
Abstract
A decrease in the expression of Islet-1 (Isl-1), an islet transcription factor, has been reported in several physiological settings of reduced β-cell function. Here, we investigate whether an increased level of Isl-1 in islet cells can enhance β-cell function and/or mass. We demonstrate that transgenic mice with Isl-1 overexpression display improved glucose tolerance and enhanced insulin secretion without significant changes in β cell mass. From our microarray study, we identify approximately 135 differentially expressed genes in the islets of Isl-1 overexpressing mice that have been implicated to function in numerous biological processes including protein trafficking, metabolism and differentiation. Using real-time PCR we have confirmed upregulation of Caps2, Sec14l4, Slc2a10, P2rx7, Afamin, and Neurogenin 3 that may in part mediate the observed improved insulin secretion in Isl-1 overexpressing mice. These findings show for the first time that Isl-1 is a key factor in regulating adult β cell function in vivo, and suggest that Isl-1 elevation could be beneficial to improve glucose homeostasis.
Introduction
Diabetes mellitus is a multi-organ disease that results from the loss of β-cell function in the pancreas and insulin resistance in peripheral tissues. To understand the pathophysiology of diabetes, gene expression analyses of multiple tissues, including pancreatic islets, have been performed in different physiological conditions of altered β-cell function.Citation1,Citation2 By analyzing changes in gene expression under different conditions such as obesity, strain and age, Keller and colleagues have detected a decrease of Islet-1 (Isl-1) expression in the islets of obese diabetic mice.Citation1 Similarly, aging can induce a distinct gene expression program in mouse islets, leading to a decrease in the expression of a small number of genes, including Isl-1.Citation2-Citation5 Taken together, these findings suggest that a decrease in Isl-1 expression might contribute to the pathophysiology of the disease and an increase in Isl-1 expression might be beneficial for a β-cell to maintain its full functionality.
Isl-1, a LIM-homeodomain transcription factor and a known Insulin gene enhancer binding protein, is expressed in the developing pancreas and adult islet cells.Citation6-Citation9 Using genetic approaches in mice, Isl-1 has been shown to play an important role during early pancreatic development as well as during maturation of the islet cells at the secondary transition.Citation6,Citation10 Because most of the in vivo analyses for Isl-1 function in the pancreas reported thus far have been focused on studies performed using loss-of function mouse models,Citation6,Citation10,Citation11 we set out to test whether increasing Isl-1 expression and thus activity in islets would impact adult β-cell function and mass using transgenic overexpression in mice. Our findings suggest that a modest increase of Isl-1 expression in islets is sufficient to enhance β-cell function, but not adequate to increase β-cell mass in vivo.
Results
Increased Isl-1 expression leads to improved β-cell function
To investigate the role of Isl-1 in adult islets and to test the hypothesis that β-cell function can be enhanced upon Isl-1 overexpression, a series of functional tests were performed in control and Isl-1 overexpressing (Pdx1PB-Isl-1-Myc) mice.Citation11 In this model, a Myc-tagged rat Isl-1 gene was cloned downstream of the Pdx1PB fragment which has been shown to drive islet-specific expression of Pdx1 and is active as early as E11.5.Citation12,Citation13 Despite the known early transgenic activity of the Pdx1PB fragment in the developing islet cells,Citation12,Citation13 we only detected a few β-cells expressing the transgene at E17.5 (). It was not until adulthood that we observed a significant expression of the transgene in Pdx1PB-Isl-1-Myc mice demonstrated by Myc immunostaining with substantial islet specific expression of Isl-1-Myc proteins (). We detected approximately one-third of β-cells in 2-mo-old Pdx1PB-Isl-1-Myc mice with the Isl-1-Myc protein ().Citation11 We previously reported a 2-fold increase in total Isl-1 protein and mRNA, as detected by western blot and real-time PCR, respectively.Citation11
Figure 1. Physiological increase of Isl-1 expression is sufficient to enhance β-cell function in adult mice. (A-D) Immunohistochemical analysis for insulin and Myc expression in E17.5 and 2-mo-old control and Pdx1PB-Isl-1-Myc mice. (E) Random Fed blood glucose levels (n > 5). (F) Random Fed blood insulin levels (n > 8). (G) Fasted blood glucagon levels. (H-I) Total pancreatic insulin and glucagon content. (J) Body weight of 2-mo-old control and Pdx1PB-Isl-1-Myc mice. (K) Glucose tolerance test demonstrates Pdx1PB-Isl-1-Myc mice exhibit improved glucose clearance (n > 10). (L) Measurements of plasma insulin levels during glucose tolerance test show increased insulin secretion in Pdx1PB-Isl-1-Myc mice. (M) Static incubation analysis demonstrates enhanced insulin secretion in islets of Pdx1PB-Isl-1-Myc mice challenged with 16mM glucose. (N) Body weight of 9-mo-old control and Pdx1PB-Isl-1-Myc mice. (O) Glucose tolerance test demonstrates Pdx1PB-Isl-1-Myc mice exhibit modest improvement of glucose tolerance (n > 6). (P) Measurements of plasma insulin levels during glucose tolerance test show increased insulin secretion in 9-mo-old Pdx1PB-Isl-1-Myc mice. (Q) Static incubation analysis demonstrates enhanced insulin secretion in islets of 9-mo-old Pdx1PB-Isl-1-Myc mice challenged with 16mM glucose and 0.2 μM glyburide (n > 3 animals/group). Data represents the mean ± SEM * P-value < 0.05.
