
Abstract
It has been reported that lipopeptides can be used to elicit cytotoxic T lymphocyte (CTL) responses against viral diseases and cancer. In our previous study, we determined that mono-palmitoylated peptides can enhance anti-tumor responses in the absence of adjuvant activity. To investigate whether di-palmitoylated peptides with TLR2 agonist activity are able to induce anti-tumor immunity, we synthesized a di-palmitic acid-conjugated long peptide that contains a murine CTL epitope of HPV E749–57 (Pam2IDG). Pam2IDG stimulated the maturation of bone marrow-derived dendritic cells (BMDCs) through TLR2/6. After immunization, Pam2IDG induced higher levels of T cell responses than those obtained with its non-lipidated counterpart (IDG). In the prophylactic model, Pam2IDG immunization completely inhibited tumor growth, whereas IDG immunization was unable to inhibit tumor growth. However, Pam2IDG immunization could not effectively inhibit the growth of established tumors. Therefore, we further investigated whether the depletion of immunosuppressive factors could improve the therapeutic effects of Pam2IDG. Our data indicate that treatment with Pam2IDG combined with clodronate/liposome delays tumor growth and increases the survival rate. We also observed that the therapeutic effects of Pam2IDG are improved by diminishing the function of tumor-associate macrophages (TAMs) and through the use of an IL10 receptor blocking antibody or a Cyclooxygenase 2 (Cox-2) inhibitor. In conclusion, the depletion of TAMs may enhance the anti-tumor immunity of a TLR2 agonist-conjugated peptide.
Introduction
Cervical cancer is associated with the life-threatening human papillomavirus (HPV).Citation1 Although current prophylactic vaccines effectively prevent HPV infection and decrease the incidence of cervical cancer, there are no therapeutic effects against pre-existing HPV infection and tumor progression.Citation2 During HPV infection, E6 and E7 oncoproteins control the cell cycle to transform cells into malignancy.Citation3 Therefore, HPV E6 and E7 oncoproteins have been identified as candidates of therapeutic vaccines to induce tumor regression.Citation4,5 Recently, mixed synthetic long peptides derived from HPV16 E6 and E7 have exhibited promising effects in women with HPV16-positive and grade 3 vulvar intraepithelial neoplasia.Citation6 In animals, a long peptide that contains both cytotoxic T (Tc) and helper T cell (Th) epitopes of HPV 16 was found to effectively induce HPV 16-expressing tumor regression.Citation7 Because it is easy to manufacture a synthetic peptide that satisfies the regulatory requirements, the above-mentioned finding entails a major advancement in the strategy used to treat human cancer.
It is known that peptide-based vaccines are a safe and feasible approach for the induction of CTL responses against cancer. The major disadvantage is their low immunogenicity, which makes it necessary to use adjuvants. Therefore, peptide-based vaccines have been developed as self-adjuvants with an immune stimulator to enhance their immunogenicity.Citation8 It has been reported that synthetic lipopeptides induce both humoral and cellular immune responses without conventional adjuvants.Citation9,10 The mono-palmitoylated peptides derived from viral or tumor-associated antigens are able to effectively elicit antigen-specific cytotoxic T lymphocyte (CTL) activities.Citation11-14 However, mono-palmitoylated peptides do not have adjuvant activity but do exhibit increased antigen-cross presentation ability.Citation13,14 In contrast, di-palmitoylated peptides activate dendritic cells (DCs) and enhance Th1 immune responses through TLR2.Citation15-17 Thus, synthetic TLR2 agonist-linked antigens can target and activate DCs to induce effective CTL responses.Citation18,19
Although immune-stimulator conjugated peptides enhance antigen-specific CTL responses, the tumor-infiltrating immunosuppressive cells, tumor-associated macrophages (TAMs), and myeloid-derived suppressor cells (MDSCs) may block their killing effects, resulting in so-called immune escapes.Citation20 Macrophages are innate immune cells that potentially differentiate into conventional M1 macrophages for pathogen clearance or alternative M2 macrophages for tissue healing with immunosuppressive ability.Citation21 During tumor progression, tumors induce macrophages to differentiate into IL-10-producing M2-like TAMs to inhibit CTL anti-tumor responses.Citation22 Many reports have demonstrated that TAMs are potential immune escape mechanisms of HPV-associated cancer.Citation23,24 Macrophage depletion by clodronate/liposome has been found to delay tumor growth.Citation25,26 Cyclooxygenase 2 (Cox-2) inhibitors have been found to be able to block the differentiation of TAMs and induce tumor regression.Citation27,28 To develop efficient therapeutic approaches for HPV-associated cancer, it is necessary to both increase CTLs and reduce tumor-infiltrating TAMs.
In this study, we aimed to evaluate whether a TLR2 agonist conjugated-peptide can induce the synergistic regression of an established large tumor through the depletion of TAMs. The self-adjuvanting di-palmitoylated peptide synthesized in the present study contains a murine CTL epitope derived from HPV E749–57 (Pam2IDG). The Pam2IDG epitope activate dendritic cells through TLR2/6 and induced higher levels of CTL responses than its non-lipidated counterpart IDG. The anti-tumor effects are limited in small tumors but enhanced in large tumors. Furthermore, the depletion of macrophages in the presence of a Cox-2 inhibitor may further delay tumor growth.
Results
Pam2IDG induces BMDCs maturation through TLR2/6
Although it has been suggested that di-palmitoylated peptides engage TLR2/6 to activate antigen-presenting cells, the peptide length or composition may have diverse co-receptor usages.Citation29,30 To confirm that Pam2IDG is able to activate antigen-presenting cells through TLR2, BMDCs of WT or TLR-knockout (KO) mice were used to characterize the activity of pam2IDG. To elucidate whether Pam2IDG promotes BMDC maturation, BMDCs were stimulated with Pam2IDG or IDG for 18 h to determine the level of IL-12p40 secretion and the expression of co-stimulatory molecules, namely CD40, CD80, CD86, and MHC class II. The Pam2IDG-stimulated BMDCs secrete IL-12p40 in a dose-dependent manner (). The expression levels of CD40, CD80, and CD86 in the Pam2IDG-stimulated BMDCs are 1.5- to 2- fold higher than those of the IDG-stimulated BMDCs (). To determine the co-receptor usage of TLR2, BMDCs from WT, TLR1KO, TLR2KO, and TLR6KO mice were stimulated with Pam2IDG or IDG.
