.1016/j.reprotox.2006.08.012; PMID: 17046196
- Gibney ER, Nolan CM. Epigenetics and gene expression. Heredity 2010; 105:4 - 13; http://dx.doi.org/10.1038/hdy.2010.54; PMID: 20461105
- Sleiman SF, Basso M, Mahishi L, Kozikowski AP, Donohoe ME, Langley B, et al. Putting the 'HAT' back on survival signalling: the promises and challenges of HDAC inhibition in the treatment of neurological conditions. Expert Opin Investig Drugs 2009; 18:573 - 84; http://dx.doi.org/10.1517/13543780902810345; PMID: 19388875
- Hahnen E, Hauke J, Trankle C, Eyupoglu IY, Wirth B, Blumcke I. Histone deacetylase inhibitors: possible implications for neurodegenerative disorders. Expert Opin Investig Drugs 2008; 17:169 - 84; http://dx.doi.org/10.1517/13543784.17.2.169; PMID: 18230051
- Kouzarides T. Chromatin modifications and their function. Cell 2007; 128:693 - 705; http://dx.doi.org/10.1016/j.cell.2007.02.005; PMID: 17320507
- Mosammaparast N, Shi Y. Reversal of histone methylation: biochemical and molecular mechanisms of histone demethylases. Annu Rev Biochem 2010; 79:155 - 79; http://dx.doi.org/10.1146/annurev.biochem.78.070907.103946; PMID: 20373914
- Ebert A, Lein S, Schotta G, Reuter G. Histone modification and the control of heterochromatic gene silencing in Drosophila. Chromosome Res 2006; 14:377 - 92; http://dx.doi.org/10.1007/s10577-006-1066-1; PMID: 16821134
- Renthal W, Nestler EJ. Chromatin regulation in drug addiction and depression. Dialogues Clin Neurosci 2009; 11:257 - 68; PMID: 19877494
- Kriaucionis S, Heintz N. The nuclear DNA base 5-hydroxymethylcytosine is present in Purkinje neurons and the brain. Science 2009; 324:929 - 30; http://dx.doi.org/10.1126/science.1169786; PMID: 19372393
- Globisch D, Munzel M, Muller M, Michalakis S, Wagner M, Koch S, et al. Tissue distribution of 5-hydroxymethylcytosine and search for active demethylation intermediates. PLoS ONE 2010; 5:e15367; http://dx.doi.org/10.1371/journal.pone.0015367; PMID: 21203455
- Maunakea AK, Nagarajan RP, Bilenky M, Ballinger TJ, D'Souza C, Fouse SD, et al. Conserved role of intragenic DNA methylation in regulating alternative promoters. Nature 2010; 466:253 - 7; http://dx.doi.org/10.1038/nature09165; PMID: 20613842
- Deaton AM, Webb S, Kerr AR, Illingworth RS, Guy J, Andrews R, et al. Cell type-specific DNA methylation at intragenic CpG islands in the immune system. Genome Res 2011; 21:1074 - 86; http://dx.doi.org/10.1101/gr.118703.110; PMID: 21628449
- Thomson JP, Skene PJ, Selfridge J, Clouaire T, Guy J, Webb S, et al. CpG islands influence chromatin structure via the CpG-binding protein Cfp1. Nature 2010; 464:1082 - 6; http://dx.doi.org/10.1038/nature08924; PMID: 20393567
- Tsankova NM, Berton O, Renthal W, Kumar A, Neve RL, Nestler EJ. Sustained hippocampal chromatin regulation in a mouse model of depression and antidepressant action. Nat Neurosci 2006; 9:519 - 25; http://dx.doi.org/10.1038/nn1659; PMID: 16501568
- Wilkinson MB, Xiao G, Kumar A, LaPlant Q, Renthal W, Sikder D, et al. Imipramine treatment and resiliency exhibit similar chromatin regulation in the mouse nucleus accumbens in depression models. J Neurosci 2009; 29:7820 - 32; http://dx.doi.org/10.1523/JNEUROSCI.0932-09.2009; PMID: 19535594
- Perisic T, Zimmermann N, Kirmeier T, Asmus M, Tuorto F, Uhr M, et al. Valproate and amitriptyline exert common and divergent influences on global and gene promoter-specific chromatin modifications in rat primary astrocytes. Neuropsychopharmacology 2010; 35:792 - 805; http://dx.doi.org/10.1038/npp.2009.188; PMID: 19924110
- Mao X, Hou T, Cao B, Wang W, Li Z, Chen S, et al. The tricyclic antidepressant amitriptyline inhibits D-cyclin transactivation and induces myeloma cell apoptosis by inhibiting histone deacetylases: in vitro and in silico evidence. Mol Pharmacol 2011; 79:672 - 80; http://dx.doi.org/10.1124/mol.110.068122; PMID: 21220410
- Hunter RG, McCarthy KJ, Milne TA, Pfaff DW, McEwen BS. Regulation of hippocampal H3 histone methylation by acute and chronic stress. Proc Natl Acad Sci U S A. 2009.
