
Abstract
Probiotics facilitate mucosal repair and maintain gut homeostasis. They are often used in adjunct with rehydration or antibiotic therapy in enteric infections. Lactobacillus spp have been tested in infants for the prevention or treatment of various enteric conditions. However, to aid in rational strain selection for specific treatments, comprehensive studies are required to delineate and compare the specific molecules and pathways involved in a less complex but biologically relevant model (gnotobiotic pigs). Here we elucidated Lactobacillus rhamnosus (LGG) and L. acidophilus (LA) specific effects on gut transcriptome responses in a neonatal gnotobiotic (Gn) pig model to simulate responses in newly colonized infants. Whole genome microarray, followed by biological pathway reconstruction, was used to investigate the host-microbe interactions in duodenum and ileum at early (day 1) and later stages (day 7) of colonization. Both LA and LGG modulated common responses related to host metabolism, gut integrity, and immunity, as well as responses unique to each strain in Gn pigs. Our data indicated that probiotic establishment and beneficial effects in the host are guided by: (1) down-regulation or upregulation of immune function-related genes in the early and later stages of colonization, respectively, and (2) alternations in metabolism of small molecules (vitamins and/or minerals) and macromolecules (carbohydrates, proteins, and lipids). Pathways related to immune modulation and carbohydrate metabolism were more affected by LGG, whereas energy and lipid metabolism-related transcriptome responses were prominently modulated by LA. These findings imply that identification of probiotic strain-specific gut responses could facilitate the rational design of probiotic-based interventions to moderate specific enteric conditions.
Introduction
Enteric pathogens cause diarrheal diseases in both adults and infants, with the latter being more susceptible due to an immature gut with increased permeability to antigens, as well as an immature immune system.Citation1,Citation2 Given this scenario, particularly in developing countries where availability of enteric vaccines or lower efficacy of vaccines (e.g., rotavirus vaccine) are problems, other means of treatment in addition to oral rehydration are often required. Probiotics that facilitate mucosal repair and maintain gut homeostasis are invaluable treatment modalities for neonatal enteric infections including rotavirus diarrhea.Citation3 Recent research has highlighted the effective role of probiotics in modulation of the gut microbiota, thereby alleviating intestinal disorders like inflammatory bowel disease, travelers’ diarrhea, colitis, Crohn disease, and antibiotic associated diarrhea.Citation4,Citation5 However, pathways and mechanisms by which probiotics mediate their beneficial effects, especially in neonates, are largely undefined.
Among several probiotic bacteria, Gram-positive bacteria dominate, especially ones belonging to the Lactobacillus and Bifidobacterium genera. Lactobacillus species are probably the most widely consumed and marketed in the world because of the long history of safety in the food and dairy industry.Citation6 However, overall they comprise only a small portion of the natural gut microflora.Citation7 Many Lactobacillus spp. have been tested for the prevention or treatment of various pathological conditions. Numerous clinical studies have shown the beneficial role of Lactobacillus rhamnosus GG (LGG) in treatment of acute diarrhea in pediatric patients.Citation8-Citation10 LGG was effective in decreasing both duration and frequency of rotaviral diarrhea and also antibiotic-associated diarrhea in neonates.Citation11-Citation13 People consuming L. acidophilus (LA) NCFM had reduced fever, cough, and runny nose.Citation14 Additionally, LA strains have also been shown to possess anti-inflammatory and breast cancer inhibition effects.Citation14 These studies demonstrate that probiotics mediate their effects in a species and strain specific manner; however, mechanisms are still largely undefined.Citation15
The Lactobacillus strains exert their beneficial effects in multiple ways: producing antimicrobial substances, enhancing barrier functions (i.e., improved gut integrity), producing cryoprotective heat shock protein (Hsp72) and antiapoptotic proteins (p40 and p75), regulating Th1-Th2 balance, enhancing B cell and IFN responses, and inhibiting cytokine induced apoptosis.Citation16-Citation22 Additionally, the type and timing of colonization of Lactobacillus strains influence the early neonatal gut mucosal immune responses and have a significant impact on resistance to diseases postnatally; however, further studies are required to understand the underlying molecular mechanisms. Investigations into these basic mechanisms will identify specific molecules and pathways mediating the beneficial effects on the host, which in turn aid in probiotic strain selection to modulate the specific clinical conditions.
Investigation of in vivo probiotic-host cell interactions is difficult considering the complex gut microflora in infants. Germ-free animal models are critical to dissect the molecular interactions of probiotics. The neonatal piglets are similar to infants in terms of anatomy, physiology, and mucosal immunology and serve as a useful model to elucidate mechanisms of probiotic modulation of neonatal host responses.Citation23 In the present study, we used neonatal Gnotobiotic (Gn) pig model to better understand L. rhamnosus (LGG) and L. acidophilus (LA) strain-specific responses. In vivo strain-specific colonization dynamics were investigated by conventional bacterial culturing and host-microbe interactions by in vivo transcriptome analysis of the small intestine (SI). We used duodenum and ileum for transcriptome analyses because they represent both inductive (ileum) and effector (duodenum) mucosal sites to assess probiotic-host interactions. The duodenum is the first segment of the SI to come in contact with probiotic bacteria, and it also helps in minimizing adaptive changes that probiotics might go through during passage of the intestinal tract.Citation24 The ileum contains specialized structures (Peyer patches) for sampling and induction of immune responses to antigens and is also the site of enteropathogenic bacterial invasion.Citation25 We investigated both early (post-bacterial colonization day 1, PBCD1) and later (PBCD7) transcriptome responses to probiotics. Comprehensive analysis of our microarray data indicated that both LA and LGG modulated expression of genes regulating host metabolism, mucosal cell integrity, and immunity in neonatal Gn pigs. Immune modulation and carbohydrate metabolism genes were most affected in LGG, whereas energy and lipid metabolism genes were pronounced in LA-colonized Gn pigs. Collectively, our study provides new insights to the distinct and common cellular pathways modulated in Gn pigs by LA and LGG.
Results
Colonization begins at early stage (PBCD1) and increases at later stage (PBCD7)
On PBCD1, LA colonized duodenum (1.0 × 103 CFU/g), jejunum (9.4 × 102 CFU/g), and ileum (3.3 × 104 CFU/g) with higher CFU counts in the large intestine, reaching a maximum in the cecum (1.8 × 105 CFU/g). Surprisingly, LA was also detected in the spleen, liver, and MLNs in two of the three pigs. On the other hand, for LGG on PBCD1, the CFU counts were one log higher than LA in duodenum (2.6 × 104 CFU/g) and ileum (2.8 × 105 CFU/g), and the colonization in jejunum was similar to LA (1.5 × 102 CFU/g). Similarly LGG also colonized the large intestine in higher numbers than LA, with maximum colonization in the cecum (1.7 × 106 CFU/g). Unlike LA, no LGG was detected (detection limit, 10 CFU/g) in the extra-intestinal tissues.
On PBCD7 the LA colonization pattern remained similar to PBCD1, except more bacteria (>1-log) were recovered from all intestinal tissues and in higher numbers from all the extra-intestinal tissues. However, LGG colonization was similar to PBCD1 and was also recovered from spleen (1 pig), liver (3 pigs), and MLNs (1 pig) on PBCD7. Translocation of both LA and LGG to secondary lymphoid organs (spleen, MLN) may serve for development of appropriate protective and controlled immune responses in the host.Citation26,Citation27 Rectal swab cultures indicated that both LA and LGG shedding increased from PBCD1 (LA, 2.2 × 104 CFU/g; LGG, 8 × 104 CFU/g) to PBCD7 (LA, 6 × 106 CFU/g; LGG, 6.5 × 105 CFU/g) consistent with the increased intestinal colonization.
