Abstract
Despite the enormous efforts displayed globally in the fight against tuberculosis, the disease incidence has modified slightly, which has led to a renewed interest in immunotherapy. In general, successful immunotherapeutic candidates against tuberculosis are agents that can trigger strong, specific pro-inflammatory responses, especially of the T-helper (Th) 1 pattern. However, how these pro-inflammatory agents effectively kill the bacteria without eliciting immunopathology is not well understood. We reasoned that, in addition to the specific immune response elicited by immunotherapy, the evaluation of the overall pro-inflammatory responses should provide additional and valuable information that will be useful in avoiding immunopathology. We evaluated the overall IFN-γ and IL-17 pro-inflammatory responses among CD4+, CD8+ and γδ T cells in the lungs of mice that were infected with M. tuberculosis and treated with a DNA vaccine in an immunotherapeutic regimen. Our results demonstrate that mice that effectively combat the pathogen develop a strong, specific Th1 immune response against the therapeutic antigen and have reduced lung inflammation, present in parallel a fine-tuning in the total IFN-γ- and IL-17-mediated immunity in the lungs. This modulation of the total immune response involves reducing the Th17 cell population, augmenting CD8+ T cells that produce IFN-γ and increasing the total γδ T cell frequency. These results stress the importance of a broad evaluation of not only the specific immune response at the time to evaluate new immune interventional strategies against tuberculosis but also non-conventional T cells, such as γδ T lymphocytes.
Introduction
Infection by Mycobacterium tuberculosis results in the development of a chronic infection that can persist throughout the life of the host. The reactivation of the disease and improper treatment can cause the death of the patient due to chronic inflammation that destroys the lung parenchyma.Citation1 Both factors are not rare to observe because active tuberculosis is associated with a patient’s poor immunological condition. Additionally, the long period of treatment is difficult to complete and potentially causes the development of multi-drug resistant (MDR-TB) and extensively drug-resistant (XDR-TB) cases of tuberculosis.Citation2
The concept of boosting the immune system of the patient using immunotherapeutic regimens has recently gained new attention, while facing the frustrating reality that no new specific anti-tuberculosis drugs are available in more than 40 y.Citation3,Citation4 Several candidates for tuberculosis immunotherapy are being studied including different strains of mycobacterial species, M. tuberculosis-derived preparations and several antigens including heat-shock proteins in the form of recombinant proteins or genetic vaccines, as is the case of DNA-hsp65.Citation3 Extensive evidence demonstrated that this molecule can protect against a subsequent challenge by M. tuberculosisCitation5-Citation7 and can also treat the active disease by stimulating strong anti-Hsp65-specific IFN-γ-secreting Th1 cells, increasing the activity of cytotoxic CD8+ lymphocytes and reducing the bacterial loads.Citation8-Citation10 The reduction of lung inflammation in DNA-hsp65-treated mice has also been observed with this strong pro-inflammatory response.Citation8,Citation9,Citation11 These data represent a paradox wherein a pro-inflammatory response elicited by immunization with DNA-hsp65 can effectively kill bacteria but does not worsen the inflammation of the lungs. Moreover, anti-Hsp65-specific cells that are activated by the immunotherapy represent only a small fraction of the lymphocytes present in the lung to combat the bacillus. This fact indicates the necessity to evaluate the overall immune response that occurs in the lungs in addition to the classical evaluation of antigen-specific cells elicited by the vaccine antigen. This is especially important to better understand how to achieve the balance between the pro-inflammatory response and tissue preservation in successful immune interventional strategies against tuberculosis.Citation12 Therefore, in the present study, we evaluated the IFN-γ and IL-17 overall responses in the lungs of mice that were infected with M. tuberculosis and treated with DNA-hsp65 immunotherapy during a short follow-up (10 d) and long follow-up (60 d) after treatment completion. We observed a refinement in the IFN-γ and IL-17-mediated immunity in the lungs along with a reduction in the total Th17 cell frequency, an increase in CD8+ T lymphocytes producing IFN-γ and an increase of γδ T cell frequency in parallel with the Th1 antigen-specific immune response elicited by DNA-hsp65 therapy.
Results
Systemic reduction of bacilli loads 60 d after the end of DNA-hsp65 immunotherapy
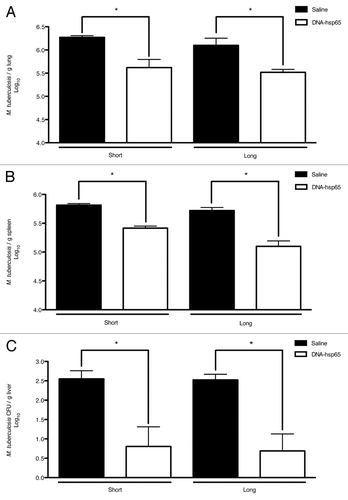
Previous observations of our group demonstrated that the DNA-hsp65 genetic vaccine reduces the bacilli loads not only in the lungs, but also in the spleen and liver of mice evaluated during a short follow-up (10 d) after therapy completion.Citation11 In this context, we first wanted to determine if this reduction was maintained throughout the long follow-up (60 d). To this aim, the mice started on the DNA-hsp65 immunotherapy 30 d after infection with M. tuberculosis. The animals received a total of four doses of DNA-hsp65 at 10- days intervals, and the determination of the colony-forming units were assessed during short and long follow-up after the end of the therapy. illustrates that the therapeutic effects of DNA-hsp65 were maintained at least 60 d after the last dose, not only in lungs but also in the spleen and liver. These results indicated that the systemic effects of DNA-hsp65 against the bacillus were maintained for a long period after therapy completion.
Figure 1. Therapeutic efficacy of DNA-hsp65 immunization against tuberculosis. M. tuberculosis challenge was performed on day 0. The immunotherapy was initiated 30 d later. The mice received four doses of 100 μg/dose of DNA-hsp65 (ten days interval). At day 70 and 120 after challenge, corresponding to 10 d (short period) and 60 d (long period) after the end of the therapy, the number of colony forming units were determined in (A) the lungs, (B) spleen and (C) liver. The significant differences are indicated and correspond to p < 0.05 compared with the saline group by Student’s t-test. The data are presented as the means ± SEM of 7 mice per group of a representative experiment.
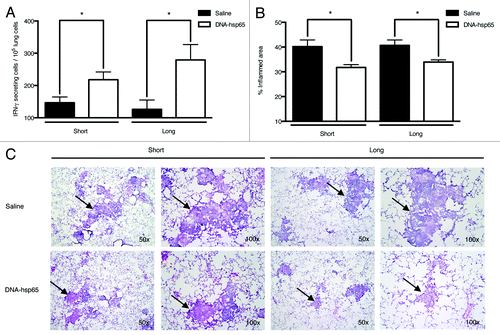
The reduction of bacilli burden is accompanied by a reduction in lung inflammation and an increased frequency of Hsp65-specific Th1 cells
An important aspect of immune interventional strategies against tuberculosis is that the agent must be able to elicit a Th1 immune response, which is essential for reducing bacilli loads, without promoting excessive inflammation and thus avoiding extensive and irreversible lung damage. For this reason, we evaluated the number of anti-Hsp65-specific T lymphocytes that secrete IFN-γ, a key effector cytokine for combating M. tuberculosis. As previously described in prophylactic studies on M. tuberculosisCitation7 as well as after short-term tuberculosis therapy,Citation8 we observed an increase in the number of anti-Hsp65-specific T cells that secrete IFN-γ in the lungs of DNA-hsp65-treated mice during short and long follow-up after therapy completion (). These results were accompanied by reduction of the pneumonic process where less leucocyte infiltration, more focalization of the granulomatous process and larger areas of lung tissue preservation were observed compared with untreated animals ( and C, respectively), demonstrating the ability of DNA-hsp65 to trigger a specific Th1 response to effectively combat the infection.