Figure 1. Pam2IDG enhances BMDC maturation and CTL activities to improve anti-tumor immunity. BMDCs were treated with 1 μM IDG or Pam2IDG at the indicated concentration for 18 h. (A) The level of IL-12p40 in the supernatant was determined by ELISA. (B) The mean fluorescence intensities of the surface molecules were analyzed by flow cytometry. Relative MFI = (MFI of IDG- or Pam2IDG-treated cells / MFI of untreated cells) x 100%. BMDCs cultured from WT, TLR1KO, TLR2KO, and TLR6KO mice were treated with 1 μM IDG, Pam2IDG, or 100 ng/ml LPS at the indicated concentration for 18 h. The levels of (C) IL-12p40 and (D) IL-6 in the supernatant were determined by ELISA.
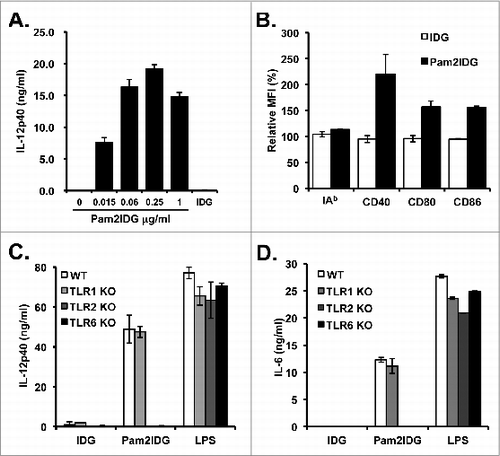
To demonstrate whether TLR2/6 is necessary for BMDC maturation after Pam2IDG treatment, BMDCs cultured from WT, TLR1KO, TLR2KO, and TLR6KO mice were pulsed with 1 μM Pam2IDG for 18 h, and the levels of IL-12p40 and IL-6 in the culture supernatants were determined by ELISA. Pam2IDG enhanced the secretion of IL-12p40 and IL-6 from WT and TLR1KO BMDCs but not TLR2KO and TLR6KO BMDCs ( and D). These data indicate that TLR2/6 is required for Pam2IDG to promote BMDC maturation.
Pam2IDG enhances CTL responses via TLR2/6
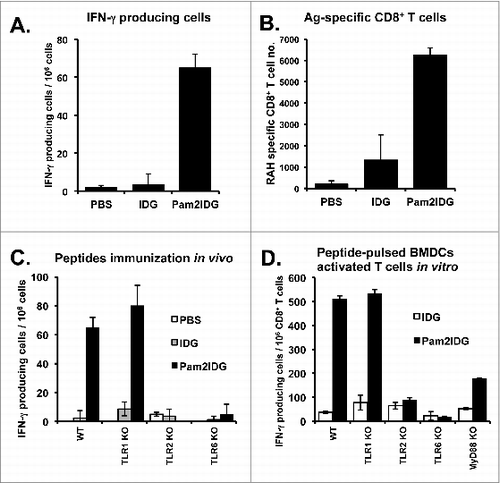
We first demonstrated that Pam2IDG can enhance BMDC maturation. We then further explored whether Pam2IDG can induce antigen-specific CTL responses. To demonstrate the CTL responses induced by Pam2IDG immunization, C57BL/6 mice were immunized with 10 μg of IDG or Pam2IDG via the footpad on days 0 and 5, and these lymphocytes were then re-stimulated with RAH to determine the numbers of IFN-γ-producing cells by ELISPOTs. Pam2IDG immunization was found to increase the number of IFN-γ producing cells after re-stimulation with RAH compared with the numbers obtained with IDG immunization (). The results indicate that Pam2IDG immunization can elicit more antigen-specific CTLs. To further analyze the numbers of RAH-specific CD8+ CTLs, lymphocytes collected from IDG- and Pam2IDG-immunized mice were double-stained with FITC-conjugated anti-CD8 antibody and PE-conjugated RAH tetramer, and the percentage of the antigen-specific CTL population was then analyzed by flow cytometry. Pam2IDG immunization resulted in a 2.5- to 3-fold increase in the numbers of CTLs compared with IDG (). Moreover, to investigate whether CTL induction by Pam2IDG immunization requires TLR2/6, WT, TLR1KO, TLR2KO, and TLR6KO mice were immunized with IDG or Pam2IDG respectively, and the numbers of IFN-γ-producing cells were then determined by ELISPOT, as shown in . Pam2IDG immunization was found to elicit antigen-specific CTL responses in WT and TLR1KO mice. However, TLR2KO and TLR6KO mice exhibited impaired CTL induction in response to Pam2IDG immunization. To further investigate whether Pam2IDG-pulsed BMDCs can enhance CTL cross-priming via the TLR2/6 pathway, BMDCs cultured from WT, TLR1KO, TLR2KO, TLR6KO, or MyD88KO mice were pulsed with 1 μM IDG or Pam2IDG for 2.5 h. After washing out the free peptides, the CD8+ T cells purified from RAH/IFA-immunized mice were co-cultured with peptide-pulsed BMDCs for 2 d, and the number of IFN-γ-producing cells was then determined by ELISPOT. Pam2IDG-pulsed WT and -TLR1KO BMDCs were found to activate CD8+ T cells to secrete IFN-γ, whereas TLR2KO, TLR6KO, or MyD88KO BMDCs were unable to achieve this activation (). Therefore, TLR2/6 and MyD88 are necessary for the enhancement of CTL cross-priming by Pam2IDG-pulsed BMDCs.
Figure 2. Pam2IDG increases the number of IFN-γ-producing cells. C57BL/6 naïve mice were immunized with 10 μg of IDG or Pam2IDG via the footpad on days 0 and 5. On day 10, the lymphocytes were collected from inguinal lymph nodes. (A) A total of 2 × 105 lymphocytes from each treatment group were re-stimulated with 5 μg/ml RAH or control peptide in anti-IFN-γ antibody-coated plates for 48 h. The IFN-γ-producing cells were examined by ELISPOT. (B) The lymphocytes were double-stained with RAH tetramer (Tet-RAH) and anti-CD8 antibody and analyzed by flow cytometry. The data are presented as the cell numbers of Tet-RAH+ CD8+ T cells from two independent experiments. (C) In vivo analysis of WT, TLR1KO, TLR2KO, and TLR6KO mice immunized as in (A), and the numbers of IFN-γ-producing cells were then examined by ELISPOT. (D) In vitro analysis of WT, TLR1KO, TLR2KO, TLR6KO and MyD88KO BMDCs pulsed with 1 μM IDG or Pam2IDG for 2.5 h (the free peptides were then washed out). A total of 2 × 104 peptide-pulsed BMDCs were co-cultured with 2 × 105 responder cells, which were purified from CD8+ T cells from the spleens of RAH/IFA-immunized mice. After 48 h, the IFN-γ-producing cells were examined by ELISPOT.