Ileum in early (PBCD1) and duodenum in later (PBCD7) stages are the major sites of transcriptome changes in response to probiotic colonization
A summary of transcriptome responses following probiotic administration is listed in . Comparison of differentially expressed transcripts revealed more changes in the ileum for PBCD1 and in the duodenum for PBCD7, irrespective of probiotic bacteria treatment. On PBCD1 LA modulated 286 genes in ileum, whereas LGG modulated 292 genes. On PBCD7 in duodenum, LA affected 137 genes, and in contrast, LGG affected 569 genes. Another trend noted for both lactobacilli strains at both times was that more genes were downregulated in duodenum and upregulated in ileum, with the exception of ileum for LGG at PBCD 7. The transcriptional changes observed by microarray analyses were further confirmed by RT-qPCR for selected genes (Fig. S1), which also indicated a similar overall trend in the transcription.
Table 1. Summary of gut transcriptome responses to LA and LGG*
The microarray-generated transcripts were further mapped to available gene annotations in the IPA. Only up to 55% of transcripts were mapped due to lack of complete pig genome annotation. The number of IPA-mapped genes was further compared across probiotic strains, tissues, and days post-colonization. On PBCD1 numbers of mapped genes were not significantly different across strains and tissues; however, significant differences were observed in duodenum on PBCD7 where LGG modulated higher numbers (2.1-fold) of unique as well common mapped genes compared to LA (Fig. S2; Table S1).
Cellular growth and proliferation, inflammatory response, and immune cell trafficking pathways were modulated by LA and LGG
To determine the relationship of genes identified in our microarray analysis using IPA, we compared the major well-defined molecular (canonical) pathways that were significantly affected by probiotic administration. Our results highlighted the distinct modulation of host gene expression in duodenum and ileum by both LA and LGG (). Specifically, in most categories, LGG induced a pronounced response in duodenum, particularly on PBCD7, whereas an opposite trend was observed with LA, which induced a pronounced response on several pathways in ileum on PBCD7. On PBCD1 in duodenum, LGG modulated pathways associated with inflammatory response, hematological system development, and immune cell trafficking; however, both LA and LGG modulated cell-to-cell signaling and interaction, cellular growth, and proliferation pathways. On PBCD7 LGG also modulated cellular growth and proliferation, inflammatory response, and immune cell trafficking, whereas both LA and LGG affected cell signaling and cellular growth and proliferation pathways. Contrastingly, on PBCD7 in ileum, LA-modulated host tissue development, cellular movement, immune cell trafficking, and cell-to-cell signaling pathways, while both LA and LGG modulated inflammatory response and free radical scavenging pathways. Collectively, our analyses highlight the unique and common major cellular pathways modulated in duodenum and ileum by LA and LGG.
Figure 1. Major cellular pathways (canonical pathways) modulated by LA and LGG were generated using ingenuity biological process analysis. These pathways were compared in duodenum and ileum for both days. The selected pathways were the significantly modulated ones and were above the threshold (-log [p value)] analyzed using IPA. Numbers on the top of the bars represent the total number of genes involved in a given canonical pathway. The distinction between probiotic strains was based on log p value and number of genes involved. (A) duodenum and (B) ileum.
![Figure 1. Major cellular pathways (canonical pathways) modulated by LA and LGG were generated using ingenuity biological process analysis. These pathways were compared in duodenum and ileum for both days. The selected pathways were the significantly modulated ones and were above the threshold (-log [p value)] analyzed using IPA. Numbers on the top of the bars represent the total number of genes involved in a given canonical pathway. The distinction between probiotic strains was based on log p value and number of genes involved. (A) duodenum and (B) ileum.](/cms/asset/f449e53a-5c65-4eef-95a8-cd80a8430ab1/kgmi_a_10927877_f0001.gif)
In the early stage at PBCD1, LA and LGG administration affected cell morphology, cellular movements, and anti-inflammatory genes in ileum
To better understand the biological relevance of transcriptome responses observed after probiotic colonization, we analyzed the networks resulting from these transcriptome responses using IPA. The top affected networks are shown in . The top network in each category was compared (LA vs. LGG) and further analyzed using “Molecular Activity Predictor” (MAP). The predicted activities of molecules were in turn linked to significant canonical pathways representing the major underlying biological processes. The gene regulatory networks with allied canonical pathways are depicted (; Figs. S3 and S4).
Table 2. Top gene regulatory networks associated with LA and LGG in duodenum and ileum based on focus molecules and scores*
Figure 2. Top associated networks with their linked canonical pathways (CP) in ileum on PBCD1. In the graphical representation of a network, genes or gene products are represented as nodes, and the biological relationship between two nodes is represented as an edge (line). Human, mouse, and rat orthologs of a gene are represented as a single node in the network. The intensity of the node color indicates the degree of up- (red) or down- (green) regulation, and more confidence predicted activation (blue) or inhibition (brown) of a given gene. Edges are displayed with various labels that describe the nature of the relationship between the nodes. Both LA and LGG were associated with glucocorticoid receptor (GR) signaling canonical pathway. The connecting sky blue line indicates the associated molecules with their predicted biological status, whereas the pink lines highlight the direct relation (solid line) between the genes. Only genes that are involved in canonical pathway are shown, and indirect interactions are depicted in dashed arrows with different colors representing the predicted biological status as indicated in the legend.
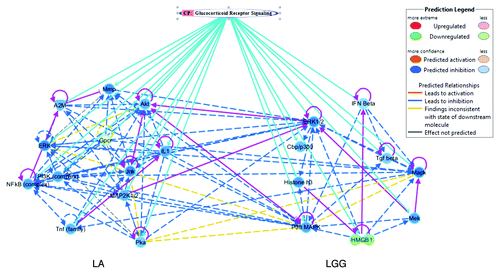
On PBCD1 the top network associated with LA in ileum was cellular movement, hematological system development and function, and immune cell trafficking. This network was linked to the canonical pathway involved in glucocorticoid receptor (GR) signaling. The majority of molecules involved (A2M, Akt, ERK, ERK1/2, GPCR, IL1, JNK, MAP2K1/2, MAPK, Mmp, NFκB [complex], P38 MAPK, PI3K [complex], Pka, and TNF [family]) were predicted to be inhibited (). MAPK, ERK1/2, and JNK have an important function in dynamic regulation of the cell cytoskeleton, tight junction (TJ), and epithelial barrier function.Citation28 Selective interaction of cytoskeleton protein networks provides a structural framework for cell morphology, stabilizes the other membrane systems, and mediates cellular movements.Citation29 This suggests that LA modulates the host cell morphology and cellular movement processes by selectively inhibiting these proteins. Further, glucocorticoids act through GR signaling and thereby mediate anti-inflammatory, anti-proliferative, and immunomodulatory activity.Citation30 Dampening of GR signaling by LA indicated probable early anti-inflammatory and immune modulatory responses in the ileum, permitting more effective colonization with higher CFU observed vs. duodenum.