Figure 2. Reduction of lung inflammation and induction of the Th1-specific response in mice treated with DNA-hsp65. (A) The number of T cell clones secreting IFN-γ after Hsp65 stimulation in cells from the lungs of mice untreated or treated with DNA-hsp65. (B) The measurement of the inflamed area in the lungs from treated and untreated mice. (C) The histological evaluation of lung samples stained with hematoxylin-eosin. The photographs obtained at 50x and 100x magnifications demonstrate the differences in lung inflammation (arrows) observed among the experimental groups. *p < 0.05 by Student’s t-test. The data are presented as the means ± SEM of 7 mice per group of a representative experiment.
The therapeutic efficacy of DNA-hsp65 is associated with a differential modulation of the IL-17+ lymphocytes
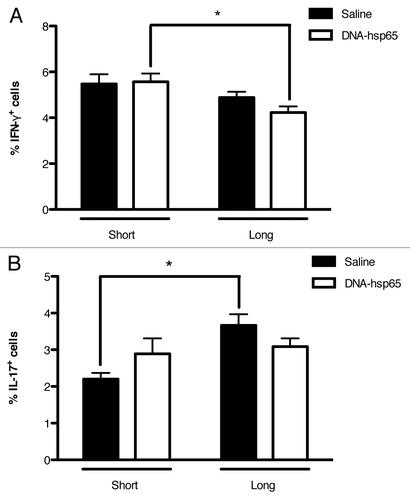
As in the case of other antigen-specific cells in various promising vaccine candidates, Hsp65-specific T cells that secrete IFN-γ appear to be instrumental in eliciting and maintaining effective anti-tuberculosis activity. However, these cells represent only a small fraction of the total population of lymphocytes present in the lungs during the infection. Therefore, in attempt to better characterize how lung damage is reduced in DNA-hsp65 animals, we evaluated the total population of T cells that produce the pro-inflammatory cytokines IFN-γ and IL-17 during short and long follow-up after therapy completion.
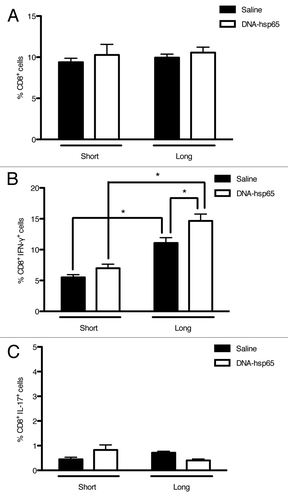
Interestingly, we observed that there was no difference in the total frequency of IFN-γ+ cells between treated and untreated animals during both periods evaluated () and that DNA-hsp65-treated animals presented a reduced frequency of IFN-γ+ cells during long follow-up (). In contrast, untreated animals that did not control the infection presented the lowest frequency of IL-17+ cells during short follow-up, exhibiting twice the frequency upon long follow-up. On the other hand, DNA-hsp65-treated mice demonstrated no change in the frequency of IL-17+ at both evaluated time points (). These results suggest that, in our model, the increasing IL-17-mediated response observed in untreated mice is associated to the chronic and unremitting process of lung inflammation.
Figure 3. Percentage of IFN-γ - and IL-17-producing cells in mice treated with DNA-hsp65. Lung cell suspensions were stained for intracellular detection of (A) IFN-γ or (B) IL-17. *p < 0.05 by two-way ANOVA with Bonferroni post-test. The data are presented as the means ± SEM of 7 mice per group of a representative experiment.
The Hsp65-specific immune response is accompanied by a fine-tuning of total Th17, CD8+ and γδ+ lymphocytes
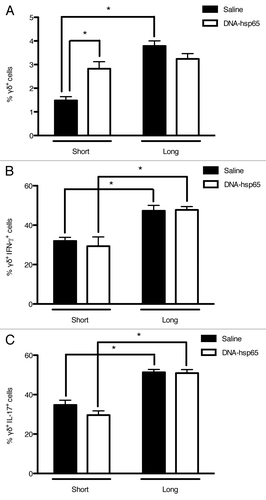
Considering that we observed differential modulation of the pro-inflammatory IFN-γ and IL-17-secreting cell frequency in the lungs of DNA-hsp65-treated compared with untreated animals, we next analyzed the main T lymphocyte populations that produce both cytokines.
First, we observed that, contrary to untreated mice, there was an increased frequency of γδ T cells immediately after the end of the immunotherapy, and these levels were maintained for at least 60 d (). In contrast, there was no difference between treated and untreated animals in the frequency of γδ T cells that secreted IFN-γ () or IL-17 (); however, the frequency of cells that secreted these cytokines increased in both groups upon the long follow-up compared with the short follow-up. These results indicate that γδ T cells play an active role throughout the infection and that one of the associated effects of DNA-hsp65 therapy is the modulation of γδ T cells, increasing their frequency in the lungs of treated animals faster than in untreated animals.
Figure 4. Effects of DNA-hsp65 immunization on γδT cells. Percentages of (A) total γδ+, (B) γδ+IFN-γ+ or (C) γδ+IL-17+ cells from the lungs. *p < 0.05 by two-way ANOVA with Bonferroni post-test. The data are presented as the means ± SEM of 7 mice per group of a representative experiment.
In contrast to that observed in γδ T cells, there was no difference in the CD8+ cell frequency during both periods between treated and untreated animals (). Nonetheless, CD8+ lymphocytes that secreted IFN-γ were more frequent in the treated group from the short to the long follow-up, and this frequency was higher than in untreated animals (). As expected, less than 1% of CD8+ cells were IL-17+ with no difference between the groups. Therefore, these results indicate that another associated effect of DNA-hsp65 treatment is the increased number of IFN-γ-secreting cytotoxic T cells.
Figure 5. Effects of DNA-hsp65 immunization on CD8+ cells. Percentages of (A) total CD8+, (B) CD8+IFN-γ+ or (C) CD8+IL-17+ cells from the lungs. *p < 0.05 by two-way ANOVA with Bonferroni post-test. The data are presented as the means ± SEM of 7 mice per group of a representative experiment.