- Karpova NN, Lindholm J, Pruunsild P, Timmusk T, Castren E. Long-lasting behavioural and molecular alterations induced by early postnatal fluoxetine exposure are restored by chronic fluoxetine treatment in adult mice. Eur Neuropsychopharmacol 2009; 19:97 - 108; http://dx.doi.org/10.1016/j.euroneuro.2008.09.002; PMID: 18973993
- Cassel S, Carouge D, Gensburger C, Anglard P, Burgun C, Dietrich JB, et al. Fluoxetine and cocaine induce the epigenetic factors MeCP2 and MBD1 in adult rat brain. Mol Pharmacol 2006; 70:487 - 92; http://dx.doi.org/10.1124/mol.106.022301; PMID: 16670375
- Covington HE III, Vialou VF, LaPlant Q, Ohnishi YN, Nestler EJ. Hippocampal-dependent antidepressant-like activity of histone deacetylase inhibition. Neurosci Lett 2011; 493:122 - 6; http://dx.doi.org/10.1016/j.neulet.2011.02.022; PMID: 21335060
- Melas PA, Rogdaki M, Lennartsson A, Bjork K, Qi H, Witasp A, et al. Antidepressant treatment is associated with epigenetic alterations in the promoter of P11 in a genetic model of depression. Int J Neuropsychopharmacol 2011; 1 - 11; http://dx.doi.org/10.1017/S1461145711000940; PMID: 21682946
- Culhane JC, Wang D, Yen PM, Cole PA. Comparative analysis of small molecules and histone substrate analogues as LSD1 lysine demethylase inhibitors. J Am Chem Soc 2010; 132:3164 - 76; http://dx.doi.org/10.1021/ja909996p; PMID: 20148560
- Binda C, Valente S, Romanenghi M, Pilotto S, Cirilli R, Karytinos A, et al. Biochemical, structural, and biological evaluation of tranylcypromine derivatives as inhibitors of histone demethylases LSD1 and LSD2. J Am Chem Soc 2010; 132:6827 - 33; http://dx.doi.org/10.1021/ja101557k; PMID: 20415477
- Lee MG, Wynder C, Schmidt DM, McCafferty DG, Shiekhattar R. Histone H3 lysine 4 demethylation is a target of nonselective antidepressive medications. Chem Biol 2006; 13:563 - 7; http://dx.doi.org/10.1016/j.chembiol.2006.05.004; PMID: 16793513
- Li J, Guo Y, Schroeder FA, Youngs RM, Schmidt TW, Ferris C, et al. Dopamine D2-like antagonists induce chromatin remodeling in striatal neurons through cyclic AMP-protein kinase A and NMDA receptor signaling. J Neurochem 2004; 90:1117 - 31; http://dx.doi.org/10.1111/j.1471-4159.2004.02569.x; PMID: 15312167
- Bertran-Gonzalez J, Bosch C, Maroteaux M, Matamales M, Herve D, Valjent E, et al. Opposing patterns of signaling activation in dopamine D1 and D2 receptor-expressing striatal neurons in response to cocaine and haloperidol. J Neurosci 2008; 28:5671 - 85; http://dx.doi.org/10.1523/JNEUROSCI.1039-08.2008; PMID: 18509028
- Bertran-Gonzalez J, Hakansson K, Borgkvist A, Irinopoulou T, Brami-Cherrier K, Usiello A, et al. Histone H3 phosphorylation is under the opposite tonic control of dopamine D2 and adenosine A2A receptors in striatopallidal neurons. Neuropsychopharmacology 2009; 34:1710 - 20; http://dx.doi.org/10.1038/npp.2008.228; PMID: 19158668
- Shimabukuro M, Jinno Y, Fuke C, Okazaki Y. Haloperidol treatment induces tissue- and sex-specific changes in DNA methylation: a control study using rats. Behav Brain Funct 2006; 2:37; http://dx.doi.org/10.1186/1744-9081-2-37; PMID: 17132176
- Dong E, Nelson M, Grayson DR, Costa E, Guidotti A. Clozapine and sulpiride but not haloperidol or olanzapine activate brain DNA demethylation. Proc Natl Acad Sci USA 2008; 105:13614 - 9; http://dx.doi.org/10.1073/pnas.0805493105; PMID: 18757738
- Huang HS, Matevossian A, Whittle C, Kim SY, Schumacher A, Baker SP, et al. Prefrontal dysfunction in schizophrenia involves mixed-lineage leukemia 1-regulated histone methylation at GABAergic gene promoters. J Neurosci 2007; 27:11254 - 62; http://dx.doi.org/10.1523/JNEUROSCI.3272-07.2007; PMID: 17942719