Similarly, the top network associated with LGG in ileum was cell morphology, cellular function and maintenance, and cell cycle, which was also linked to the GR signaling canonical pathway (Cbp/p300, ERK1/2, Histone h3, HMGB1, IFNβ, MAPK, MEK, P38 MAPK, and TGFβ). The majority of the molecules involved in this pathway were predicted to be inhibited (). Cbp/p300 is a transcriptional co-activator protein that, besides exerting tumor-suppressive effects, also modulates proliferation, differentiation, and apoptosis in various cell types.Citation31 It is likely that inhibition of Cbp/p300 by LGG stimulates cell proliferation, differentiation, and anti-apoptosis. LGG also seems to mediate host anti-inflammatory effects like LA by modulating genes associated with GR signaling in ileum.
In the later stage at PBCD7, LA and LGG modulated cell signaling and metabolism genes in duodenum
On PBCD7 the top network associated with LA in duodenum was cell-to-cell signaling and interaction, tissue development, cell death, and survival. This network was associated with canonical pathway involved in integrin linked kinase (ILK) signaling (CDH1, ERK1/2, FOS, JUN, and VIM). The associated molecules were predicted to be inhibited (). The ILK signaling connects integrins to the cytoskeleton, thereby mediating cell signaling and also has important roles in cancer progression, and is a valid therapeutic target in cancer research.Citation32,Citation33 Downregulation of ILK signaling is likely to favor more colonization (> 1 × 10Citation5) vs. PBCD1. The inhibitory status of transcriptional factors such as FOS and JUN, which downregulates the release of inflammatory cytokine (anti-inflammatory effects),Citation34 is likely to decrease innate immunity to promote LA colonization.
Figure 3. Top associated networks with their linked canonical pathways in duodenum on PBCD7. LA was associated with integrin linked kinase (ILK) and LGG with thyroid receptor complex (TR/RXR) activation canonical pathways. The connecting sky blue line indicates the associated molecules with their predicted biological status, whereas the pink lines highlight the direct relation between the genes. Only genes that are involved in canonical pathway are shown, and indirect interactions are depicted in dashed arrows with different colors representing the predicted biological status as indicated in the legend.
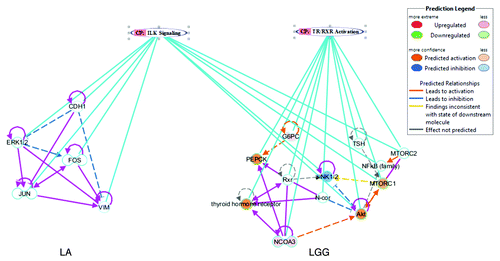
The top network associated with LGG in duodenum was cell cycle, carbohydrate metabolism, cellular function, and maintenance, which was linked to the canonical pathway involved in thyroid receptor complex (TR/RXR) activation signaling (). The corresponding molecules (Akt, G6PC, mTORC1, mTORC2, N-Cor, NCOA3, PEPCK, Rxr, thyroid hormone receptor, and TSH) were predicted to be activated except TSH (). Activation of TR/RXR signaling affects various processes including lipid/carbohydrate/steroid metabolism, thermogenesis, and CNS function.Citation35 In addition, functional Akt and mTORC1 complex promotes lipid anabolism in the liver,Citation36 and activation of G6PC (glucose-6-phosphatase), has consequences for host carbohydrate metabolism, as it is a key enzyme in glucose homeostasisCitation37 (; Table S2).
Table 3. Metabolism genes influenced by administration of probiotics LA and LGG in duodenum and ileum.*
The genes associated with metabolism, cellular integrity, and immune responses were either commonly or uniquely modulated by LA and LGG
Probiotic strain-specific host responses are well documented.Citation22,Citation25 The growth-phase-specific role for probiotic strains has also been reported recently.Citation24 In this regard, our data showed differentially expressed genes in different functional categories, particularly those involved in metabolism (; Table S2). Both LA and LGG modulated the genes involved in energy metabolism, whereas LA and LGG negatively regulated cytochrome subunits' genes (e.g., COX1) in duodenum and positively regulated ATPase subunits (e.g., ATP5F1) in ileum. Similarly, LGG downregulated the pyruvate kinase isozymes and phosphoglycerate mutase 2 (PGAM2) genes involved in glycolysis, but LA upregulated the gluconeogenesis gene phosphoenolpyruvate carboxykinase (PCK1). Conversely, in protein metabolism LA downregulated the 3-oxoacid CoA transferase 1 gene (OXCT1) involved in branched chain amino acid synthesis, whereas LGG upregulated the branched chain amino-acid transaminase 2 mitochondrial (BCAT2) gene also involved in branched chain amino acid synthesis. Genes involved in dietary fat metabolism (PNLIP) and cholesterol clearance (APOA1) were upregulated by both LA and LGG. LA and LGG also modulated genes involved in vitamin and mineral metabolism. A majority of genes were induced for vitamin A (e.g., RDH) and vitamin D (e.g., CYP24A1) metabolism. LA also induced the gene responsible for iron storage (FTH1).
Our data also supports common pathway modulation in duodenum and ileum: for example, the tight junction signaling on day 1, although the mechanistic regulation may vary for both LA and LGG due to involvement of different up stream regulators (Table S3). In duodenum transcription of catenin, claudin, and FOS molecules were affected where catenin was downregulated and claudin and FOS molecules were upregulated. In ileum, however, catenin, actin (α, αl and α2), FOS, and myosin (heavy and light chain) were involved, and all molecules were predicted to be activated except NF-κB. The NF-κB signaling inhibits MLCK (myosine light chain kinase) to decrease the permeability of tight junctions in the ileum (Fig. S5A and B), and this may suggest that MLCK inhibition may limit the extraintestinal dissemination of LGG to spleen, liver, and MLN on PBCD1.
Both LA and LGG induced genes involved in cellular integrity such as sialomucin (CD164 molecule), integrin, β 3 (ITGB3), heat shock 27 kDa protein 8 (Hsp27), and glucagon-like peptide-2 receptor (GLP2R) (; Table S4). Additionally, both LA and LGG tended to downregulate the innate immune response genes in the early stages (PBCD1) to favor colonization. For example, cytosolic- and membrane-bound forms of glutathione S transferase (GSTA1), lysozyme (LYZ), polymeric immunoglobulin receptors (PIGR), surfactant protein D (SFTPD), and an antibacterial protein regenerating islet-derived 3 gamma (REG3G) were downregulated in duodenum (; Table S4). However, in the later stages (PBCD7), complement component C9 and C8G, cellular innate mediators, CD1d, non-classical MHC I, and Th1-specific IFN-induced chemokine CXCL9, were induced by both LA and LGG.
Figure 4. Genes involved in cell integrity and immunity. The gene functions were determined using IPA and NCBI database and were manually compared with their regulated status. The genes involved were color coded to indicate regulated status (grey, downregulation; black, upregulation). The genes responsible for innate immunity were commonly regulated in both LA and LGG.