When the T helper cell population was evaluated, we observed an increased frequency of CD4+ lymphocytes in DNA-hsp65-treated animals upon short follow-up, with a reduction to levels similar to that of untreated mice upon long follow-up (). In addition, the classical Th1 cells presented a significant augment from the short to the long follow-up but at equal percentages between treated and untreated mice (). Finally, in contrast to Th1 cells, the total population of Th17 lymphocytes was differentially modulated in treated and untreated mice. These IL17+ cells augmented their frequency almost 3-fold in untreated mice during the long follow-up. In contrast, DNA-hsp65-treated mice presented a more discrete increase in the frequency of IL17+ cells at 60 d compared with 10 d post-treatment ().
Figure 6. Effects of DNA-hsp65 immunization on CD4+ cells. Percentages of (A) total CD4+, (B) CD4+IFN-γ+ or (C) CD4+IL-17+ cells from the lungs. *p < 0.05 by two-way ANOVA with Bonferroni post-test. The data are presented as the means ± SEM of 7 mice per group of a representative experiment.
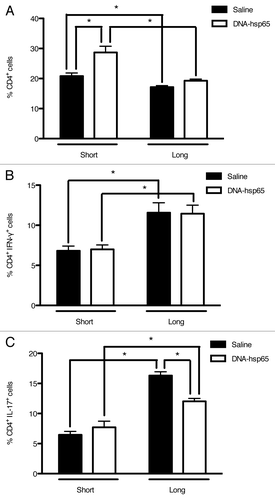
Taken together, these results indicate that, apart from the Th1 Hsp65-specific T cell priming by DNA-hsp65 during tuberculosis therapy, a fine-tuning of the total IFN-γ- and IL-17-producing cells occurred in the lungs of treated mice. In general, the therapeutic efficacy is associated with an increased frequency of total γδ+ and CD4+ T cells during the short period post-treatment; however, during the long period, the increase in cytotoxic T lymphocytes and the reduction in the Th17 cell response seem to account for the continuation of the bacilli growth restriction in DNA-hsp65-treated animals.
Discussion
One of the most ambitious challenges in the fight against tuberculosis is the development of a new effective vaccine. Innumerous strategies and candidates for the new prophylactic vaccine against tuberculosis have been developed and tested in different animal models and there are at least 11 currently being evaluated in clinical trials.Citation13,Citation14 On the other hand, considering that one third of the world’s population is infected with M. tuberculosis, the immunocompromised status of tuberculosis patients, the spread of MDR and XDR strain cases and the lack of new drugs against tuberculosis have prompted experts postulate that the effective combat of the pathogen will require immunotherapy rather than vaccination in the classical sense.Citation15,Citation16 However, immunotherapy is probably even a bigger challenge than vaccination in tuberculosis, because it is well known that the chronic nature of the infection in humans could result in a great variety of different outcomes that are not reproduced perfectly by any animal model due to the combination of different factors involving the bacteria, the host and the environment.Citation17 Furthermore, studies evaluating new prophylactic vaccines determine the “protective efficacy” of the new candidate by comparing the results obtained to BCG, which is considered a gold standard; and commonly this protection is reflected in the reduction of 0.5 to 3 log10 of bacilli loads, as extensively reviewed by Ly and McMurray.Citation18
Nonetheless, BCG has no effect as a therapeutic agent, and therefore there is no gold standard for immunotherapy screening. This multifaceted problem is reflected in the fewer immunotherapeutic molecules tested in comparison to the classical vaccine candidates against tuberculosis. In fact two formulations, M. vaccae and RUTI are considered as the main candidates for tuberculosis immunotherapy.Citation19 These formulations are based in inactivated and fragmented mycobacteria, respectively, representing a complex antigenic mixture and both have been tested regarding safety and immunogenicity in clinical trials.Citation20,Citation21
However, considerable variability was observed with M. vaccae,Citation22,Citation23 and RUTI was able to reduce bacilli loads (around 0.5 log) in the lung, but not in the spleen of guinea pigs.Citation24 As mentioned before, these limitations are due to the inherent complexity of tuberculosis disease. Because even in the case of well-controlled animal studies with formulations based on single or few antigens, considerable variation is observed when used as immunotherapies.Citation9,Citation10,Citation25-Citation29
For these reasons the determination of therapeutic efficacy is based not only in bacilli load reduction, but also in the ability to avoid lung damage, to improve the results obtained with conventional chemotherapy and increase survival, as observed with M. vaccae, RUTI and other molecules, including DNA-hsp65.Citation12,Citation16,Citation30,Citation31
In this context we have been using DNA-hsp65 as a tool to learn about immuno-interventional strategies against active tuberculosis. We and others have extensively showed that immunotherapy with DNA-hsp65 results in strong increase of the IFN-γ and reduction of IL-4 production from anti-Hsp65 specific T cells.Citation8,Citation25
These studies agree with classical evaluations of prophylactic and/or immunotherapeutic candidates, in which determination of potential clinical application is based on their ability to trigger a certain type of specific immune response, which is the Th1 pattern in tuberculosis. However, in parallel to the specific response, a large number of other cells not specific to the vaccine antigen are present and participating in the immune response against the infectious agent in the target organ. Therefore, understanding how this major fraction of cells is modulated during an immune intervention protocol could provide insight into the protective results and perhaps lead to new alternatives for improvement. In this light, we describe here what occurs within the overall cell populations that produce IFN-γ and IL-17 cytokines in a model of tuberculosis immunotherapy.
Considering that our model employs a high quantity of M. tuberculosis cells delivered by an intra-tracheal injection in contrast to the low-dose model where the bacilli are delivered by the aerosol route, and therefore a more physiological than the intra-tracheal injection, we first wanted to determine how similar were the data obtained here in the infected untreated mice in comparison to previously published data. Interestingly, we observed that in general the modulation of the T cell populations evaluated in the present study are compatible with previous reports. In this regard, the laboratory of Dr JL Flynn was the first to describe the γδ T lymphocytes as the main source of IL-17 during the immune response to M. tuberculosis.Citation32 The authors isolated cells from the lungs and performed stimulation with phorbol myristate acetate (PMA) and ionomycin previously to detect the intracellular production of IL-17 on those cells. They showed that 35% of γδ T cells are able to produce IL-17 at 56 d after infection. The results presented here at 60 d post-infection are similar to that obtained by Lockhart et al.,Citation32 and in addition to this, we also observed an increment in the γδ T cells secreting IL-17 up to 50% at 120 d post-infection. In our knowledge, this is the first description of the γδ lymphocytes producing IL-17 in the high dose infection model of tuberculosis.
In 2005, Bafica et al.,Citation33 showed the importance of toll-like receptor (TLR)-9 and TLR-2 during the immune response to tuberculosis. As part of their observations, they evaluated the production of IFN-γ by CD8+ T cells after isolation from the lungs and stimulation for a short period with anti-CD3 antibodies. The percentage of CD8+IFN-γ+ T cells detected was 6% among the lymphocyte population at 30 d after challenge. This is the same percentage detected in the present report at 60 d post-infection and interestingly, we observed a duplication of this percentage in the lungs of mice with 120 d of infection.
The participation of CD4+ T cells of the Th1 immune pattern has been broadly studied during tuberculosis as mentioned before, and is widely accepted that this response is capable to start the inhibition of the bacilli growth after 3 weeks of infection. According to Jung et al.,Citation34 using the aerosol model of tuberculosis in C57BL/6 mice, around 10.5% of CD4+ T cells secrete IFN-γ at day 30 post-infection and this number is maintained until day 85. In our case, the CD4+IFN-γ+ cells represented 7% of the CD4+ lymphocytes and this percentage increased up to 12% at 120 d post-infection.