- Guidotti A, Auta J, Chen Y, Davis JM, Dong E, Gavin DP, et al. Epigenetic GABAergic targets in schizophrenia and bipolar disorder. Neuropharmacology 2011; 60:1007 - 16; http://dx.doi.org/10.1016/j.neuropharm.2010.10.021; PMID: 21074545
- Matrisciano F, Dong E, Gavin DP, Nicoletti F, Guidotti A. Activation of Group II Metabotropic Glutamate Receptors Promotes DNA Demethylation in the Mouse Brain. Mol Pharmacol 2011; 80:174 - 82; http://dx.doi.org/10.1124/mol.110.070896; PMID: 21505039
- Simonini MV, Camargo LM, Dong E, Maloku E, Veldic M, Costa E, et al. The benzamide MS-275 is a potent, long-lasting brain region-selective inhibitor of histone deacetylases. Proc Natl Acad Sci USA 2006; 103:1587 - 92; http://dx.doi.org/10.1073/pnas.0510341103; PMID: 16432198
- Li J, Guo Y, Schroeder FA, Youngs RM, Schmidt TW, Ferris C, et al. Dopamine D2-like antagonists induce chromatin remodeling in striatal neurons through cyclic AMP-protein kinase A and NMDA receptor signaling. J Neurochem 2004; 90:1117 - 31; http://dx.doi.org/10.1111/j.1471-4159.2004.02569.x; PMID: 15312167
- D'Souza A, Onem E, Patel P, La Gamma EF, Nankova BB. Valproic acid regulates catecholaminergic pathways by concentration-dependent threshold effects on TH mRNA synthesis and degradation. Brain Res 2009; 1247:1 - 10; http://dx.doi.org/10.1016/j.brainres.2008.09.088; PMID: 18976638
- Fukuchi M, Nii T, Ishimaru N, Minamino A, Hara D, Takasaki I, et al. Valproic acid induces up- or down-regulation of gene expression responsible for the neuronal excitation and inhibition in rat cortical neurons through its epigenetic actions. Neurosci Res 2009; 65:35 - 43; http://dx.doi.org/10.1016/j.neures.2009.05.002; PMID: 19463867
- Wu X, Chen PS, Dallas S, Wilson B, Block ML, Wang CC, et al. Histone deacetylase inhibitors up-regulate astrocyte GDNF and BDNF gene transcription and protect dopaminergic neurons. Int J Neuropsychopharmacol 2008; 11:1123 - 34; http://dx.doi.org/10.1017/S1461145708009024; PMID: 18611290
- Yasuda S, Liang MH, Marinova Z, Yahyavi A, Chuang DM. The mood stabilizers lithium and valproate selectively activate the promoter IV of brain-derived neurotrophic factor in neurons. Mol Psychiatry 2009; 14:51 - 9; http://dx.doi.org/10.1038/sj.mp.4002099; PMID: 17925795
- Morita K, Gotohda T, Arimochi H, Lee MS, Her S. Histone deacetylase inhibitors promote neurosteroid-mediated cell differentiation and enhance serotonin-stimulated brain-derived neurotrophic factor gene expression in rat C6 glioma cells. J Neurosci Res 2009; 87:2608 - 14; http://dx.doi.org/10.1002/jnr.22072; PMID: 19360904
- Sharma RP, Rosen C, Kartan S, Guidotti A, Costa E, Grayson DR, et al. Valproic acid and chromatin remodeling in schizophrenia and bipolar disorder: preliminary results from a clinical population. Schizophr Res 2006; 88:227 - 31; http://dx.doi.org/10.1016/j.schres.2006.07.015; PMID: 16996718
- Kundakovic M, Chen Y, Guidotti A, Grayson DR. The reelin and GAD67 promoters are activated by epigenetic drugs that facilitate the disruption of local repressor complexes. Mol Pharmacol 2009; 75:342 - 54; http://dx.doi.org/10.1124/mol.108.051763; PMID: 19029285
- Gavin DP, Kartan S, Chase K, Jayaraman S, Sharma RP. Histone deacetylase inhibitors and candidate gene expression: An in vivo and in vitro approach to studying chromatin remodeling in a clinical population. J Psychiatr Res 2009; 43:870 - 6; http://dx.doi.org/10.1016/j.jpsychires.2008.12.006; PMID: 19187942