Discussion
The beneficial role of probiotics in nutritional physiology is less documented compared with its anti-diarrheal effects and role in immune stimulation.Citation38 However, few studies highlighted the beneficial role of lactobacilli in enhancing mineral and nutrient absorption, vitamin metabolism, or modulation of intestinal physiology.Citation39 Human interventional studies in children using L. rhamnosus and L. acidophilus showed improvement in weight and size of the subjects, likely due to an increased feed conversion ratio;Citation40,Citation41 however, mechanisms still remain unclear.Citation39 Our study provides mechanistic insights into the role of both LA and LGG in nutrient metabolism. LA upregulated two important host genes, ferritin (FTH1) and ceruloplasmin (Cp), which are involved in iron and copper homeostasis.Citation42 Furthermore, upregulation of antimicrobial peptide hepcidin (HAMP) and ferritin, the host iron sequestration genes, limit iron availability to intruding pathogens, thereby preventing pathogen establishment in the host.Citation43 Both LA and LGG may also play a role in bioavailability of small molecules, calcium, and phosphorus. Host intestinal absorption and homeostasis of calcium and phosphorus requires the vitamins A and D. The gene product of CYP2D25 and CYP24A1 encodes vitamin D 25-hydroxylase and 1, 25-dihydroxyvitamin D3 24-hydroxylase, respectively, the key enzymes in the vitamin D metabolism. Our data support the strain specific induction of CYP2D25 gene by LGG in duodenum on PBCD7 and common induction of CYP24A1 gene on PBCD1. It is interesting to note that LGG on PBCD1 differentially modulated the CYP24A1 gene in the duodenum and ileum, suggesting calcium absorption was favored in duodenum (; Table S2). Likewise, β-carotene 15,15'-monooxygenase (BCMO1) gene involved in vitamin A metabolism was upregulated by LA in duodenum (PBCD1); conversely, LGG in both tissues upregulated RDH16, retinol dehydrogenase 1, an enzyme involved in the visual cycle (; Table S2). Both vitamins A and D can modulate the innate and adaptive immune responses as well as metabolites of vitamins A and D can induce tissue-specific immune responses and have been investigated for preventing and/or treating inflammation and autoimmunity.Citation44
Previously, colonization of B. thetaiotaomicron in germ-free mice produced changes in host genes involved in breakdown and absorption of complex carbohydrates and lipids.Citation45 In our study lactobacilli administration altered expression of genes involved in glycolysis and/or gluconeogenesis. For example, pyruvate kinase isozymes R/L, like LOC100621940 and phosphoglycerate mutase 2 (PGAM2), were negatively regulated, whereas phosphoenolpyruvate carboxykinase 1 (PCK1) and glucose-6-phosphatase, catalytic subunit (G6PC) were positively regulated. This suggests a potential role for lactobacillus in host carbohydrate metabolism (). Lactobacillus contributes to host lipid metabolism through their ability to metabolize bile acids, the primary function of which is to emulsify fats. Hence, probiotics seem to regulate how much fat the body can absorb.Citation28 LA on PBCD1 in duodenum induced genes pancreatic lipase-related protein 1 (PNLIPRP1) (involved in breakdown of triacylglycerols) and pancreatic colipase (CLPS) (prevents the inhibitory effect of bile salts on lipase), whereas, on PBCD7 in duodenum, LGG upregulated pancreatic lipase (PNLIP), the primary enzyme required for hydrolyzing dietary fat. Recently, it was shown that yogurt containing probiotic bacterial strains, L. acidophilus and B. lactis, had a cholesterol-lowering effect in hypercholesterolemic subjects.Citation46 Both LA and LGG on PBCD7 in duodenum induced a 300-fold upregulation of apolipoprotein A-I (APOA1), a major protein component of high density lipoprotein (HDL) in plasma, promoting cholesterol efflux from tissues to the liver for excretion and helping to clear cholesterol from arteries.Citation47 L. casei Shirota and B. breveCitation48 have been shown to enhance colonic protein and ammonia metabolism in healthy humans. In our study genes involved in catabolism of branched chain amino acids (3-oxoacid CoA transferase 1 OXCT1), tyrosine (4-hydroxyphenylpyruvate dioxygenase HPD), and arginine (arginine decarboxylase SADC) were negatively regulated, whereas genes involved in amino acid synthesis (5-methyltetrahydrofolate-homocysteine methyltransferase reductase, MTRR) and branched chain amino acid synthesis (branched chain amino-acid transaminase 2, mitochondrial BCAT2) were positively regulated (). This suggests that both LA and LGG may play a role in host protein metabolism, and this may have been mediated through activation of TR/RXR signaling (), as in the case of LGG. TR/RXR signaling is attributed in metabolic signaling pathways (glucose, fatty acid, and cholesterol metabolism) besides morphogenesis, cell proliferation, cell differentiation, and cell death.Citation35
Alterations in the epithelial barrier functions are implicated in various intestinal disorders, and probiotics are suggested to enhance or maintain the epithelial barrier, but the molecules mediating these effects are only partially defined. In our study both LGG and LA altered gene expression in canonical pathways involving cell signaling, tissue development, cellular growth, and proliferation. For example, both LA and LGG ingestion enhanced expression of glucagon-like peptide-2 receptor (GLP2R) gene (). The GLP2R regulates intestinal growth stimulation and upregulation of villus height in the small intestine, concomitant with increased crypt cell proliferation and decreased enterocyte apoptosis. Enterocyte transcriptome profiling in neonatal mice has shown that ingestion of probiotic L. reuteri strains altered expression of genes associated with multiple canonical pathways involving cell motility, as well as increased enterocyte migration and proliferation in the ileum in a strain-specific manner.Citation22 ILK signaling stabilizes enterocyte migration and has been implicated in intestinal epithelial cell proliferation regulation and certain bacteria hijack ILK signaling to stabilize focal adhesions and prevent cell detachment.Citation49-Citation51 It is possible that LA might exploit ILK signaling to colonize the Gn pig intestine (). LGG also induced expression of claudin-8, which is associated with decreased paracellular gut permeabilityCitation52 (Table S3). Further, our results suggested an anti-apoptotic role for both LA and LGG on the host cell through induction of heat shock proteins (e.g., Hsp27) (). Increased expression of Hsp27 has been shown to suppress stress- and receptor-induced apoptotic pathways. Also, Hsp27 increases the antioxidant defense of cells by decreasing intracellular reactive oxygen species (ROS).Citation53 Thus, both LA and LGG may contribute to maintaining gut homeostasis; however, LGG may play a significant role as a probiotic in influencing gut epithelial integrity.
Our study also highlighted the immunomodulatory functions of LA and LGG. Both LA and LGG on PBCD1 showed downregulation of various genes related to innate immune responses, such as anti-microbial peptides (REG3G, surfactant protein D), lysozyme, antigen presentation (MHC class 1 and 2) and regulation (CD32), and immune cell trafficking (CCL20/28, only LGG; ; Table S4), which may allow early colonization and adaptation of these probiotics to the host. GR signaling leads to enhanced release of pro-inflammatory cytokines IL-1β, TNF-α, and IL-6,Citation54 hence, it may be conceivable that downregulation of GR signaling at an early stage by both LA and LGG could suppress the initial host response that otherwise prevents probiotic colonization (). Once colonized on PBCD7, both probiotics induced stimulatory effects in duodenum and ileum, as indicated by higher expression of soluble (complement component 9 and 8G genes) and cellular innate mediators (non-classical MHC class I CD1d expressed on NKT cells). Furthermore, both LA and LGG induced CCL9 expression, which is a Th1-specific IFN induced chemokine similar to CXCL10 and CXCL11 genes,Citation55 and LA also increased granzyme A/B, which is expressed by cytotoxic T cells and NK cells (both associated with Th1 responses), suggesting that these probiotics may favor Th1 immune responses. Interestingly, in our previous studies we have shown Th1 immunomodulating effects of LA and LGG (in combination with Bb12) on attenuated HRV vaccine in a Gn pig model,Citation3,Citation20 suggesting that the initial Th1 microenvironment (observed on PBCD7) induced by these probiotics may result in Th1-biased adaptive immune response to specific microbial agents. Thus both LA and LGG may be useful as adjuvant for viral vaccines and infections, where Th1 immune responses are critical.