Pam2IDG can inhibit TC-1 tumor growth but does not inhibit large established tumors
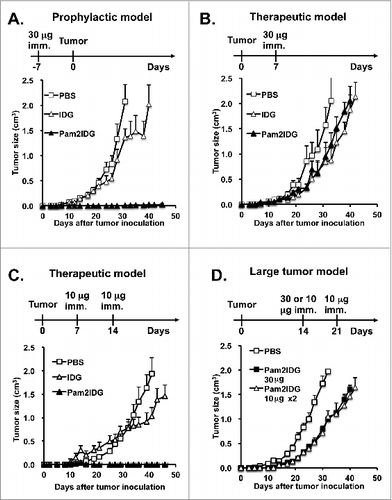
To evaluate the anti-tumor effect of Pam2IDG on the TC-1 tumor model, C57BL/6 mice were immunized with 30 μg of IDG or Pam2IDG and inoculated with 2 × 105 TC-1 tumor cells seven days after immunization. Pam2IDG immunization significantly inhibited TC-1 tumor growth until day 45, and this effect was not observed with IDG immunization (). In contrast, to evaluate the therapeutic effect of Pam2IDG, the mice were inoculated with TC-1 tumor and then immunized with 30 μg of IDG or Pam2IDG seven days after inoculation (palpable tumor was observed at this time). The average tumor volume of the Pam2IDG-immunized mice was 0.94 ± 0.13 cm3, which it not significantly different compared with that observed in the IDG-immunized mice (0.85 ± 0.28 cm3) (). However, the palpable tumor could be inhibited by immunization with 10 μg of Pam2IDDG on days 7 and 14 (). To further investigate the therapeutic effects of this peptide on a large tumor, the treatment was initiated on day 14 (the tumor diameter was 6–8 mm). A single dose of 30 μg of Pam2IDG or two doses of 10 μg of Pam2IDG administered at a 7-d interval did not completely inhibit the tumor growth but did delay the growth of the tumor (). Therefore, we hypothesize that Pam2IDG immunization can induce the regression of smaller but not large tumors.
Figure 3. Pam2IDG immunization inhibits palpable tumor growth. C57BL/6 naïve mice were inoculated subcutaneously with 2 × 105 TC-1 tumor cells in the right leg. (A) Seven days before tumor inoculation, the mice were subcutaneously immunized with PBS or 30 μg of IDG or Pam2IDG (n = 5 in each group). (B) The TC-1 tumor-bearing mice were immunized subcutaneously with PBS or 30 μg of IDG or Pam2ID on day 7 (n = 5 in each group). (C) The TC-1 tumor-bearing mice were subcutaneously immunized with PBS or 10 μg of IDG or Pam2IDG on days 7 and 14 (n = 6 in each group). (D) The TC-1 tumor-bearing mice were immunized with 30 μg of Pam2IDG on day 14 and 10 μg of Pam2IDG on days 14 and 21 (n = 6 in each group).
Depletion of macrophages improves the therapeutic effects of Pam2IDG
TAMs play an important role in the suppression of CTLs against tumor cells. To investigate whether the depletion of TAMs is able to enhance the therapeutic effects of Pam2IDG, the mice were treated with 1 mg of clodronate/liposome via i.p. injection on day 13 to deplete the macrophages and then immunized with or without 30 μg of Pam2IDG on day 14. The average tumor size of the Pam2IDG-and-clodronate-treated mice (0.32 ± 0.07 cm3) was significantly smaller than the average tumor size of the Pam2IDG-immunized mice (0.79 ± 0.07 cm3) on day 30. In contrast, the tumor size of the clodronate-treated mice was not significant different compared with that of the PBS-treated mice (1.68 ± 0.04 cm3 vs. 1.72 ± 0.15 cm3) on day 30 (). All of the mice in the PBS control and clodronate-treated groups were dead before day 35. The Pam2IDG -treated mice were dead before day 47 (). It is noteworthy that the Pam2IDG-and-clodronate-treated mice exhibited a 50% survival rate on day 60. Therefore, these data indicate that the depletion of TAMs enhances the anti-tumor effects of Pam2IDG immunization.
Figure 4. Therapeutic effects of Pam2IDG are improved by the reduction of immunosuppressive factors. C57BL/6 naïve mice were inoculated subcutaneously with 2 × 105 TC-1 tumor cells in the right leg. (A and B) TC-1 tumor-bearing mice were intraperitoneally injected with 1 mg of clodronate/liposome on day 13 and subcutaneously immunized with 30 μg of Pam2IDG on day 14 (n = 10) in each group; Pam2IDG combined with clodronate vs. Pam2IDG, P < 0.01). (C and D) TC-1 tumor-bearing mice were intraperitoneally injected with 200 μg of anti-IL10R antibody on day 13 and subcutaneously immunized with 30 μg of Pam2IDG on day 14 (n = 6 in each group; Pam2IDG combined with anti-IL10R antibody vs. Pam2IDG, P < 0.01). (E and F) TC-1 tumor-bearing mice were intraperitoneally injected with 0.5 mg/kg celecoxib at 2-d intervals until day 30 and subcutaneously immunized with 30 μg of Pam2IDG on day 14 (n = 10 in each group; Pam2IDG combined with celecoxib vs. Pam2IDG, P < 0.01)
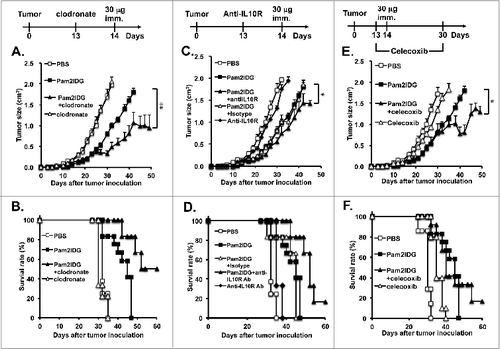
To further investigate whether the M2 macrophage-associated cytokine IL-10 reduces the anti-tumor effects of Pam2IDG, tumor-bearing mice were injected with 200 μg of IL-10R antibody or isotype control via i.p. on day 13 and then immunized with 30 μg of Pam2IDG on day 14. The tumor size of the Pam2IDG-and-anti-IL10R antibody-treated mice was smaller than the tumor size of the Pam2IDG-immunized mice (0.56 ± 0.09 vs.0.79 ± 0.07 cm3) and the Pam2IDG-and-isotype control antibody-treated mice (0.56 ± 0.09 vs. 0.81 ± 0.1 cm3) on day 30 (). The survival rate of the Pam2IDG-and-anti-IL10R antibody-treated mice was 66% on day 50 and decreased to 16% on day 60; in contrast, all of the mice in the Pam2IDG treatment group were dead on day 45 (). Therefore, we suggest that IL-10 can reduce the antitumor effects of Pam2IDG immunization.