On the other hand, the CD4+IL-17+ cells were reported to represent as few as 1% of the CD4+ T cells in the lungs of M. tuberculosis infected mice,Citation32 while we observed around 5% at 60 d and more than 15% at 120 d post-infection.
Taken together, these data indicate that the high dose model of tuberculosis triggers an immune response with various elements similar to those observed in the low dose model that employs the aerosol route of infection, with the difference on the size of the Th17 cell response. In addition, it was already demonstrated that the high dose model of tuberculosis results in a chronic and progressive disease and in contrast the low dose model induces latent non-progressive tuberculosis, nonetheless both forms are observed in human patients.Citation35,Citation36
In the particular case of tuberculosis treatment, one of the main objectives is to avoid the irreversible lung damage observed during the chronic phase of the disease, an episode that could occur even in the case of successful chemotherapy.Citation3,Citation12 In addition, as mentioned before, it is well known that an effective immune response to tuberculosis involves generating specific T cells that produce high levels of IFN-γ, a main pro-inflammatory mediator that activates phagocytes for eliminating the bacteria through reactive mediators, such as nitric oxide (NO).Citation37-Citation39 However, the process of achieving a balance between eliciting a strong pro-inflammatory response and preserving the lung parenchyma is not well understood.Citation39 In this sense, strategies for shortening the long drug treatment while reducing the lung parenchyma remodeling are of special interest.Citation16 This combination of beneficial effects may be achieved via utilizing aspects of both immunotherapy and conventional chemotherapy in treating tuberculosis. For these reasons, we also evaluated the overall pro-inflammatory response after DNA-hsp65 immunotherapy for tuberculosis. As described in previous reports from us and other research groups, immunizing with mycobacterial Hsp65 in the form of a genetic vaccine elicits a strong Th1-specific T cell response that is fundamental to combating the tuberculosis bacteria.Citation8-Citation10 We extended these results to a long follow-up of 60 d after therapy completion, representing 120 d after M. tuberculosis challenge. The treatment was able to reduce the bacilli counts not only in the lungs but also in the spleen and liver, demonstrating the systemic and sustained effects of DNA-hsp65 against tuberculosis. In parallel to the decrease in bacilli loads, the reduction of the inflamed area in the lungs was also observed in treated mice during the long follow-up after treatment completion.
Contrary to our expectations, we did not observe a difference in the size of the total IFN-γ-secreting lymphocyte populations between treated and untreated animals at short and long follow-up. We also observed a reduction in the number of IFN-γ secreting cells in treated mice upon short follow-up compared with the long follow-up evaluation. However, we did observe an increase in the frequency of the IFN-γ producers in the cytotoxic fraction of T cells. This observation is in accordance with the well-known importance of this type of lymphocytes in the immune response against intracellular pathogens, such as M. tuberculosis.Citation39 This result also highlights the ability of DNA-hsp65 to induce the immune response in a Th1 pattern, with the subsequent activation of CD8+ cells that eliminate infected cells.Citation8
The results also indicate that, in our model, is important to restrict the Th17 immune response given that a lower frequency of these lymphocytes was observed upon long follow-up in DNA-hsp65-treated mice compared with untreated animals. Although IL-17 is necessary in the response against M. tuberculosis infection, it was observed that the Th17 response is important at earlier phases of the disease. However, it is accepted that an extensive Th17 response is associated with disease progression during chronic tuberculosisCitation40 and that IFN-γ is important for limiting IL-17 production to avoid mycobacterial immune-mediated pathology.Citation41-Citation43
Caution should be taken to avoid misinterpretation of the results presented in this study regarding the frequencies of cytokine secreting cells in the DNA-hsp65 immunized animals, once a strong Th1 immune response is expected from a promising candidate against tuberculosis according to classical vaccine evaluation studies. As mentioned earlier, this strong Th1 immune response is also observed after DNA-hsp65 immunization in relation to the specific anti-Hsp65 response, as is the case of several other immunogenic antigens including Ag85, TB10.4 and ESAT-6 that elicit strong specific Th1 responses (e.g., Dietrich et al.,Citation44 Hervas-Stubbs et al.Citation45 Romano et al.Citation46). However, here we presented the overall evaluation of lung isolated T cells not only specific to Hsp65. In fact, these results are also different from several other studies where cytokine levels were determined in lung homogenates from mice vaccinated and/or treated with different vaccine candidates (including Hsp65). In these studies the amounts of cytokines detected corresponds to the production of the whole resident lung cells and not only to T cells (e.g., Bonato et al.,Citation8 Sable et al.Citation47 and Cervantes-VillagranaCitation48).
For these reasons, the data presented here is not possible to be directly compared with those from the studies recently mentioned.
One of the limitations of our results is the absence of evaluation of regulatory T cells (Tregs). It has been reported that circulating Tregs are increased in frequency in patients with tuberculosis,Citation49,Citation50 and that when patients receive anti-M. tuberculosis chemotherapy the production of IFN-γ increases with a parallel reduction of Tregs.Citation50,Citation51 In this same direction, our group has recently showed that mice vaccinated with BCG and DNA-hsp65 in an heterologous immunization procedure followed by challenge with M. tuberculosis present an increase in lung parenchyma preservation which correlates with higher CD4+/CD4+Foxp3+ T cell ratio (and also lower CFU counts in the lungs) than BCG or DNA-hsp65 single vaccinated animals.Citation52 These results suggest that one of the possible alternatives for improvement the anti-M. tuberculosis effects of DNA-hsp65, either in the vaccine or therapeutic protocols, could be by targeting Treg cells, although this also might result in reduction of the lung parenchyma preservation. In this sense, the role of Tregs should be addressed in further studies.
The lack of determination of the specificities of the evaluated cell populations, other than the anti-Hsp65 specific response (showed in ), also limits the interpretation of our results. Nevertheless, it was extensively showed that the immune response to M. tuberculosis infection results in the induction of T cell clones specific to a great number and variety of mycobacterial proteins including ESAT-6, CFP-10, TB10.4, Ag85A and B, and Hsp65.Citation53-Citation56 The analysis of the antigenic specificities in our system could be important to contribute in the identification of biomarkers of different phases of the disease, vaccination and treatment; that is one of the main goals in tuberculosis research.Citation57
Finally, an intriguing observation was obtained during the evaluation of γδ T cells. These cells presented the highest frequency of cytokine production among the three populations studied, with no difference between treated and untreated animals. Nonetheless, because their frequency upon short follow-up was clearly higher in DNA-hsp65-treated mice, it is possible that these cells present a different phenotype that was not evaluated here and that could be important in overcoming the infection. In fact, several recent studies reported a remarkable plasticity of γδ T cell population with characteristics ranging from classical adaptive T cell receptor-bearing cells to antigen-presenting cells with innate and adaptive properties. This plasticity demands further evaluation in the context of tuberculosis and also in the development of new vaccines and immunotherapeutic protocols.Citation58-Citation60
In conclusion, these results stress the importance not only of evaluating the specific response elicited by a candidate antigen at the time of vaccine and immunotherapeutics development but also that other non-conventional T cells as the γδ+ lymphocytes.