Though our study used a Gn pig model, similar observations have been made using other conventional animal models, as well as in clinical trials, supporting the usefulness of Gn pigs in elucidating mechanisms underlying the beneficial roles of probiotics. For example, comparison of transcriptome responses of germ-free piglets vs. the piglets with intestinal microbiota indicated that genes involved in biological processes like epithelial cell turnover, nutrient transport and metabolism, xenobiotic metabolism, JAK-STAT signaling pathway, and immune response were altered.Citation56 Similarly, administration of LA and LGG in a human interventional study identified mucosal transcriptional responses associated with immune response, tissue growth and development, and ion homeostasis.Citation18 In our study, genes involved in nutrient transport and metabolism, immune responses, and epithelial cell turnover are also the major classes of genes that were differentially regulated by the colonization of Gn pigs with lactobacillus species. In addition, previous studies using Gn and conventionalized mice revealed major molecular responses in metabolism, intestinal morphology and cell proliferation, and immunity as early as days 1–4 post-conventionalization, with pronounced changes occurring after day 4 post-conventionalization.Citation57,Citation58 Consistent with these findings, in our study more robust responses were observed on PBCD7, particularly in duodenum, illustrating the tissue-specific changes in the biological processes. However, detailed temporal analysis of transcriptome responses to lactobacilli colonization in neonatal Gn and conventional pigs (mimicking the infants) would provide better understanding of the biological processes modulated by these known probiotics. In addition, studies of the host proteome and metabolome should further enhance our understanding of how probiotic bacteria confer health benefits. Understanding these interactions will provide information for the rational treatment of disease phenotypes with known probiotics and will have significant implications in personalized healthcare medicine.
Materials and Methods
Bacterial strains and growth conditions
The Lactobacillus rhamnosus GG (ATCC 53103) and L. acidophilus NCFM™ (ATCC 700396) strains were routinely cultured in Lactobacilli MRS media (Hardy Diagnostics) at 37 °C with BD GasPak EZ anaerobic container system (Becton, Dickinson and Company). The freezer stocks of strains were stored at -80 °C in MRS medium with 30% glycerol (v/v). The cells from the freezer stock were plated on MRS agar, and overnight-grown culture was used to inoculate 5 ml of MRS broth and grown overnight in anaerobic conditions. The OD600nm was measured to calculate the colonization dose of 1 × 108 CFU/pig.
Ethical statement and animal experiment
All animal experiments were approved by the Institutional Animal Care and Use Committee (IACUC) of The Ohio State University. Piglets were derived surgically from near-term sows (Landrace × Yorkshire × Duroc) and maintained in sterile isolators as previously described.Citation59 Conventional culture methods (blood agar plates and thioglycolate broth culturing) were used to confirm the sterility of Gn pigs before probiotic administration. Gn pigs were divided randomly into three groups: group 1, LGG (n = 8); group 2, LA (n = 8); and group 3, non-colonized controls (n = 6). The pigs were inoculated orally with 1.0 × 108 CFU/pig at 3 d of age. Throughout the experiment, animals received the same type of feed (ultra-high temperature processed commercial cow milk [Parmalat]). The animals were euthanized on PBCD1 and 7, and tissue samples (duodenum, jejunum, ileum, cecum, colon, rectum, spleen, liver, and mesenteric lymph nodes [MLN]) were collected. In addition, rectal swabs were also collected on PBCD1, 3, and 7 to monitor colonization dynamics. Tissue samples were suspended in buffered peptone water, homogenized, suspensions were 10-fold serially diluted, and then 100 μL of each dilution was spread onto lactobacilli MRS agar plates and incubated for 24 h. The CFU/g of tissue or per ml of rectal swab was determined. The statistical analysis was performed using independent group t test or paired t test to compare colonization between strains, tissues, and days. The P value of ≤ 0.05 was considered as significant.
RNA isolation and analysis
We investigated both early (PBCD1) and late (PBCD7) transcriptome responses to probiotics in the duodenum and ileum. Since probiotic bacteria begin to colonize by day 1 and attain the maximum colonization by day 7 in these pigs, we selected these time points for analysis. A modified Trizol method was followed for RNA extraction. Briefly, the tissue samples were homogenized with Trizol reagents (Life Technologies) followed by chloroform treatment. Subsequently, the samples were centrifuged at 12,000 × g to obtain a clear lysate. The lysates were mixed immediately with an equal volume of 70% ethanol (v/v), and RNA was extracted using RNeasy mini kit (Qiagen Inc.) followed by DNase treatment. The concentration and purity of RNA samples were determined using Nanodrop ND2000c spectrophotometer (Thermo Scientific) and agarose gel electrophoresis. RNA integrity and purity was further confirmed using Agilent 2100 bioanalyzer (Agilent Technologies). The samples with ratio of 18S:28S (Sus scrofa rRNA subunits) closer to 2 were used for microarray analyses.
Transcriptome analysis
The expression analysis was performed using Sus scrofa Sureprint 4 × 44k arrays (Agilent Technologies). The Agilent Sureprint 4 × 44k high-density arrays contain in situ synthesized 60 mer oligo’s representing 44 000 pig expressed sequence tags. Microarray hybridizations were performed according to the manufacturer’s instructions at the Biomedical Genomics Core facility, The Research Institute at Nationwide Children's Hospital, Columbus, Ohio. In brief, samples were labeled with Cy3, purified using Qiagen columns, and checked for labeling efficiency using the Nanodrop. The labeled test and control samples were fragmented and hybridized to the array overnight. Microarray slides were then washed and scanned with Agilent G2505C Microarray Scanner, at 2 µM resolution. Subsequently, images were analyzed with Feature Extraction 10.10 (Agilent Technologies). Median foreground intensities were obtained for each spot. The data set was filtered to remove positive control elements and any elements that had been flagged as bad.
Statistical analysis
For the microarray intensity data, the data was background corrected and normalized with a quantile normalization method.Citation60,Citation61 A filtering method was applied to filter out low expression genes if more than 80% of arrays had expression level at or below the noise cutoff. Linear mixed effects’ models were used to account for the dependencies among observations from the same subject (each subject provided both ileum and duodenum samples). Treatment types, organ types, treatment days, and interactions among them were included as fixed effects in the model. A time factor was also included to control for the batch effects because the microarray samples were run on two different days from samples obtained from 2 independent experiments. For each comparison type I error was controlled by allowing for number of false positives among the tested genes.Citation62 Because of the small sample size, to determine the expression of a gene is significantly different between conditions, we controlled type I error at 0.0003 by controlling 10 false positives out of the tested genes (around 30 000) for each comparison (cutoff for p-value = 0.0003). Again, due to the nature of the study, for the pathway analysis, we also included some genes with more than 2-fold changes, but with p-values larger than the cutoff.
Ingenuity pathway analysis (IPA)
The web-based pathway analysis tool, IPA (www.ingenuity.com, Ingenuity Systems®) was used to identify biological functions and molecular networks modulated in duodenum and ileum by LA and LGG. The significant (± 2 ≥ or ≤ 2) regulated genes were uploaded into IPA along with the gene identifiers and corresponding fold change values. Each gene identifier was mapped to its corresponding gene object in the Ingenuity Pathways Knowledge Base (IPKB), and a set of relevant networks, focus genes, and canonical pathways were identified. The significance of the association between the genes from the data set and the canonical pathways was measured as described in IPA.