To further investigate whether blocking the Cox-2 pathway enhances the antitumor effects of Pam2IDG, tumor-bearing mice were injected with 0.5 mg/kg celecoxib via i.p. from day 13 to 30 at 2-d intervals and then immunized with or without 30 μg of Pam2IDG on day 14. The tumor size of the Pam2IDG-and-celecoxib-treated mice was smaller than the tumor size of the Pam2IDG-immunized mice (0.78 ± 0.15 vs. 1.6 ± 0.09 cm3) on day 40. We also observed that the survival rate of the Pam2IDG-and-celecoxib-treated mice was 16% on day 60 (). Hence, we hypothesize that the inhibition of Cox-2 may promote the antitumor effects of Pam2IDG. In summary, the elimination of TAMs, IL-10, or Cox-2 enhances the anti-tumor effects of Pam2IDG.
Clodronate can alter the ratio of M1 to M2 macrophages to improve the effects of Pam2IDG immunization
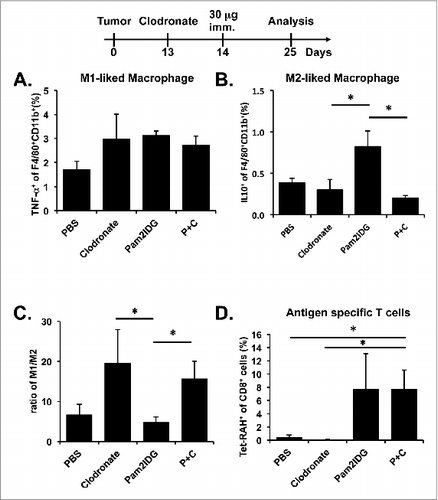
To investigate whether clodronate alters the M1 and M2 macrophage populations, tumor-infiltrating cells were treated with LPS for 4 h and then stained with anti-F4/80, anti-CD11b, anti-CD45, anti-TNF-α, or anti-IL10 antibodies. The M1 macrophage population was slightly increased in the mice treated with clodronate, Pam2IDG, and both Pam2IDG and clodronate compared with the mice treated with PBS (). However, the M2 macrophage population was increased in the Pam2IDG-treated mice compare with the mice treated with clodronate or both Pam2IDG and clodronate (). The ratio of M1 to M2 macrophages in the clodronate- and Pam2IDG-and-clodronate-treated mice was higher than that observed in the PBS- and Pam2IDG-treated mice (). Our results indicate that clodronate can alter the M1 and M2 macrophage population in the tumor micro-environment. To study whether the depletion of TAMs leads to an increase in antigen-specific CTLs in tumors, tumor-infiltrating cells were stained with anti-CD8 or anti-CD45 antibody or antigen-specific MHC class I tetramer (Tet-RAH) and then analyzed by flow cytometry. The numbers of antigen-specific CTLs in the tumor infiltrate are increased by Pam2IDG and by Pam2IDG combined with clodronate but by clodronate alone (). These data support the hypothesis that clodronate may reduce TAMs and change the ratio of M1/M2 to increase the cancer-killing ability of CTLs.
Figure 5. Pam2IDG immunization combined with clodronate increases tumor-infiltrating M1/M2 macrophage ratio. Five C57BL/6 naïve mice in each group were subcutaneously inoculated with 2 × 105 TC-1 tumor cells in the right leg. The TC-1 tumor-bearing mice were intraperitoneally injected with 1 mg of clodronate/liposome on day 13 and subcutaneously immunized with or without 30 μg of Pam2IDG on day 14. The tumor infiltrates were analyzed on day 25 by flow cytometry. (A) The TNF-α− and (B) IL10-secreting F4/80+ CD11b+ cells indicate the M1 and M2 macrophages, respectively. (C) The ratio was calculated as the percentage of M1 macrophages divided by the percentage of M2 macrophages. (D) The RAH-specific CTLs were detected using a tetramer. P+C: mice were treated with clodronate and then immunized with Pam2IDG.
Discussion
Even though antitumor immunity can be induced by a TLR2 agonist-conjugated long peptide, this treatment does not efficiently eliminate large tumors. One of the major obstacles of cancer immunotherapy is the recruitment of tumor-infiltrated immunosuppressive cells. Some reports have demonstrated that TAMs are potential immune escape mechanisms of HPV-associated cancer.Citation23,24 TAMs also express B7H4 which negative regulate T cell activation and are correlated with high numbers of FoxP3+ Tregs and PD-1 expressing T cells.Citation31 M2-like TAMs secrete IL-10 for inducing the Treg differentiation and PGE2 to increase the number of MDSCs.Citation28 These data indicated that TAMs are able to regulate T cells functions either directly or indirectly. Although the depletion of TAMs did not increase antigen-specific CTLs, the reduction of M2-like TAMs could increase CTLs function. Our recent report also showed that TLR9 agonist synergistically enhanced anti-tumor immunity of TLR2 agonist-conjugated protein and reduced the total numbers of TAMs. The TLR2 agonist-conjugated protein alone did not have effects on reduction of TAMs.Citation32 The combination of administrating macrophage-depleting drugs with immunotherapy is promising and feasible for improving anti-tumor immunity. This study has led to the hypothesis that the depletion of TAMs or associated factors may enhance therapeutic effects of TLR2 agonist-conjugated long peptide treatment. TLR2 agonist-conjugated peptides have been reported to induce CTL responses against viral infections and tumors.Citation33,34 However, it has been reported that the activation of TLR2 enhances regulatory T cell function.Citation35 Peptide immunization in the presence of a TLR7 agonist not only increases antigen-specific CTLs but also increases both MDSCs and Tregs.Citation36 The adjuvant-induced immunosuppressive cells limit the therapeutic effects of cancer vaccines. In this study, we observed that immunization with a TLR2 agonist-conjugated peptide increases the number of tumor-infiltrating M2-like macrophages (). M2-like macrophages are able to secrete IL-10, which has been shown to directly affect the function of antigen-presenting cells, downregulate the expression of Th1 cytokines, and induce Tregs.Citation37 The inhibition of IL-10 signaling by anti-IL10R antibodies is able to suppress the M2 phenotype differentiation and the induction of tumor regression.Citation38 Accordingly, we administered an anti-IL10R antibody before Pam2IDG immunization to increase the survival rate of mice with large tumors (). The results indicate that the reduction of tumor-associated immunosuppressive cells or IL-10 may overcome the adjuvant-induced immunosuppressive effects of cancer vaccines.
Prostaglandin E2 (PGE2) is synthesized from arachidonic acid by Cox and prostaglandin synthase. The production of PGE2 by Cox-2 is responsible for the differentiation of Tregs and the suppression of T cell function.Citation39 Eberstal showed that intratumoral Cox-2 inhibition enhances GM-CSF immunotherapy against tumors.Citation40 Cox-2 is expressed not only in immune cells but also in tumor cells and stromal cells.Citation41 We found that the administration of the Cox-2 inhibitor Celecoxib via i.p. every two days and a single dose of Pam2IDG significantly prolongs the survival time of tumor-bearing mice (). These data indicate that the Cox-2 inhibitor is able to enhance the therapeutic effects of cancer vaccines.