- Yu IT, Park JY, Kim SH, Lee JS, Kim YS, Son H. Valproic acid promotes neuronal differentiation by induction of proneural factors in association with H4 acetylation. Neuropharmacology 2009; 56:473 - 80; http://dx.doi.org/10.1016/j.neuropharm.2008.09.019; PMID: 19007798
- Guidotti A, Dong E, Kundakovic M, Satta R, Grayson DR, Costa E. Characterization of the action of antipsychotic subtypes on valproate-induced chromatin remodeling. Trends Pharmacol Sci 2009; 30:55 - 60; http://dx.doi.org/10.1016/j.tips.2008.10.010; PMID: 19110320
- Veldic M, Kadriu B, Maloku E, Agis-Balboa RC, Guidotti A, Davis JM, et al. Epigenetic mechanisms expressed in basal ganglia GABAergic neurons differentiate schizophrenia from bipolar disorder. Schizophr Res 2007; 91:51 - 61; http://dx.doi.org/10.1016/j.schres.2006.11.029; PMID: 17270400
- Chen Y, Sharma RP, Costa RH, Costa E, Grayson DR. On the epigenetic regulation of the human reelin promoter. Nucleic Acids Res 2002; 30:2930 - 9; http://dx.doi.org/10.1093/nar/gkf401; PMID: 12087179
- Mitchell CP, Chen Y, Kundakovic M, Costa E, Grayson DR. Histone deacetylase inhibitors decrease reelin promoter methylation in vitro. J Neurochem 2005; 93:483 - 92; http://dx.doi.org/10.1111/j.1471-4159.2005.03040.x; PMID: 15816871
- Dong E, Guidotti A, Grayson DR, Costa E. Histone hyperacetylation induces demethylation of reelin and 67-kDa glutamic acid decarboxylase promoters. Proc Natl Acad Sci USA 2007; 104:4676 - 81; http://dx.doi.org/10.1073/pnas.0700529104; PMID: 17360583
- Dong E, Agis-Balboa RC, Simonini MV, Grayson DR, Costa E, Guidotti A. Reelin and glutamic acid decarboxylase67 promoter remodeling in an epigenetic methionine-induced mouse model of schizophrenia. Proc Natl Acad Sci USA 2005; 102:12578 - 83; http://dx.doi.org/10.1073/pnas.0505394102; PMID: 16113080
- Tremolizzo L, Doueiri MS, Dong E, Grayson DR, Davis J, Pinna G, et al. Valproate corrects the schizophrenia-like epigenetic behavioral modifications induced by methionine in mice. Biol Psychiatry 2005; 57:500 - 9; http://dx.doi.org/10.1016/j.biopsych.2004.11.046; PMID: 15737665
- Kim B, Rincon Castro LM, Jawed S, Niles LP. Clinically relevant concentrations of valproic acid modulate melatonin MT(1) receptor, HDAC and MeCP2 mRNA expression in C6 glioma cells. Eur J Pharmacol 2008; 589:45 - 8; http://dx.doi.org/10.1016/j.ejphar.2008.04.058; PMID: 18550052
- Shen HY, Kalda A, Yu L, Ferrara J, Zhu J, Chen JF. Additive effects of histone deacetylase inhibitors and amphetamine on histone H4 acetylation, cAMP responsive element binding protein phosphorylation and DeltaFosB expression in the striatum and locomotor sensitization in mice. Neuroscience 2008; 157:644 - 55; http://dx.doi.org/10.1016/j.neuroscience.2008.09.019; PMID: 18848971
- Dickinson M, Johnstone RW, Prince HM. Histone deacetylase inhibitors: potential targets responsible for their anti-cancer effect. Invest New Drugs 2010; 28:Suppl 1 S3 - 20; http://dx.doi.org/10.1007/s10637-010-9596-y; PMID: 21161327
- Davie JR. Inhibition of histone deacetylase activity by butyrate. J Nutr 2003; 133:2485S - 93S; PMID: 12840228
- Stefanko DP, Barrett RM, Ly AR, Reolon GK, Wood MA. Modulation of long-term memory for object recognition via HDAC inhibition. Proc Natl Acad Sci USA 2009; 106:9447 - 52; http://dx.doi.org/10.1073/pnas.0903964106; PMID: 19470462