For network analysis, networks of differentially expressed genes were algorithmically generated based on their connectivity. Two genes are considered to be connected if there is a path in the network between them, and the highly interconnected networks likely represent significant biological function. The functional analysis of a network identifies the biological functions and/or diseases that were most significant to the genes in the network, and the functional analysis of the entire data set identifies the biological functions and/or diseases that are most significant to the data set. Fischer’s exact test was used to calculate a P value for each biological function assigned to that network.
Reverse-transcription quantitative PCR (RT-qPCR)
To validate the microarray data we tested the expression profiles of selected genes using RT-qPCR. The available similar sequences were searched using Tentative Consensus number (TC #) with Sus scrofa deposited reference sequence at http://compbio.dfci.harvard.edu/cgi-bin/tgi/gimain.pl?gudb=pig to design primers for RT-qPCR. Primers were designed and commercially synthesized using Integrated DNA Technologies (IDT). The primers used with their gene description are listed (Table S5). The cDNA was synthesized from total RNA using SuperScript® III First-Strand Synthesis SuperMix kit (Invitrogen). cDNA concentration was normalized, and RT-qPCR was performed using SensiMixPlus® SYBR RT-PCR Kit (Bioline) in a realplexCitation2 mastercycler (Eppendorf). The difference in expression of genes was calculated using the comparative threshold cycle (∆∆Ct) methodCitation63 to yield fold-difference in transcript levels.
| Abbreviations: | ||
| LGG | = | Lactobacillus rhamnosus |
| LA | = | Lactobacillus acidophilus |
| Gn pigs | = | gnotobiotic pigs |
| PBCD | = | post bacterial colonization day |
| CFU | = | colony forming unit |
| IPA | = | Ingenuity pathway analysis |
| IPKB | = | Ingenuity pathways knowledge base, MLN, mesenteric lymph node |
| CP | = | canonical pathway |
Additional material
Download Zip (1.1 MB)Disclosure of Potential Conflicts of Interest
No potential conflict of interest was disclosed.
Acknowledgments
We gratefully acknowledge the technical assistance of Dr Juliette Hanson, Rich McCormick, Lindsey Good, Joshua Amimo, Isaac Kashoma, and Kyle T Scheuer. This work was supported by grants from the NIH, NCCAM R21AT004716 and NIAID R01 A1099451 (L.J.S. [PI], A.N.V., and G.R. [co-PI]), and federal funds appropriated to the Ohio Agricultural Research and Development Center (OARDC) of The Ohio State University.
References
- Eastham EJ, Lichauco T, Grady MI, Walker WA. Antigenicity of infant formulas: role of immature intestine on protein permeability. J Pediatr 1978; 93:561 - 4; http://dx.doi.org/10.1016/S0022-3476(78)80888-9; PMID: 568172
- Axelsson IE, Ivarsson SA, Räihä NC. Protein intake in early infancy: effects on plasma amino acid concentrations, insulin metabolism, and growth. Pediatr Res 1989; 26:614 - 7; http://dx.doi.org/10.1203/00006450-198912000-00020; PMID: 2689986
- Chattha KS, Vlasova AN, Kandasamy S, Rajashekara G, Saif LJ. Divergent immunomodulating effects of probiotics on T cell responses to oral attenuated human rotavirus vaccine and virulent human rotavirus infection in a neonatal gnotobiotic piglet disease model. J Immunol 2013; 191:2446 - 56; http://dx.doi.org/10.4049/jimmunol.1300678; PMID: 23918983
- de Vrese M, Schrezenmeir J. Probiotics, prebiotics, and synbiotics. Adv Biochem Eng Biotechnol 2008; 111:1 - 66; http://dx.doi.org/10.1007/10_2008_097; PMID: 18461293
- Boirivant M, Strober W. The mechanism of action of probiotics. Curr Opin Gastroenterol 2007; 23:679 - 92; http://dx.doi.org/10.1097/MOG.0b013e3282f0cffc; PMID: 17906447
- Petrof EO. O Probiotics and gastrointestinal diseases: clinical evidence and basic science. Antiinflamm Antiallergy Agents Med Chem 2009; 8:260 - 9; PMID: 20890386
- Walter J. Ecological role of lactobacilli in the gastrointestinal tract: implications for fundamental and biomedical research. Appl Environ Microbiol 2008; 74:4985 - 96; http://dx.doi.org/10.1128/AEM.00753-08; PMID: 18539818
- Vanderhoof JA, Whitney DB, Antonson DL, Hanner TL, Lupo JV, Young RJ. Lactobacillus GG in the prevention of antibiotic-associated diarrhea in children. J Pediatr 1999; 135:564 - 8; http://dx.doi.org/10.1016/S0022-3476(99)70053-3; PMID: 10547243
- Guandalini S, Pensabene L, Zikri MA, Dias JA, Casali LG, Hoekstra H, Kolacek S, Massar K, Micetic-Turk D, Papadopoulou A, et al. Lactobacillus GG administered in oral rehydration solution to children with acute diarrhea: a multicenter European trial. J Pediatr Gastroenterol Nutr 2000; 30:54 - 60; http://dx.doi.org/10.1097/00005176-200001000-00018; PMID: 10630440
- Szajewska H, Kotowska M, Mrukowicz JZ, Armańska M, Mikołajczyk W. Efficacy of Lactobacillus GG in prevention of nosocomial diarrhea in infants. J Pediatr 2001; 138:361 - 5; http://dx.doi.org/10.1067/mpd.2001.111321; PMID: 11241043
- Fang SB, Lee HC, Hu JJ, Hou SY, Liu HL, Fang HW. Dose-dependent effect of Lactobacillus rhamnosus on quantitative reduction of faecal rotavirus shedding in children. J Trop Pediatr 2009; 55:297 - 301; http://dx.doi.org/10.1093/tropej/fmp001; PMID: 19203988
- Majamaa H, Isolauri E, Saxelin M, Vesikari T. Lactic acid bacteria in the treatment of acute rotavirus gastroenteritis. J Pediatr Gastroenterol Nutr 1995; 20:333 - 8; http://dx.doi.org/10.1097/00005176-199504000-00012; PMID: 7608829
- Rosenfeldt V, Michaelsen KF, Jakobsen M, Larsen CN, Møller PL, Pedersen P, Tvede M, Weyrehter H, Valerius NH, Paerregaard A. Effect of probiotic Lactobacillus strains in young children hospitalized with acute diarrhea. Pediatr Infect Dis J 2002; 21:411 - 6; http://dx.doi.org/10.1097/00006454-200205000-00012; PMID: 12150178
- Yuan-Kun Lee. Rev. ed. of: Handbook of probiotics 2nd ed. 2009; Hoboken, N.J.: John Wiley & Sons. pp. 441–443.