Materials and Methods
Plasmid DNA production and quality control
DNA-hsp65 was prepared as described previously.Citation5 Briefly, the hsp65 gene from M. leprae was cloned into BamH I-Not I restriction sites of a pVAX1 vector (Invitrogen). The pVAX1-hsp65 construct (DNA-hsp65) was prepared using an Endo-Free Plasmid Giga kit (Qiagen) from competent Escherichia coli growth in LB medium. The kit purification protocol is based on a modified lysis procedure, followed by binding of plasmid DNA to an anion-exchange resin under appropriate low-salt and pH conditions. Plasmid DNA is eluted in a high salt buffer and then concentrated and desalted by isopropanol precipitation. The obtained material was checked by restriction analysis and sequencing. Plasmid concentration was determined by absorbance at 260 nm and purity by calculating the ratio A260 nm/A280 nm (NanoDrop -1000). Endotoxin-free condition for DNA vaccination was determined by a Limulus Amebocyte Lysate (LAL) test as recommended by European and US Pharmacopeias,Citation61 using the QCL 1000-LAL test kit (Cambrex Bio Science).
Infection, immunotherapy and CFU determination
Female, Specific Pathogen-Free, BALB/c mice (8 weeks old) from the local animal facility were anaesthetized with 10% ketamine chloridrate (100 mg/Kg) and 2% xylazine chloridrate (20 mg/Kg) (Agener União) and infected on day 0 with 1.0 × 105 bacilli of the M. tuberculosis H37Rv strain via the intra-tracheal route, as previously described.Citation7,Citation62
Thirty days after infection, the mice began treatment with 100 μg of DNA-hsp65, in a final volume of 100 μl (25% sucrose), receiving 50 μl in each quadriceps at 10-d intervals until day 60 (total of four doses). The mice were euthanized by cervical dislocation at either day 70 or 120 after infection, corresponding to 10 and 60 d after completing immunotherapy, respectively. All procedures were performed in a level III bio-safety room and approved by the local ethical committee (COBEA, CETEA, process number 155/2006).
Colony-forming unit (CFU) determinations were performed in samples from the liver, spleen and lungs. The organs were weighed and placed in RPMI-1640 medium (Sigma-Aldrich) and homogenized. The lung samples were incubated under agitation at 37°C for 30 min in digestion solution containing 5 μg/ml of Liberase Blendzyme 2 (Roche Diagnostics, Indianapolis, USA). Petri dishes of 7H11 Milddlebrook’s medium were used to incubate serial dilutions of the organs. The colonies were counted 28 d after incubation at 37°C.
Flow cytometry
For flow cytometric analysis, the entire right lung was removed and processed as described for CFU determination. After incubation in digestion solution, the organs were filtered, and the cell suspensions were treated with 1 ml of erythrocyte lysis solution (0.15 M NH4Cl, 10 mM KHCO3 and 0.1 mM EDTA) for 1 min. The cells were washed, counted and checked for cell viability with Tripan blue 0.5% staining. The cells were plated at 1.0 × 106 cells per well and incubated in complete RPMI-1640 medium in the presence of 20 μg/ml concanavalin A (Con-A) (Sigma-Aldrich) for 6 h. After incubation, the cells were washed and incubated at 4°C for 30 min with FcBlock (supernatant of 2.4G2 cell culture, ATCC-HB-197). Cells were stained with surface markers and intracellular cytokines using the Cytofix/Cytoperm Plus Fixation/Permeabilization kit with BD Golgi Plug (BD Biosciences) following manufacter’s instructions. The following fluorochrome-labeled mAb were used: anti-TCRγδ-FITC (clone GL3), anti-CD4-PerCP (clone RM4–5), anti-CD8-APC (clone 53-6.7), anti-IFN-γ-PE (clone XMG1.2) anti-IL-17-PE (clone TC11–18H10). The isotype controls were: IgG1κ-PE (clone R3-34), IgG2κ-FITC (clone B81–3), IgG2a,κ-PerCP and IgG2a,κ-APC (clone R35-95). The cells were fixed in 2% paraformaldehyde overnight before removal from the level III bio-safety room. Samples were run in a FACSCantoTM I flow cytometer (BD Biosciences) using the FACSDiva 6.1.3 software for data acquisition. A minimum of 100,000 events was acquired per sample and analysis was performed on live, single cell lymphocytes using FlowJo software (Tree Star Inc.).
ELISPOT
ELISPOT assay was conducted using Mouse IFN-γ Set (BD Biosciences) according to the manufacturer's instructions. Briefly, 100 μl of anti-IFN-γ capture antibody solution (5 µg/ml) were distributed to each well of the ELISPOT plate, following overnight incubation at 4°C. Coating was discarded and blocking solution was added to avoid unspecific results. Once plates were prepared, 1.0 × 105 lung cells suspensions were incubated with 10 μg/ml of recombinant Hsp65 in complete RPMI medium at 37°C, 5% CO2 and 99% humidity for 48 h. Con-A was used as control. Cell suspensions were aspirated and wells were washed and filled with100 μl of biotinylated detection antibody solution (2 µg/ml) for 2 h at room temperature. The wells were washed and filled with 100 μl/well Streptavidin-Horseradish Peroxidase (1:100) for 1 h at room temperature. Finally, the wells were washed and filled with 100 μl of substrate solution (3-Amino-9-ethylcarbazole, AEC). Spot development was monitored and the reaction was stopped by washing wells with deionized water. Plates were air-dried overnight at room temperature in the dark. Spots were detected using Bioreader-3000 (Bio-Sys, GmbH) and ImmunoSpot software (version 3.0). Values for each animal were obtained by average of triplicate wells. Results on graph represent the mean number of IFN-γ- secreting cells (spots) for 7 mice per group.
Histological and morphometric analysis of the lungs
The morphometric analysis was performed following recommendations as described before.Citation7,Citation63 Briefly, left lungs from mice at days 10 and 60 after infection, were inflated and used to obtain paraffin sections that were stained with hematoxylin-eosin and evaluated by a pathologist in a single-blinded manner. The morphometric analysis was performed using a conventional light microscope (Axioplan) with an integrated eyepiece and a system made of a 100-point grid consisting of 10 × 10 lines of known length. The volume fraction of normal and affected tissue (that included granuloma and non-granulomatous penumonitis) areas was measured at 200 × magnification using the point-counting technique in 10 random, non-coincident microscopic fields. The points that hit the tissue areas were counted and divided by the total number of points in each microscopic field. The data were reported as the fractional area of pulmonary tissue presented pneumonic score.