We found that the long peptide with TLR2 agonist activity, which was designed as a tumor therapeutic vaccine, not only stimulates BMDC activation but also cross-primes CTLs via TLR2/6. A single dose of Pam2IDG prevents TC-1 tumor growth and inhibits established tumor growth ( and ). The palpable tumors were found to regress after two immunizations with Pam2IDG. However, the growth of large tumors (6–8 mm in diameter) was delayed after immunization with Pam2IDG in combination with the depletion of TAMs. Although Pam2IDG immunization did not induce the regression of established tumors, Pam2IDG immunization combined with clodronate increased the ratio of the M1/M2 macrophage population and prolonged the survival time. Although macrophages were depleted by clodronate administration, clodronate did not abrogate the function of antigen-presenting cells.Citation42 However, the use of a high dose of clodronate to completely deplete the macrophages may impair the cross-priming of CTLs by dendritic cells.Citation43 In this study, we demonstrated that Pam2IDG combined with clodronate can induce tumor regression. Clodronate depletes not only macrophages but also CD11b+Gr-1+ myeloid-derived suppressor cells (MDSCs).Citation44 Although we did not find any significant decrease in the numbers of MDSCs in this tumor model post-inoculation (data not shown), Pam2IDG immunization combined with clodronate induced antigen-specific CTLs to the same degree as Pam2IDG alone (). Therefore, CTLs can be elicited by Pam2IDG immunization and reach the tumor site.
In this report, we demonstrated that TLR2 agonist-conjugated peptide could induce anti-tumor immunity in mouse model. To apply in human, we have to design peptides that contained human CTL epitopes. In our previous studies, we have identified the HLA-A11-restricted CTL epitopes of HPV 18, 52 and 58.Citation45-47 These long peptides could be synthesized with TLR2 agonist. The synthetic di-acyl lipopeptides are easy to manufacture for clinical used. Alternatively, we can design the lipopeptide mixture that contained different CTL epitopes to treat human. The mixtures of HPV16 E6 and E7-derived long peptides have been used in human clinical trials which showed promising results in VIN3 patients.Citation6 However, the therapeutic effect in invasive cancer is still a big challenge and certainly need new vaccine formulations. Here, we showed the liposomal clodronate, celecoxib or anti-IL10R is able to improve the therapeutic effects of TLR2 agonist-conjugated long peptide. In the future, the combination of mixtures of TLR2 agonist-conjugated long peptides and anti-immmunosuppressive drugs is feasible for cancer immunotherapy in human.
In conclusion, the reduction of immunosuppressive factors or cells enhances the effects of TLR2 agonist-conjugated peptide immunotherapy against cervical cancer. These results suggest that the targeting of tumor-suppressive cells or factors is necessary for successful cancer immunotherapy.
Materials and Methods
Animals
C57BL/6 mice were purchased from the National Animal Center in Taiwan. All of the mice were maintained in the Animal Center of the National Health Research Institutes (NHRI, Miaoli, Taiwan). TLR1KO, TLR2KO, TLR6KO, and MyD88KO mice were purchased from Oriental Bioservice, Inc. (Tokyo, Japan). All of the animal studies were approved by the Institutional Animal Care and Use Committee of the NHRI.
Peptides and di-palmitic peptides
The peptides (IDGPAGQAEPDRAHYNIVTFCCKC) and di-palmitic peptides (Pam2-KSS-IDGPAGQAEPDRAHYNIVTFCCKC) containing an H-2Db-restricted CTL epitope (amino acids 49–57, RAHYNIVTF) derived from the HPV16 E7 protein were purchased from GL Biochem (Shanghai, China). These peptides were dissolved in DMSO at 10 mg/ml and stored at −20°C.
BMDC generation and maturation analysis
The BMDCs were generated as previously described.Citation48 Briefly, bone marrow cells were collected from the femurs and tibiae and incubated with RBC lysis buffer (BioLegend, CA, USA) for 2 min to remove the RBCs. A total of 2 × 106 bone marrow cells were plated into 10-cm petri dishes in 10 ml of RPMI 1640 medium supplemented with 100 U/ml penicillin/streptomycin, 50 μM 2-mercaptoethanol (Sigma-Aldrich, China), 10% heat-inactivated FBS, and 20 ng/ml mouse rGM-CSF (PeproTech, NJ, USA). The cells were incubated at 37°C with 5% CO2. On day 3, 10 ml of fresh media was added, and the cells were cultured for an additional 3 d. On day 6, the detached cells in each dish were collected by washing with medium and counted to determine the cell number. To determinate the cytokine secretion of BMDCs, the supernatants from 1 × 106/ml BMDCs incubated with peptides at the indicated concentration or 100 ng/ml LPS (Sigma-Aldrich, China) for 18 h were collected. The production of IL-6 and IL-12p40 by BMDCs were determined by ELISA (eBioscience, San Diego, CA, USA). To analyze the expression of BMDC surface molecules, 5 × 105 BMDCs were re-suspended in staining buffer (PBS containing 1% FCS) and stained with 0.2 μg of anti-CD11c-FITC (N418, BioLegend), CD40-PE (3/23, BD Biosciences), CD80-PE (16–10A1, BD Biosciences), CD86-PE (GL-1, BD Biosciences), and IAb-PE (AF6–120.1, BioLegend) monoclonal antibodies at 4°C for 30 min. The BMDCs were acquired by a FACSCalibur (BD Biosciences), and the mean of fluorescence intensity of the gated CD11c+ cell populations was analyzed using the CellQuest Pro software.
ELISPOT assay
To determinate the IFN-γ production of the cytotoxic T cells, 96-well Multiscreen PI plates (Millipore, Billerica, MA, USA) were coated with rat anti-mouse IFN-γ antibody (clone R4–6A2; eBioscience) in PBS overnight at 4°C. After washing with PBS, the plates were blocked with 200 μl of RPMI-10 for 1 h at room temperature. A total of 2 × 105 lymphocytes were seeded per well and incubated with 5 μg/ml of RAH or control peptide (VVYRDSIPH) in 200 μl of RPMI for 48 h. The wells were sequentially washed three times with washing buffer (PBS containing 0.05% Tween 20) and then incubated for 4 h at room temperature with biotinylated anti-mouse IFN-γ antibody (4 μg/ml, clone XMG1.2, eBioscience) in assay diluent. After an additional wash, the wells were incubated with peroxidase-labeled streptavidin (2 μg/ml; eBioscience) for 15 min at room temperature. The spots were developed using freshly prepared substrate (ACE; Sigma, St. Louis, MO, USA) dissolved in distilled water. The IFN-γ-secreting T cells were measured using an ELISPOT reader (Cellular Technology Ltd.). The results are shown as the numbers of IFN-γ-secreting cells of RAH-stimulated group after subtracting the value of the control peptides-stimulated group.