- Plantinga TS, van Maren WW, van Bergenhenegouwen J, Hameetman M, Nierkens S, Jacobs C, de Jong DJ, Joosten LA, van’t Land B, Garssen J, et al. Differential Toll-like receptor recognition and induction of cytokine profile by Bifidobacterium breve and Lactobacillus strains of probiotics. Clin Vaccine Immunol 2011; 18:621 - 8; http://dx.doi.org/10.1128/CVI.00498-10; PMID: 21288993
- Silva M, Jacobus NV, Deneke C, Gorbach SL. Antimicrobial substance from a human Lactobacillus strain. Antimicrob Agents Chemother 1987; 31:1231 - 3; http://dx.doi.org/10.1128/AAC.31.8.1231; PMID: 3307619
- Yan F, Polk DB. Probiotic bacterium prevents cytokine-induced apoptosis in intestinal epithelial cells. J Biol Chem 2002; 277:50959 - 65; http://dx.doi.org/10.1074/jbc.M207050200; PMID: 12393915
- van Baarlen P, Troost F, van der Meer C, Hooiveld G, Boekschoten M, Brummer RJ, Kleerebezem M. Human mucosal in vivo transcriptome responses to three lactobacilli indicate how probiotics may modulate human cellular pathways. Proc Natl Acad Sci U S A 2011; 108:Suppl 1 4562 - 9; http://dx.doi.org/10.1073/pnas.1000079107; PMID: 20823239
- Troost FJ, van Baarlen P, Lindsey P, Kodde A, de Vos WM, Kleerebezem M, Brummer RJ. Identification of the transcriptional response of human intestinal mucosa to Lactobacillus plantarum WCFS1 in vivo. BMC Genomics 2008; 9:374; http://dx.doi.org/10.1186/1471-2164-9-374; PMID: 18681965
- Zhang W, Azevedo MS, Gonzalez AM, Saif LJ, Van Nguyen T, Wen K, Yousef AE, Yuan L. Influence of probiotic Lactobacilli colonization on neonatal B cell responses in a gnotobiotic pig model of human rotavirus infection and disease. Vet Immunol Immunopathol 2008; 122:175 - 81; http://dx.doi.org/10.1016/j.vetimm.2007.10.003; PMID: 18023882
- Wen K, Li G, Zhang W, Azevedo MS, Saif LJ, Liu F, Bui T, Yousef A, Yuan L. Development of γδ T cell subset responses in gnotobiotic pigs infected with human rotaviruses and colonized with probiotic lactobacilli. Vet Immunol Immunopathol 2011; 141:267 - 75; http://dx.doi.org/10.1016/j.vetimm.2011.03.016; PMID: 21489639
- Preidis GA, Saulnier DM, Blutt SE, Mistretta TA, Riehle KP, Major AM, Venable SF, Finegold MJ, Petrosino JF, Conner ME. Versalovic.J. Probiotics stimulate enterocyte migration and microbial diversity in the neonatal mouse intestine. FASEB 2012; 26:1960 - 9; http://dx.doi.org/10.1096/fj.10-177980
- Meurens F, Summerfield A, Nauwynck H, Saif L, Gerdts V. The pig: a model for human infectious diseases. Trends Microbiol 2012; 20:50 - 7; http://dx.doi.org/10.1016/j.tim.2011.11.002; PMID: 22153753
- van Baarlen P, Troost FJ, van Hemert S, van der Meer C, de Vos WM, de Groot PJ, Hooiveld GJ, Brummer RJ, Kleerebezem M. Differential NF-kappaB pathways induction by Lactobacillus plantarum in the duodenum of healthy humans correlating with immune tolerance. Proc Natl Acad Sci U S A 2009; 106:2371 - 6; http://dx.doi.org/10.1073/pnas.0809919106; PMID: 19190178
- Kleerebezem M, Vaughan EE. Probiotic and gut lactobacilli and bifidobacteria: molecular approaches to study diversity and activity. Annu Rev Microbiol 2009; 63:269 - 90; http://dx.doi.org/10.1146/annurev.micro.091208.073341; PMID: 19575569
- Ibnou-Zekri N, Blum S, Schiffrin EJ, von der Weid T. Divergent patterns of colonization and immune response elicited from two intestinal Lactobacillus strains that display similar properties in vitro. Infect Immun 2003; 71:428 - 36; http://dx.doi.org/10.1128/IAI.71.1.428-436.2003; PMID: 12496193
- Belkaid Y, Naik S. Compartmentalized and systemic control of tissue immunity by commensals. Nat Immunol 2013; 14:646 - 53; http://dx.doi.org/10.1038/ni.2604; PMID: 23778791
- Lebeer S, Vanderleyden J, De Keersmaecker SC. Genes and molecules of lactobacilli supporting probiotic action. Microbiol Mol Biol Rev 2008; 72:728 - 64; http://dx.doi.org/10.1128/MMBR.00017-08; PMID: 19052326
- Doherty GJ, McMahon HT. Mediation, modulation, and consequences of membrane-cytoskeleton interactions. Annu Rev Biophys 2008; 37:65 - 95; http://dx.doi.org/10.1146/annurev.biophys.37.032807.125912; PMID: 18573073
- Miner JN, Hong MH, Negro-Vilar A. New and improved glucocorticoid receptor ligands. Expert Opin Investig Drugs 2005; 14:1527 - 45; http://dx.doi.org/10.1517/13543784.14.12.1527; PMID: 16307492
- Chan HM, La Thangue NB. p300/CBP proteins: HATs for transcriptional bridges and scaffolds. J Cell Sci 2001; 114:2363 - 73; PMID: 11559745
- Legate KR, Montañez E, Kudlacek O, Fässler R. ILK, PINCH and parvin: the tIPP of integrin signalling. Nat Rev Mol Cell Biol 2006; 7:20 - 31; http://dx.doi.org/10.1038/nrm1789; PMID: 16493410
- Hannigan G, Troussard AA, Dedhar S. Integrin-linked kinase: a cancer therapeutic target unique among its ILK. Nat Rev Cancer 2005; 5:51 - 63; http://dx.doi.org/10.1038/nrc1524; PMID: 15630415
- Bladh LG, Lidén J, Pazirandeh A, Rafter I, Dahlman-Wright K, Nilsson S, Okret S. Identification of target genes involved in the antiproliferative effect of glucocorticoids reveals a role for nuclear factor-(kappa)B repression. Mol Endocrinol 2005; 19:632 - 43; http://dx.doi.org/10.1210/me.2004-0294; PMID: 15528271
- Harvey CB, Williams GR. Mechanism of thyroid hormone action. Thyroid 2002; 12:441 - 6; http://dx.doi.org/10.1089/105072502760143791; PMID: 12165104
- Wan M, Leavens KF, Saleh D, Easton RM, Guertin DA, Peterson TR, Kaestner KH, Sabatini DM, Birnbaum MJ. Postprandial hepatic lipid metabolism requires signaling through Akt2 independent of the transcription factors FoxA2, FoxO1, and SREBP1c. Cell Metab 2011; 14:516 - 27; http://dx.doi.org/10.1016/j.cmet.2011.09.001; PMID: 21982711
- Chou JY, Mansfield BC. Mutations in the glucose-6-phosphatase-alpha (G6PC) gene that cause type Ia glycogen storage disease. Hum Mutat 2008; 29:921 - 30; http://dx.doi.org/10.1002/humu.20772; PMID: 18449899
- Wolvers D, Antoine JM, Myllyluoma E, Schrezenmeir J, Szajewska H, Rijkers GT. Guidance for substantiating the evidence for beneficial effects of probiotics: prevention and management of infections by probiotics. J Nutr 2010; 140:698S - 712S; http://dx.doi.org/10.3945/jn.109.113753; PMID: 20107143