Statistical analysis
The GraphPad Prism software, version 5.0 (GraphPad Software) was used to perform the statistical analysis. The Student’s t-test was applied for two-group comparison or two-way ANOVA with Bonferroni post-test for multiple comparisons. The values of p < 0.05 were considered to be statistically significant. All experiments were repeated at least twice. The data are presented as the means ± SEM of 7 mice per group of a representative experiment.
| Abbreviations: | ||
| Hsp65 | = | heat-shock protein of 65 kDa |
| IFN-γ | = | Interferon-gamma |
| IL-17 | = | Interleukin 17 |
| WHO | = | World Health Organization. CFU, colony-forming units |
Acknowledgments
This study was supported by the Fundação de Amparo à Pesquisa do Estado de São Paulo, FAPESP (Grant number 2006/05963–8). We are thankful to Dr Patricia Dillenburg-Pilla and the National Institutes of Health Fellows Editorial Board (NIH-FEB, Bethesda, USA) for a critical review of the manuscript.
Note
At the time this article was in press, the Food and Drug Administration (FDA) of the United States of America approved bedaquiline, the first new anti-tuberculosis drug in more than 40 y. Bedaquiline is approved only for MDR-TB cases and should be used in combination with other drugs.Citation64
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Related Research Data
References
- Svenson S, Källenius G, Pawlowski A, Hamasur B. Towards new tuberculosis vaccines. Hum Vaccin 2010; 6:309 - 17; http://dx.doi.org/10.4161/hv.6.4.10711; PMID: 20372087
- Global Tuberculosis Control WHO. WHO Report 2010. WHO 2010.
- Churchyard GJ, Kaplan G, Fallows D, Wallis RS, Onyebujoh P, Rook GA. Advances in immunotherapy for tuberculosis treatment. [ix.] Clin Chest Med 2009; 30:769 - 82, ix; http://dx.doi.org/10.1016/j.ccm.2009.08.009; PMID: 19925966
- Ma Z, Lienhardt C, McIlleron H, Nunn AJ, Wang X. Global tuberculosis drug development pipeline: the need and the reality. Lancet 2010; 375:2100 - 9; http://dx.doi.org/10.1016/S0140-6736(10)60359-9; PMID: 20488518
- de Paula L, Silva CL, Carlos D, Matias-Peres C, Sorgi CA, Soares EG, et al. Comparison of different delivery systems of DNA vaccination for the induction of protection against tuberculosis in mice and guinea pigs. Genet Vaccines Ther 2007; 5:2; http://dx.doi.org/10.1186/1479-0556-5-2; PMID: 17250766
- dos Santos SA, Zárate-Bladés CR, de Sá Galetti FC, Brandão IT, Masson AP, Soares EG, et al. A subunit vaccine based on biodegradable microspheres carrying rHsp65 protein and KLK protects BALB/c mice against tuberculosis infection. Hum Vaccin 2010; 6:1047 - 53; http://dx.doi.org/10.4161/hv.6.12.13350; PMID: 21157178
- Souza PR, Zárate-Bladés CR, Hori JI, Ramos SG, Lima DS, Schneider T, et al. Protective efficacy of different strategies employing Mycobacterium leprae heat-shock protein 65 against tuberculosis. Expert Opin Biol Ther 2008; 8:1255 - 64; http://dx.doi.org/10.1517/14712598.8.9.1255; PMID: 18694348
- Bonato VL, Gonçalves ED, Soares EG, Santos Júnior RR, Sartori A, Coelho-Castelo AA, et al. Immune regulatory effect of pHSP65 DNA therapy in pulmonary tuberculosis: activation of CD8+ cells, interferon-gamma recovery and reduction of lung injury. Immunology 2004; 113:130 - 8; http://dx.doi.org/10.1111/j.1365-2567.2004.01931.x; PMID: 15312144
- Nuermberger E, Tyagi S, Williams KN, Rosenthal I, Bishai WR, Grosset JH. Rifapentine, moxifloxacin, or DNA vaccine improves treatment of latent tuberculosis in a mouse model. Am J Respir Crit Care Med 2005; 172:1452 - 6; http://dx.doi.org/10.1164/rccm.200507-1047OC; PMID: 16151038
- Okada M, Kita Y, Nakajima T, Kanamaru N, Hashimoto S, Nagasawa T, et al. Novel therapeutic vaccine: granulysin and new DNA vaccine against Tuberculosis. Hum Vaccin 2011; 7:Suppl 60 - 7; http://dx.doi.org/10.4161/hv.7.0.14563; PMID: 21546794
- Zárate-Bladés CR, Bonato VL, da Silveira EL, Oliveira e Paula M, Junta CM, Sandrin-Garcia P, et al. Comprehensive gene expression profiling in lungs of mice infected with Mycobacterium tuberculosis following DNAhsp65 immunotherapy. J Gene Med 2009; 11:66 - 78; http://dx.doi.org/10.1002/jgm.1269; PMID: 19035575
- Rook GA, Lowrie DB, Hernàndez-Pando R. Immunotherapeutics for tuberculosis in experimental animals: is there a common pathway activated by effective protocols?. J Infect Dis 2007; 196:191 - 8; http://dx.doi.org/10.1086/518937; PMID: 17570105
- Ernst JD. The immunological life cycle of tuberculosis. Nat Rev Immunol 2012; 12:581 - 91; http://dx.doi.org/10.1038/nri3259; PMID: 22790178
- Kaufmann SH. Tuberculosis vaccine development: strength lies in tenacity. Trends Immunol 2012; 33:373 - 9; http://dx.doi.org/10.1016/j.it.2012.03.004; PMID: 22560865
- Grange JM, Brunet LR, Rieder HL. Immune protection against tuberculosis--when is immunotherapy preferable to vaccination?. Tuberculosis (Edinb) 2011; 91:179 - 85; http://dx.doi.org/10.1016/j.tube.2010.12.004; PMID: 21233019
- Doherty TM. Immunotherapy for TB. Immunotherapy 2012; 4:629 - 47; http://dx.doi.org/10.2217/imt.12.52; PMID: 22788130
- Gupta UD, Katoch VM. Animal models of tuberculosis. Tuberculosis (Edinb) 2005; 85:277 - 93; http://dx.doi.org/10.1016/j.tube.2005.08.008; PMID: 16249122
- Ly LH, McMurray DN. Tuberculosis: vaccines in the pipeline. Expert Rev Vaccines 2008; 7:635 - 50; http://dx.doi.org/10.1586/14760584.7.5.635; PMID: 18564018
- Prabowo SA, Gröschel MI, Schmidt ED, Skrahina A, Mihaescu T, Hastürk S, et al. Targeting multidrug-resistant tuberculosis (MDR-TB) by therapeutic vaccines. Med Microbiol Immunol 2012; In Press http://dx.doi.org/10.1007/s00430-012-0278-6; PMID: 23143437
- Stanford J, Stanford C, Grange J. Immunotherapy with Mycobacterium vaccae in the treatment of tuberculosis. Front Biosci 2004; 9:1701 - 19; http://dx.doi.org/10.2741/1292; PMID: 14977580