Animal tumor model
To establish the HPV16 E6/E7 tumor model in C57BL/6 mice, we inoculated the animals with 2 × 105 TC-1 cells in the right leg. For the preventive vaccination, each mouse was subcutaneously immunized with 30 μg of peptide. To elucidate the therapeutic effect, the TC-1-bearing mice were immunized subcutaneously with PBS, IDG, or Pam2IDG on day 7 post-inoculation. To assess the therapeutic effect on an established tumor, the TC-1-bearing mice were immunized subcutaneously with PBS, IDG, or Pam2IDG on day 14 post-inoculation. The local tumor diameter was measured with calipers and monitored three times per week until 45–60 d post-inoculation. The tumor volume was calculated using the following formula: length × width × width / 2. When the tumor volumes became larger than 2 cm3, the mice were euthanatized. To deplete the macrophages, the mice were i.p. injected with 1 μg of clodrosome (Encapsula NanoScience, Nashville, TN, USA) on day 13 post-tumor inoculation. To diminish the IL-10 immunosuppressive effect, the mice were i.p. injected with 200 μg of 1B1.3a antibody (BioLegend) to block the IL-10 receptor on day 13 post-tumor inoculation. To inhibit the Cox-2 pathway in vivo, the mice were i.p. injected with 0.5 mg/kg Celecoxib (Sigma) three times a week from day 13 to day 30 post-tumor inoculation.
Analysis of tumor-infiltrating cells
To collect the TC-1 tumor infiltrates, the tumors from tumor-bearing mice were dissected on day 25. Scissors were used to chop the tumors into small 2- to 4-mm pieces, and the small pieces of the tumors were further ground into single cells though 70-mm cell strainers. The cells were washed and re-suspended in staining buffer. To detect the antigen-specific T cell population, the cells were stained with anti-CD8 (53–6.7, FITC, BioLegend) and anti-CD45 (EM-05, APC, GeneTex) antibodies and Tet-RAH (tetramer RAHYNIVTF, MBL, Japan) at room temperature. To analyze the macrophage population, the cells were stained with anti-CD11b (M1/70, PE, eBioscience), anti-Ly6G (RB6–8C5, PECy7, eBioscience), anti-F4/80 (BM8, FITC, BioLegend), and anti-CD45 antibodies. To detect the cytokine secretion, the cells were treated with 100 ng/ml LPS for 4 h and then stained with anti-TNF-α (MP6-XT22, PE-Cy5.5, BioLegend) or anti-IL-10 (JES5–16E3, PE-Cy5.5, BioLegend).
Statistical analysis
The statistical significance was evaluated by Student's t test (two-tailed) at the 5% level. The statistical significance of the tumor models treated with clodronate, celecoxib, and anti-IL10R antibody were analyzed by one-way ANOVA.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Authors’ Contributions
KYS performed the experiments with contributions from YCS, IHC and PC. KYS and YCS analyzed the data. KYS and SJL designed the experiments and wrote the manuscript.
Acknowledgments
The authors thank Johns Hopkins University for providing the TC-1 cells.
Funding
This work was supported by grants from the National Science Council awarded to S.J. Liu (NSC 98–2320-B-400-011-MY3 and NSC 101–2321-B-400-022-MY2).
References
- Cho HJ, Oh YK, Kim YB. Advances in human papilloma virus vaccines: a patent review. Expert opinion on therapeutic patents 2011; 21:295-309.
- Woodman CB, Collins SI, Young LS. The natural history of cervical HPV infection: unresolved issues. Nat Rev Cancer 2007; 7:11-22; PMID:17186016; http://dx.doi.org/10.1038/nrc2050
- Hebner CM, Laimins LA. Human papillomaviruses: basic mechanisms of pathogenesis and oncogenicity. Rev Med Virol 2006; 16:83-97; PMID:16287204; http://dx.doi.org/10.1002/rmv.488
- Hung CF, Ma B, Monie A, Tsen SW, Wu TC. Therapeutic human papillomavirus vaccines: current clinical trials and future directions. Expert Opin Biol Ther 2008; 8:421-39; PMID:18352847; http://dx.doi.org/10.1517/14712598.8.4.421
- Wieking BG, Vermeer DW, Spanos WC, Lee KM, Vermeer P, Lee WT, Xu Y, Gabitzsch ES, Balcaitis S, Balint JP Jr., et al. A non-oncogenic HPV 16 E6/E7 vaccine enhances treatment of HPV expressing tumors. Cancer Gene Ther 2012; 19:667-74; PMID:22918471; http://dx.doi.org/10.1038/cgt.2012.55
- Kenter GG, Welters MJ, Valentijn AR, Lowik MJ, Berends-van der Meer DM, Vloon AP, Essahsah F, Fathers LM, Offringa R, Drijfhout JW, et al. Vaccination against HPV-16 oncoproteins for vulvar intraepithelial neoplasia. N Engl J Med 2009; 361:1838-47; PMID:19890126; http://dx.doi.org/10.1056/NEJMoa0810097
- Zwaveling S, Ferreira Mota SC, Nouta J, Johnson M, Lipford GB, Offringa R, van der Burg SH, Melief CJ. Established human papillomavirus type 16-expressing tumors are effectively eradicated following vaccination with long peptides. J Immunol 2002; 169:350-8; PMID:12077264; http://dx.doi.org/10.4049/jimmunol.169.1.350
- Prajeeth CK, Jirmo AC, Krishnaswamy JK, Ebensen T, Guzman CA, Weiss S, Constabel H, Schmidt RE, Behrens GM. The synthetic TLR2 agonist BPPcysMPEG leads to efficient cross-priming against co-administered and linked antigens. Eur J Immunol 2010; 40:1272-83; PMID:20213735; http://dx.doi.org/10.1002/eji.200939790
- Andrieu M, Loing E, Desoutter JF, Connan F, Choppin J, Gras-Masse H, Hanau D, Dautry-Varsat A, Guillet JG, Hosmalin A. Endocytosis of an HIV-derived lipopeptide into human dendritic cells followed by class I-restricted CD8(+) T lymphocyte activation. Eur J Immunol 2000; 30:3256-65; PMID:11093141; http://dx.doi.org/10.1002/1521-4141(200011)30:11≤3256::AID-IMMU3256≥3.0.CO;2-H