- Gupta S, Kim SY, Artis S, Molfese DL, Schumacher A, Sweatt JD, et al. Histone methylation regulates memory formation. J Neurosci 2010; 30:3589 - 99; http://dx.doi.org/10.1523/JNEUROSCI.3732-09.2010; PMID: 20219993
- Lattal KM, Barrett RM, Wood MA. Systemic or intrahippocampal delivery of histone deacetylase inhibitors facilitates fear extinction. Behav Neurosci 2007; 121:1125 - 31; http://dx.doi.org/10.1037/0735-7044.121.5.1125; PMID: 17907845
- Schroeder FA, Lin CL, Crusio WE, Akbarian S. Antidepressant-like effects of the histone deacetylase inhibitor, sodium butyrate, in the mouse. Biol Psychiatry 2007; 62:55 - 64; http://dx.doi.org/10.1016/j.biopsych.2006.06.036; PMID: 16945350
- Gundersen BB, Blendy JA. Effects of the histone deacetylase inhibitor sodium butyrate in models of depression and anxiety. Neuropharmacology 2009; 57:67 - 74; http://dx.doi.org/10.1016/j.neuropharm.2009.04.008; PMID: 19393671
- Kwon B, Houpt TA. Phospho-acetylation of histone H3 in the amygdala after acute lithium chloride. Brain Res 2010; 1333:36 - 47; http://dx.doi.org/10.1016/j.brainres.2010.03.068; PMID: 20346924
- Patel P, Nankova BB, LaGamma EF. Butyrate, a gut-derived environmental signal, regulates tyrosine hydroxylase gene expression via a novel promoter element. Brain Res Dev Brain Res 2005; 160:53 - 62; http://dx.doi.org/10.1016/j.devbrainres.2005.08.005; PMID: 16165221
- Tian F, Marini AM, Lipsky RH. Effects of histone deacetylase inhibitor Trichostatin A on epigenetic changes and transcriptional activation of Bdnf promoter 1 by rat hippocampal neurons. Ann N Y Acad Sci 2010; 1199:186 - 93; http://dx.doi.org/10.1111/j.1749-6632.2009.05175.x; PMID: 20633124
- Weaver IC, D'Alessio AC, Brown SE, Hellstrom IC, Dymov S, Sharma S, et al. The transcription factor nerve growth factor-inducible protein a mediates epigenetic programming: altering epigenetic marks by immediate-early genes. J Neurosci 2007; 27:1756 - 68; http://dx.doi.org/10.1523/JNEUROSCI.4164-06.2007; PMID: 17301183
- Weaver IC, Meaney MJ, Szyf M. Maternal care effects on the hippocampal transcriptome and anxiety-mediated behaviors in the offspring that are reversible in adulthood. Proc Natl Acad Sci USA 2006; 103:3480 - 5; http://dx.doi.org/10.1073/pnas.0507526103; PMID: 16484373
- Covington HE 3rd, Maze I, LaPlant QC, Vialou VF, Ohnishi YN, Berton O, et al. Antidepressant actions of histone deacetylase inhibitors. J Neurosci 2009; 29:11451 - 60; http://dx.doi.org/10.1523/JNEUROSCI.1758-09.2009; PMID: 19759294
- Sales AJ, Biojone C, Terceti MS, Guimaraes FS, Gomes MV, Joca SR. Antidepressant-like effect induced by systemic and intra-hippocampal administration of DNA methylation inhibitors. Br J Pharmacol 2011; 164:1711 - 21; http://dx.doi.org/10.1111/j.1476-5381.2011.01489.x; PMID: 21585346
- Kundakovic M, Chen Y, Costa E, Grayson DR. DNA methyltransferase inhibitors coordinately induce expression of the human reelin and glutamic acid decarboxylase 67 genes. Mol Pharmacol 2007; 71:644 - 53; http://dx.doi.org/10.1124/mol.106.030635; PMID: 17065238
- Tian F, Hu XZ, Wu X, Jiang H, Pan H, Marini AM, et al. Dynamic chromatin remodeling events in hippocampal neurons are associated with NMDA receptor-mediated activation of Bdnf gene promoter 1. J Neurochem 2009; 109:1375 - 88; http://dx.doi.org/10.1111/j.1471-4159.2009.06058.x; PMID: 19476549
- Sales AJ, Biojone C, Terceti MS, Guimaraes FS, Gomes MV, Joca SR. Antidepressant-like effect induced by systemic and intra-hippocampal administration of DNA methylation inhibitors. Br J Pharmacol 2011; 164:1711 - 21; http://dx.doi.org/10.1111/j.1476-5381.2011.01489.x; PMID: 21585346