- Vilaplana C, Montané E, Pinto S, Barriocanal AM, Domenech G, Torres F, et al. Double-blind, randomized, placebo-controlled Phase I Clinical Trial of the therapeutical antituberculous vaccine RUTI. Vaccine 2010; 28:1106 - 16; http://dx.doi.org/10.1016/j.vaccine.2009.09.134; PMID: 19853680
- Dlugovitzky D, Notario R, Martinel-Lamas D, Fiorenza G, Farroni M, Bogue C, et al. Immunotherapy with oral, heat-killed, Mycobacterium vaccae in patients with moderate to advanced pulmonary tuberculosis. Immunotherapy 2010; 2:159 - 69; http://dx.doi.org/10.2217/imt.09.90; PMID: 20635925
- Jenkins PA, Campbell IA, Banks J, Gelder CM, Prescott RJ, Smith AP. Clarithromycin vs ciprofloxacin as adjuncts to rifampicin and ethambutol in treating opportunist mycobacterial lung diseases and an assessment of Mycobacterium vaccae immunotherapy. Thorax 2008; 63:627 - 34; http://dx.doi.org/10.1136/thx.2007.087999; PMID: 18250184
- Vilaplana C, Gil O, Cáceres N, Pinto S, Díaz J, Cardona PJ. Prophylactic effect of a therapeutic vaccine against TB based on fragments of Mycobacterium tuberculosis.. PLoS One 2011; 6:e20404; http://dx.doi.org/10.1371/journal.pone.0020404; PMID: 21647222
- Lowrie DB, Tascon RE, Bonato VL, Lima VM, Faccioli LH, Stavropoulos E, et al. Therapy of tuberculosis in mice by DNA vaccination. Nature 1999; 400:269 - 71; http://dx.doi.org/10.1038/22326; PMID: 10421369
- Ha SJ, Jeon BY, Youn JI, Kim SC, Cho SN, Sung YC. Protective effect of DNA vaccine during chemotherapy on reactivation and reinfection of Mycobacterium tuberculosis.. Gene Ther 2005; 12:634 - 8; http://dx.doi.org/10.1038/sj.gt.3302465; PMID: 15690060
- Henao-Tamayo M, Palaniswamy GS, Smith EE, Shanley CA, Wang B, Orme IM, et al. Post-exposure vaccination against Mycobacterium tuberculosis.. Tuberculosis (Edinb) 2009; 89:142 - 8; http://dx.doi.org/10.1016/j.tube.2009.01.002; PMID: 19264552
- Aagaard C, Hoang T, Dietrich J, Cardona PJ, Izzo A, Dolganov G, et al. A multistage tuberculosis vaccine that confers efficient protection before and after exposure. Nat Med 2011; 17:189 - 94; http://dx.doi.org/10.1038/nm.2285; PMID: 21258338
- Ahn SS, Jeon BY, Kim KS, Kwack JY, Lee EG, Park KS, et al. Mtb32 is a promising tuberculosis antigen for DNA vaccination in pre- and post-exposure mouse models. Gene Ther 2012; 19:570 - 5; http://dx.doi.org/10.1038/gt.2011.140; PMID: 21956689
- Yang XY, Chen QF, Li YP, Wu SM. Mycobacterium vaccae as adjuvant therapy to anti-tuberculosis chemotherapy in never-treated tuberculosis patients: a meta-analysis. PLoS One 2011; 6:e23826; http://dx.doi.org/10.1371/journal.pone.0023826; PMID: 21909406
- Guo S, Zhao J. Immunotherapy for tuberculosis: what’s the better choice?. Front Biosci 2012; 17:2684 - 90; http://dx.doi.org/10.2741/4079; PMID: 22652806
- Lockhart E, Green AM, Flynn JL. IL-17 production is dominated by gammadelta T cells rather than CD4 T cells during Mycobacterium tuberculosis infection. J Immunol 2006; 177:4662 - 9; PMID: 16982905
- Bafica A, Scanga CA, Feng CG, Leifer C, Cheever A, Sher A. TLR9 regulates Th1 responses and cooperates with TLR2 in mediating optimal resistance to Mycobacterium tuberculosis.. J Exp Med 2005; 202:1715 - 24; http://dx.doi.org/10.1084/jem.20051782; PMID: 16365150
- Jung YJ, Ryan L, LaCourse R, North RJ. Properties and protective value of the secondary versus primary T helper type 1 response to airborne Mycobacterium tuberculosis infection in mice. J Exp Med 2005; 201:1915 - 24; http://dx.doi.org/10.1084/jem.20050265; PMID: 15955839
- Arriaga AK, Orozco EH, Aguilar LD, Rook GA, Hernández Pando R. Immunological and pathological comparative analysis between experimental latent tuberculous infection and progressive pulmonary tuberculosis. Clin Exp Immunol 2002; 128:229 - 37; http://dx.doi.org/10.1046/j.1365-2249.2002.01832.x; PMID: 11985512
- Morais Fonseca D, Rosada RS, e Paula MO, Wowk PF, Franco LH, Soares EG, et al. Experimental tuberculosis: designing a better model to test vaccines against tuberculosis. Tuberculosis (Edinb) 2010; 90:135 - 42; http://dx.doi.org/10.1016/j.tube.2010.01.005; PMID: 20188631
- Cooper AM, Dalton DK, Stewart TA, Griffin JP, Russell DG, Orme IM. Disseminated tuberculosis in interferon gamma gene-disrupted mice. J Exp Med 1993; 178:2243 - 7; http://dx.doi.org/10.1084/jem.178.6.2243; PMID: 8245795
- Flynn JL, Chan J, Triebold KJ, Dalton DK, Stewart TA, Bloom BR. An essential role for interferon gamma in resistance to Mycobacterium tuberculosis infection. J Exp Med 1993; 178:2249 - 54; http://dx.doi.org/10.1084/jem.178.6.2249; PMID: 7504064
- Philips JA, Ernst JD. Tuberculosis pathogenesis and immunity. Annu Rev Pathol 2012; 7:353 - 84; http://dx.doi.org/10.1146/annurev-pathol-011811-132458; PMID: 22054143
- Jurado JO, Pasquinelli V, Alvarez IB, Peña D, Rovetta AI, Tateosian NL, et al. IL-17 and IFN-γ expression in lymphocytes from patients with active tuberculosis correlates with the severity of the disease. J Leukoc Biol 2012; 91:991 - 1002; http://dx.doi.org/10.1189/jlb.1211619; PMID: 22416258
- Cruz A, Fraga AG, Fountain JJ, Rangel-Moreno J, Torrado E, Saraiva M, et al. Pathological role of interleukin 17 in mice subjected to repeated BCG vaccination after infection with Mycobacterium tuberculosis.. J Exp Med 2010; 207:1609 - 16; http://dx.doi.org/10.1084/jem.20100265; PMID: 20624887
- Cruz A, Khader SA, Torrado E, Fraga A, Pearl JE, Pedrosa J, et al. Cutting edge: IFN-gamma regulates the induction and expansion of IL-17-producing CD4 T cells during mycobacterial infection. J Immunol 2006; 177:1416 - 20; PMID: 16849446
- Torrado E, Cooper AM. IL-17 and Th17 cells in tuberculosis. Cytokine Growth Factor Rev 2010; 21:455 - 62; http://dx.doi.org/10.1016/j.cytogfr.2010.10.004; PMID: 21075039