- Turpin W, Humblot C, Thomas M, Guyot JP. Lactobacilli as multifaceted probiotics with poorly disclosed molecular mechanisms. Int J Food Microbiol 2010; 143:87 - 102; http://dx.doi.org/10.1016/j.ijfoodmicro.2010.07.032; PMID: 20801536
- Vendt N, Grünberg H, Tuure T, Malminiemi O, Wuolijoki E, Tillmann V, Sepp E, Korpela R. Growth during the first 6 months of life in infants using formula enriched with Lactobacillus rhamnosus GG: double-blind, randomized trial. J Hum Nutr Diet 2006; 19:51 - 8; http://dx.doi.org/10.1111/j.1365-277X.2006.00660.x; PMID: 16448475
- Saran S, Gopalan S, Krishna TP. Use of fermented foods to combat stunting and failure to thrive. Nutrition 2002; 18:393 - 6; http://dx.doi.org/10.1016/S0899-9007(01)00790-0; PMID: 11985943
- de Silva DM, Aust SD. Ferritin and ceruloplasmin in oxidative damage: review and recent findings. Can J Physiol Pharmacol 1993; 71:715 - 20; http://dx.doi.org/10.1139/y93-107; PMID: 8313236
- Ganz T. Hepcidin and iron regulation, 10 years later. Blood 2011; 117:4425 - 33; http://dx.doi.org/10.1182/blood-2011-01-258467; PMID: 21346250
- Wintergerst ES, Maggini S, Hornig DH. Contribution of selected vitamins and trace elements to immune function. Ann Nutr Metab 2007; 51:301 - 23; http://dx.doi.org/10.1159/000107673; PMID: 17726308
- Hooper LV, Midtvedt T, Gordon JI. How host-microbial interactions shape the nutrient environment of the mammalian intestine. Annu Rev Nutr 2002; 22:283 - 307; http://dx.doi.org/10.1146/annurev.nutr.22.011602.092259; PMID: 12055347
- Ataie-Jafari A, Larijani B, Alavi Majd H, Tahbaz F. Cholesterol-lowering effect of probiotic yogurt in comparison with ordinary yogurt in mildly to moderately hypercholesterolemic subjects. Ann Nutr Metab 2009; 54:22 - 7; http://dx.doi.org/10.1159/000203284; PMID: 19229114
- Wasan KM, Brocks DR, Lee SD, Sachs-Barrable K, Thornton SJ. Impact of lipoproteins on the biological activity and disposition of hydrophobic drugs: implications for drug discovery. Nat Rev Drug Discov 2008; 7:84 - 99; http://dx.doi.org/10.1038/nrd2353; PMID: 18079757
- De Preter V, Raemen H, Cloetens L, Houben E, Rutgeerts P, Verbeke K. Effect of dietary intervention with different pre- and probiotics on intestinal bacterial enzyme activities. Eur J Clin Nutr 2008; 62:225 - 31; http://dx.doi.org/10.1038/sj.ejcn.1602706; PMID: 17327863
- Gagné D, Groulx JF, Benoit YD, Basora N, Herring E, Vachon PH, Beaulieu JF. Integrin-linked kinase regulates migration and proliferation of human intestinal cells under a fibronectin-dependent mechanism. J Cell Physiol 2010; 222:387 - 400; http://dx.doi.org/10.1002/jcp.21963; PMID: 19885839
- Kim YB, Choi S, Choi MC, Oh MA, Lee SA, Cho M, Mizuno K, Kim SH, Lee JW. Cell adhesion-dependent cofilin serine 3 phosphorylation by the integrin-linked kinase.c-Src complex. J Biol Chem 2008; 283:10089 - 96; http://dx.doi.org/10.1074/jbc.M708300200; PMID: 18252715
- McDonald PC, Fielding AB, Dedhar S. Integrin-linked kinase--essential roles in physiology and cancer biology. J Cell Sci 2008; 121:3121 - 32; http://dx.doi.org/10.1242/jcs.017996; PMID: 18799788
- Markov AG, Veshnyakova A, Fromm M, Amasheh M, Amasheh S. Segmental expression of claudin proteins correlates with tight junction barrier properties in rat intestine. J Comp Physiol B 2010; 180:591 - 8; http://dx.doi.org/10.1007/s00360-009-0440-7; PMID: 20049600
- Schmitt E, Gehrmann M, Brunet M, Multhoff G, Garrido C. Intracellular and extracellular functions of heat shock proteins: repercussions in cancer therapy. J Leukoc Biol 2007; 81:15 - 27; http://dx.doi.org/10.1189/jlb.0306167; PMID: 16931602
- Busillo JM, Azzam KM, Cidlowski JA. Glucocorticoids sensitize the innate immune system through regulation of the NLRP3 inflammasome. J Biol Chem 2011; 286:38703 - 13; http://dx.doi.org/10.1074/jbc.M111.275370; PMID: 21940629
- Xanthou G, Duchesnes CE, Williams TJ, Pease JE. CCR3 functional responses are regulated by both CXCR3 and its ligands CXCL9, CXCL10 and CXCL11. Eur J Immunol 2003; 33:2241 - 50; http://dx.doi.org/10.1002/eji.200323787; PMID: 12884299
- Chowdhury SR, King DE, Willing BP, Band MR, Beever JE, Lane AB, Loor JJ, Marini JC, Rund LA, Schook LB, et al. Transcriptome profiling of the small intestinal epithelium in germfree versus conventional piglets. BMC Genomics 2007; 8:215; http://dx.doi.org/10.1186/1471-2164-8-215; PMID: 17615075
- El Aidy S, van Baarlen P, Derrien M, Lindenbergh-Kortleve DJ, Hooiveld G, Levenez F, Doré J, Dekker J, Samsom JN, Nieuwenhuis EE, et al. Temporal and spatial interplay of microbiota and intestinal mucosa drive establishment of immune homeostasis in conventionalized mice. Mucosal Immunol 2012; 5:567 - 79; http://dx.doi.org/10.1038/mi.2012.32; PMID: 22617837
- El Aidy S, Derrien M, Merrifield CA, Levenez F, Doré J, Boekschoten MV, Dekker J, Holmes E, Zoetendal EG, van Baarlen P, et al. Gut bacteria-host metabolic interplay during conventionalisation of the mouse germfree colon. ISME J 2013; 7:743 - 55; http://dx.doi.org/10.1038/ismej.2012.142; PMID: 23178667
- Meyer RC, Bohl EH, Kohler EM. E.H. Bohl And E.M. Kohler. Procurement and maintenance of germfree swine for microbiological investigations. Appl Microbiol 1964; 12:295 - 300; PMID: 14199016
- Rao Y, Lee Y, Jarjoura D, Ruppert AS, Liu CG, Hsu JC, Hagan JP. A comparison of normalization techniques for microRNA microarray data. Stat Appl Genet Mol Biol 2008; 7:e22; http://dx.doi.org/10.2202/1544-6115.1287; PMID: 18673291
- Irizarry RA, Hobbs B, Collin F, Beazer-Barclay YD, Antonellis KJ, Scherf U, Speed TP. Exploration, normalization, and summaries of high density oligonucleotide array probe level data. Biostatistics 2003; 4:249 - 64; http://dx.doi.org/10.1093/biostatistics/4.2.249; PMID: 12925520
- Gordon A., Glazko G., Qiu X., Yakovlev A.. Control of the mean number of false discoveries, Bonferroni and stability of multiple testing. Annals of applied statistics 2007; 1:179 - 190; http://dx.doi.org/10.1214/07-AOAS102
- Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001; 25:402 - 8; http://dx.doi.org/10.1006/meth.2001.1262; PMID: 11846609