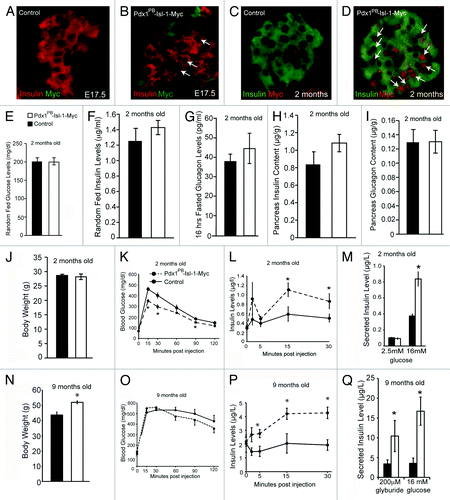
From our examination of the pancreatic sections, we found no obvious changes in the overall appearance of the pancreatic islets in Pdx1PB-Isl-1-Myc mice when compared with control littermates.Citation11 We also did not detect changes in circulating glucose, insulin or glucagon levels in two-month-old Pdx1PB-Isl-1-Myc mice (). However, a significant improvement in clearing glucose was observed in Pdx1PB-Isl-1-Myc mice during glucose tolerance tests (), which was attributed to an enhanced insulin secretory response (). This enhancement in insulin release was also observed in our in vitro static incubation studies in which isolated islets from control and Pdx1PB-Isl-1-Myc mice were treated with low and high concentration of glucose (). As animals aged, 9-mo-old Pdx1PB-Isl-1-Myc mice continued to show improved insulin secretion; however, despite this increase, we only detected a modest improvement in glucose tolerance tests, suggesting chronic high levels of insulin in Pdx1PB-Isl-1-Myc mice likely lead to increased body weight and eventual insulin insensitivity (). To further investigate if other insulin secretagogues can also enhance insulin release in Pdx1PB-Isl-1-Myc mice, isolated islets from both groups were treated for 15 min with 200 μM glyburide, a sulfonylurea that blocks KATP channels leading to a fully depolarized plasma membrane, resulting in insulin secretion.Citation14,Citation15 Compared with the control islets, glyburide-treated islets from Pdx1PB-Isl-1-Myc mice displayed a robust increase in insulin release similar to what was seen in the 16 mM glucose-treated islets (). These data indicate that islets with a higher level of Isl-1 have the ability to release more insulin upon glucose stimulation, which is similar to treatment with high dose of glyburide.