- Gahéry-Ségard H, Pialoux G, Charmeteau B, Sermet S, Poncelet H, Raux M, Tartar A, Lévy JP, Gras-Masse H, Guillet JG. Multiepitopic B- and T-cell responses induced in humans by a human immunodeficiency virus type 1 lipopeptide vaccine. J Virol 2000; 74:1694-703; PMID:10644339; http://dx.doi.org/10.1128/JVI.74.4.1694-1703.2000
- Xu DH, Zhou CH, Xia YP, Qiu ZY, Wu YZ, Jia ZC, Zhou W. Cytotoxic T lymphocyte response induced by an improved synthetic lipopeptide vaccine against cervical cancer. Acta Pharmacol Sin 2007; 28:695-702; PMID:17439726; http://dx.doi.org/10.1111/j.1745-7254.2007.00538.x
- Baz A, Buttigieg K, Zeng W, Rizkalla M, Jackson DC, Groves P, Kelso A. Branched and linear lipopeptide vaccines have different effects on primary CD4+ and CD8+ T-cell activation but induce similar tumor-protective memory CD8+ T-cell responses. Vaccine 2008; 26:2570-9; PMID:18420312; http://dx.doi.org/10.1016/j.vaccine.2008.03.022
- Andrieu M, Desoutter JF, Loing E, Gaston J, Hanau D, Guillet JG, Hosmalin A. Two human immunodeficiency virus vaccinal lipopeptides follow different cross-presentation pathways in human dendritic cells. J Virol 2003; 77:1564-70; PMID:12502869; http://dx.doi.org/10.1128/JVI.77.2.1564-1570.2003
- Song YC, Chou AH, Homhuan A, Huang MH, Chiang SK, Shen KY, Chuang PW, Leng CH, Tao MH, Chong P, et al. Presentation of lipopeptide by dendritic cells induces anti-tumor responses via an endocytosis-independent pathway in vivo. J Leukoc Biol 2011; 90:323-32; PMID:21521754; http://dx.doi.org/10.1189/jlb.0111046
- Chua BY, Zeng W, Lau YF, Jackson DC. Comparison of lipopeptide-based immunocontraceptive vaccines containing different lipid groups. Vaccine 2007; 25:92-101; PMID:17055123; http://dx.doi.org/10.1016/j.vaccine.2006.07.012
- Azuma M, Sawahata R, Akao Y, Ebihara T, Yamazaki S, Matsumoto M, Hashimoto M, Fukase K, Fujimoto Y, Seya T. The peptide sequence of diacyl lipopeptides determines dendritic cell TLR2-mediated NK activation. PLoS One 2010; 5:5; PMID:20824059; http://dx.doi.org/10.1371/journal.pone.0012550
- Zeng W, Horrocks KJ, Robevska G, Wong CY, Azzopardi K, Tauschek M, Robins-Browne RM, Jackson DC. A modular approach to assembly of totally synthetic self-adjuvanting lipopeptide-based vaccines allows conformational epitope building. J Biol Chem 2011; 286:12944-51; PMID:21321114; http://dx.doi.org/10.1074/jbc.M111.227744
- Prajeeth CK, Jirmo AC, Krishnaswamy JK, Ebensen T, Guzman CA, Weiss S, Constabel H, Schmidt RE, Behrens GM. The synthetic TLR2 agonist BPPcysMPEG leads to efficient cross-priming against co-administered and linked antigens. Eur J Immunol 2010; 40:1272-83; PMID:20213735; http://dx.doi.org/10.1002/eji.200939790
- BenMohamed L, Wechsler SL, Nesburn AB. Lipopeptide vaccines–yesterday, today, and tomorrow. Lancet Infect Dis 2002; 2:425-31; PMID:12127354; http://dx.doi.org/10.1016/S1473-3099(02)00318-3
- Sica A, Bronte V. Altered macrophage differentiation and immune dysfunction in tumor development. J Clin Invest 2007; 117:1155-66; PMID:17476345; http://dx.doi.org/10.1172/JCI31422
- Mantovani A, Sozzani S, Locati M, Allavena P, Sica A. Macrophage polarization: tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol 2002; 23:549-55; PMID:12401408; http://dx.doi.org/10.1016/S1471-4906(02)02302-5
- Hao NB, Lü MH, Fan YH, Cao YL, Zhang ZR, Yang SM. Macrophages in tumor microenvironments and the progression of tumors. Clin Dev Immunol 2012; 2012:948098; PMID:22778768; http://dx.doi.org/10.1155/2012/948098
- Bergot AS, Kassianos A, Frazer IH, Mittal D. New Approaches to Immunotherapy for HPV Associated Cancers. Cancers (Basel) 2011; 3:3461-95; PMID:24212964; http://dx.doi.org/10.3390/cancers3033461
- Guzmán-Olea E, Bermúdez-Morales VH, Peralta-Zaragoza O, Torres-Poveda K, Madrid-Marina V. Molecular Mechanism and Potential Targets for Blocking HPV-Induced Lesion Development. J Oncol 2012; 2012:278312; PMID:22220169; http://dx.doi.org/10.1155/2012/278312
- Lepique AP, Daghastanli KR, Cuccovia IM, Villa LL. HPV16 tumor associated macrophages suppress antitumor T cell responses. Clin Cancer Res 2009; 15:4391-400; PMID:19549768; http://dx.doi.org/10.1158/1078-0432.CCR-09-0489
- Allavena P, Mantovani A. Immunology in the clinic review series; focus on cancer: tumour-associated macrophages: undisputed stars of the inflammatory tumour microenvironment. Clin Exp Immunol 2012; 167:195-205; PMID:22235995; http://dx.doi.org/10.1111/j.1365-2249.2011.04515.x
- Predina J, Eruslanov E, Judy B, Kapoor V, Cheng G, Wang LC, Sun J, Moon EK, Fridlender ZG, Albelda S, et al. Changes in the local tumor microenvironment in recurrent cancers may explain the failure of vaccines after surgery. Proc Natl Acad Sci U S A 2013; 110:E415-24; PMID:23271806; http://dx.doi.org/10.1073/pnas.1211850110
- Sha W, Brüne B, Weigert A. The multi-faceted roles of prostaglandin E2 in cancer-infiltrating mononuclear phagocyte biology. Immunobiology 2012; 217:1225-32; PMID:22727331; http://dx.doi.org/10.1016/j.imbio.2012.05.001
- Buwitt-Beckmann U, Heine H, Wiesmüller KH, Jung G, Brock R, Akira S, Ulmer AJ. Toll-like receptor 6-independent signaling by diacylated lipopeptides. Eur J Immunol 2005; 35:282-9; PMID:15580661; http://dx.doi.org/10.1002/eji.200424955