- Dietrich J, Aagaard C, Leah R, Olsen AW, Stryhn A, Doherty TM, et al. Exchanging ESAT6 with TB10.4 in an Ag85B fusion molecule-based tuberculosis subunit vaccine: efficient protection and ESAT6-based sensitive monitoring of vaccine efficacy. J Immunol 2005; 174:6332 - 9; PMID: 15879133
- Hervas-Stubbs S, Majlessi L, Simsova M, Morova J, Rojas MJ, Nouzé C, et al. High frequency of CD4+ T cells specific for the TB10.4 protein correlates with protection against Mycobacterium tuberculosis infection. Infect Immun 2006; 74:3396 - 407; http://dx.doi.org/10.1128/IAI.02086-05; PMID: 16714570
- Romano M, Roupie V, Wang XM, Denis O, Jurion F, Adnet PY, et al. Immunogenicity and protective efficacy of tuberculosis DNA vaccines combining mycolyl-transferase Ag85A and phosphate transport receptor PstS-3. Immunology 2006; 118:321 - 32; http://dx.doi.org/10.1111/j.1365-2567.2006.02373.x; PMID: 16827893
- Sable SB, Cheruvu M, Nandakumar S, Sharma S, Bandyopadhyay K, Kellar KL, et al. Cellular immune responses to nine Mycobacterium tuberculosis vaccine candidates following intranasal vaccination. PLoS One 2011; 6:e22718; http://dx.doi.org/10.1371/journal.pone.0022718; PMID: 21799939
- Cervantes-Villagrana AR, Hernández-Pando R, Biragyn A, Castañeda-Delgado J, Bodogai M, Martínez-Fierro M, et al. Prime-boost BCG vaccination with DNA vaccines based in β-defensin-2 and mycobacterial antigens ESAT6 or Ag85B improve protection in a tuberculosis experimental model. Vaccine 2013; 31:676 - 84; http://dx.doi.org/10.1016/j.vaccine.2012.11.042; PMID: 23196205
- Fontenot JD, Gavin MA, Rudensky AY. Foxp3 programs the development and function of CD4+CD25+ regulatory T cells. Nat Immunol 2003; 4:330 - 6; http://dx.doi.org/10.1038/ni904; PMID: 12612578
- Ribeiro-Rodrigues R, Resende Co T, Rojas R, Toossi Z, Dietze R, Boom WH, et al. A role for CD4+CD25+ T cells in regulation of the immune response during human tuberculosis. Clin Exp Immunol 2006; 144:25 - 34; http://dx.doi.org/10.1111/j.1365-2249.2006.03027.x; PMID: 16542361
- Chen X, Zhou B, Li M, Deng Q, Wu X, Le X, et al. CD4(+)CD25(+)FoxP3(+) regulatory T cells suppress Mycobacterium tuberculosis immunity in patients with active disease. Clin Immunol 2007; 123:50 - 9; http://dx.doi.org/10.1016/j.clim.2006.11.009; PMID: 17234458
- Fedatto PF, Sérgio CA, Paula MO, Gembre AF, Franco LH, Wowk PF, et al. Protection conferred by heterologous vaccination against tuberculosis is dependent on the ratio of CD4(+) /CD4(+) Foxp3(+) cells. Immunology 2012; 137:239 - 48; http://dx.doi.org/10.1111/imm.12006; PMID: 22891805
- Kaufmann SH, Väth U, Thole JE, Van Embden JD, Emmrich F. Enumeration of T cells reactive with Mycobacterium tuberculosis organisms and specific for the recombinant mycobacterial 64-kDa protein. Eur J Immunol 1987; 17:351 - 7; http://dx.doi.org/10.1002/eji.1830170308; PMID: 3106059
- Antas PR, Cardoso FL, Pereira KC, Franken KL, Cunha KS, Klatser P, et al. T cell immune responses to mycobacterial antigens in Brazilian tuberculosis patients and controls. Trans R Soc Trop Med Hyg 2005; 99:699 - 707; http://dx.doi.org/10.1016/j.trstmh.2005.05.002; PMID: 15993451
- Leyten EM, Lin MY, Franken KL, Friggen AH, Prins C, van Meijgaarden KE, et al. Human T-cell responses to 25 novel antigens encoded by genes of the dormancy regulon of Mycobacterium tuberculosis.. Microbes Infect 2006; 8:2052 - 60; http://dx.doi.org/10.1016/j.micinf.2006.03.018; PMID: 16931093
- Chegou NN, Black GF, Loxton AG, Stanley K, Essone PN, Klein MR, et al. Potential of novel Mycobacterium tuberculosis infection phase-dependent antigens in the diagnosis of TB disease in a high burden setting. BMC Infect Dis 2012; 12:10; http://dx.doi.org/10.1186/1471-2334-12-10; PMID: 22260319
- Parida SK, Kaufmann SH. The quest for biomarkers in tuberculosis. Drug Discov Today 2010; 15:148 - 57; http://dx.doi.org/10.1016/j.drudis.2009.10.005; PMID: 19854295
- Brandes M, Willimann K, Moser B. Professional antigen-presentation function by human gammadelta T Cells. Science 2005; 309:264 - 8; http://dx.doi.org/10.1126/science.1110267; PMID: 15933162
- Casetti R, Agrati C, Wallace M, Sacchi A, Martini F, Martino A, et al. Cutting edge: TGF-beta1 and IL-15 Induce FOXP3+ gammadelta regulatory T cells in the presence of antigen stimulation. J Immunol 2009; 183:3574 - 7; http://dx.doi.org/10.4049/jimmunol.0901334; PMID: 19710458
- Davey MS, Lin CY, Roberts GW, Heuston S, Brown AC, Chess JA, et al. Human neutrophil clearance of bacterial pathogens triggers anti-microbial γδ T cell responses in early infection. PLoS Pathog 2011; 7:e1002040; http://dx.doi.org/10.1371/journal.ppat.1002040; PMID: 21589907
- Tuomela M, Stanescu I, Krohn K. Validation overview of bio-analytical methods. Gene Ther 2005; 12:Suppl 1 S131 - 8; http://dx.doi.org/10.1038/sj.gt.3302627; PMID: 16231045
- Rosada RS, de la Torre LG, Frantz FG, Trombone AP, Zárate-Bladés CR, Fonseca DM, et al. Protection against tuberculosis by a single intranasal administration of DNA-hsp65 vaccine complexed with cationic liposomes. BMC Immunol 2008; 9:38; http://dx.doi.org/10.1186/1471-2172-9-38; PMID: 18647414
- Hsia CC, Hyde DM, Ochs M, Weibel ER, ATS/ERS Joint Task Force on Quantitative Assessment of Lung Structure. An official research policy statement of the American Thoracic Society/European Respiratory Society: standards for quantitative assessment of lung structure. Am J Respir Crit Care Med 2010; 181:394 - 418; http://dx.doi.org/10.1164/rccm.200809-1522ST; PMID: 20130146
- Cohen J. Infectious Disease. Approval of novel TB drug celebrated-with restraint. Science 2013; 339:130; http://dx.doi.org/10.1126/science.339.6116.130; PMID: 23307714