- Wu W, Li R, Malladi SS, Warshakoon HJ, Kimbrell MR, Amolins MW, Ukani R, Datta A, David SA. Structure-activity relationships in toll-like receptor-2 agonistic diacylthioglycerol lipopeptides. J Med Chem 2010; 53:3198-213; PMID:20302301; http://dx.doi.org/10.1021/jm901839g
- Heusinkveld M, van der Burg SH. Identification and manipulation of tumor associated macrophages in human cancers. J Transl Med 2011; 9:216; PMID:22176642; http://dx.doi.org/10.1186/1479-5876-9-216
- Chang LS, Leng CH, Yeh YC, Wu CC, Chen HW, Huang HM, Liu SJ. Toll-like receptor 9 agonist enhances anti-tumor immunity and inhibits tumor-associated immunosuppressive cells numbers in a mouse cervical cancer model following recombinant lipoprotein therapy. Mol Cancer 2014; 13:60; PMID:24642245; http://dx.doi.org/10.1186/1476-4598-13-60
- Huang CY, Chen JJ, Shen KY, Chang LS, Yeh YC, Chen IH, Chong P, Liu SJ, Leng CH. Recombinant lipidated HPV E7 induces a Th-1-biased immune response and protective immunity against cervical cancer in a mouse model. PLoS One 2012; 7:e40970; PMID:22815882; http://dx.doi.org/10.1371/journal.pone.0040970
- Chua BY, Eriksson EM, Brown LE, Zeng W, Gowans EJ, Torresi J, Jackson DC. A self-adjuvanting lipopeptide-based vaccine candidate for the treatment of hepatitis C virus infection. Vaccine 2008; 26:4866-75; PMID:18455278; http://dx.doi.org/10.1016/j.vaccine.2008.03.032
- Zanin-Zhorov A, Cahalon L, Tal G, Margalit R, Lider O, Cohen IR. Heat shock protein 60 enhances CD4+ CD25+ regulatory T cell function via innate TLR2 signaling. J Clin Invest 2006; 116:2022-32; PMID:16767222; http://dx.doi.org/10.1172/JCI28423
- Dang Y, Wagner WM, Gad E, Rastetter L, Berger CM, Holt GE, Disis ML. Dendritic cell-activating vaccine adjuvants differ in the ability to elicit antitumor immunity due to an adjuvant-specific induction of immunosuppressive cells. Clin Cancer Res 2012; 18:3122-31; PMID:22510348; http://dx.doi.org/10.1158/1078-0432.CCR-12-0113
- Sato T, Terai M, Tamura Y, Alexeev V, Mastrangelo MJ, Selvan SR. Interleukin 10 in the tumor microenvironment: a target for anticancer immunotherapy. Immunol Res 2011; 51:170-82; PMID:22139852; http://dx.doi.org/10.1007/s12026-011-8262-6
- Bolpetti A, Silva JS, Villa LL, Lepique AP. Interleukin-10 production by tumor infiltrating macrophages plays a role in Human Papillomavirus 16 tumor growth. BMC Immunol 2010; 11:27; PMID:20525400; http://dx.doi.org/10.1186/1471-2172-11-27
- Mahic M, Yaqub S, Johansson CC, Taskén K, Aandahl EM. FOXP3+CD4+CD25+ adaptive regulatory T cells express cyclooxygenase-2 and suppress effector T cells by a prostaglandin E2-dependent mechanism. J Immunol 2006; 177:246-54; PMID:16785520; http://dx.doi.org/10.4049/jimmunol.177.1.246
- Eberstål S, Sandén E, Fritzell S, Darabi A, Visse E, Siesjö P. Intratumoral COX-2 inhibition enhances GM-CSF immunotherapy against established mouse GL261 brain tumors. Int J Cancer 2014; 134:2748-53; PMID:24243648; http://dx.doi.org/10.1002/ijc.28607
- Demasi M, Cleland LG, Cook-Johnson RJ, Caughey GE, James MJ. Effects of hypoxia on monocyte inflammatory mediator production: Dissociation between changes in cyclooxygenase-2 expression and eicosanoid synthesis. J Biol Chem 2003; 278:38607-16; PMID:12874281; http://dx.doi.org/10.1074/jbc.M305944200
- Aichele P, Zinke J, Grode L, Schwendener RA, Kaufmann SH, Seiler P. Macrophages of the splenic marginal zone are essential for trapping of blood-borne particulate antigen but dispensable for induction of specific T cell responses. J Immunol 2003; 171:1148-55; PMID:12874200; http://dx.doi.org/10.4049/jimmunol.171.3.1148
- Le Borgne M, Etchart N, Goubier A, Lira SA, Sirard JC, van Rooijen N, Caux C, Aït-Yahia S, Vicari A, Kaiserlian D, et al. Dendritic cells rapidly recruited into epithelial tissues via CCR6/CCL20 are responsible for CD8+ T cell crosspriming in vivo. Immunity 2006; 24:191-201; PMID:16473831; http://dx.doi.org/10.1016/j.immuni.2006.01.005
- Guth AM, Hafeman SD, Dow SW. Depletion of phagocytic myeloid cells triggers spontaneous T cell- and NK cell-dependent antitumor activity. Oncoimmunology 2012; 1:1248-57; PMID:23243588; http://dx.doi.org/10.4161/onci.21317
- Chen HW, Leng CH, Liu HY, Cheng WF, Chang YW, Wu PY, Lien SP, Huang TY, Chiang SK, Lin MH, et al. Identification of HLA-A11-restricted CTL epitopes derived from HPV type 18 using DNA immunization. Cancer Biol Ther 2009; 8:2025-32; PMID:19738415; http://dx.doi.org/10.4161/cbt.8.21.9732
- Chan PK, Liu SJ, Cheung TH, Yeo W, Ngai SM, Cheung JL, Chong P, Man S. T-cell response to human papillomavirus type 58 L1, E6, And E7 peptides in women with cleared infection, cervical intraepithelial neoplasia, or invasive cancer. Clin Vaccine Immunol 2010; 17:1315-21; PMID:20668141; http://dx.doi.org/10.1128/CVI.00105-10
- Chan PK, Liu SJ, Cheung JL, Cheung TH, Yeo W, Chong P, Man S. T-cell response to human papillomavirus type 52 L1, E6, and E7 peptides in women with transient infection, cervical intraepithelial neoplasia, and invasive cancer. J Med Virol 2011; 83:1023-30; PMID:21503915; http://dx.doi.org/10.1002/jmv.21889
- Chen HW, Liu SJ, Liu HH, Kwok Y, Lin CL, Lin LH, Chen MY, Tsai JP, Chang LS, Chiu FF, et al. A novel technology for the production of a heterologous lipoprotein immunogen in high yield has implications for the field of vaccine design. Vaccine 2009; 27:1400-9; PMID:19150476; http://dx.doi.org/10.1016/j.vaccine.2008.12.043