Isl-1 overexpression does not increase β-cell mass
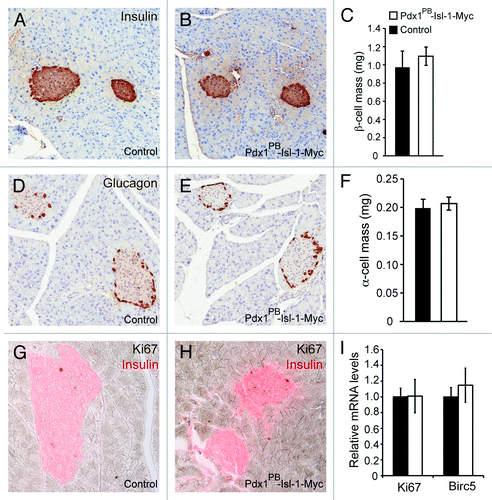
The enhanced response to glyburide observed in the islets of Pdx1PB-Isl-1-Myc mice pointed to a possibility of increased insulin production. Therefore, we examined total pancreatic insulin content of control and Pdx1PB-Isl-1-Myc mice, which surprisingly showed no significant differences (). Total glucagon content was also measured which also showed comparable levels between control and Pdx1PB-Isl-1-Myc mice (). Recently, it has been shown that rat islets, when transduced with lentivirus overexpressing Isl-1, increased cell proliferation, which was the result of an upregulation of c-Myc and CyclinD1.Citation16 Therefore, we tested whether increased Isl-1 expression in vivo affected β- or α-cell mass in the Pdx1PB-Isl-1-Myc mice. Unlike what was reported, we found no significant differences in β-cell or α-cell mass between control and Pdx1PB-Isl-1-Myc mice likely due to the lower level of Isl-1 expression (). To investigate whether cell survival and/or proliferation were affected in the Pdx1PB-Isl-1-Myc mice, we performed immunostaining for Caspase 3 and Ki67, respectively, and found no significant difference (; and data not shown). Real-time PCR analysis of isolated islets for Ki67 and Birc5 expression, markers for proliferating cells,Citation17 further confirmed that there were no significant changes in proliferation in the islets of Pdx1PB-Isl-1-Myc mice (). Taken together, these findings suggest that while a 2-fold increase of Isl-1 is sufficient to enhance β-cell function, this increase is not enough to promote β- or α-cell mass expansion in vivo.
Figure 2. Isl-1 overexpression does not affect β cell mass and cell proliferation in 2-mo-old adult mice. Immunostaining analysis for insulin (A-C) and glucagon (D-F) were performed to measure β- and α-cell mass in control and Pdx1PB-Isl-1-Myc mice. Immunostaining analysis for Ki67 (G-H) shows no significant changes in β-cell proliferation. (I) Real-time PCR analysis shows comparable mRNA levels of Ki67 and Birc5 in islets between control Pdx1PB-Isl-1-Myc mice. Bars represent the mean ± SEM.
Altered gene expression in the islets of Pdx1PB-Isl-1-Myc mice
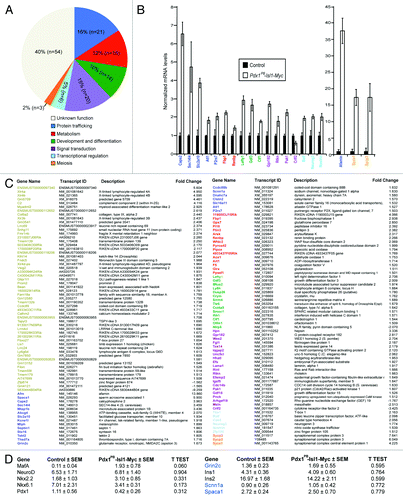
To evaluate the molecular targets that may mediate improved β-cell function in Pdx1PB-Isl-1-Myc mice, we performed microarray analysis on purity-matched islets from two-month-old control and Pdx1PB-Isl-1-Myc mice. We detected 135 differentially expressed genes implicated to function in protein trafficking, metabolism, development, signal transduction, and transcriptional regulation (). To validate results obtained from the microarray study, we chose several candidate genes from each of the six categories and focused specifically on several transcripts that have implicated functions in exocytosis, secretion and transport in other systems (). From our real-time PCR, we detected an increase in the expression of Afamin,Citation23 Caps2,Citation21,Citation22 Slc2a10,Citation26,Citation27 Sec14l4,Citation24,Citation25 and P2rx7Citation18-Citation20 as well as Neurogenin3Citation28 in the Pdx1PB-Isl-1-Myc islets (). In the canonical model of insulin secretion, following a rise in blood glucose, ATP/ADP ratio increases which leads to ATP-sensitive K+ channels closure. This closure triggers membrane depolarization, which in turn increases intracellular Ca2+ concentration leading to insulin granule exocytosis. Interestingly, both Caps2 and P2rx7 have implicated functions in regulating secretion by affecting Ca2+ signaling through binding to Ca2+ (Caps2) and mediating the effect of ATP (P2rx7), respectively.Citation18-Citation22 In contrary, while Afamin and Sec14l4 have no known functions in mediating insulin secretion, they have been implicated in transport of α-tocopherol, a form of vitamin E.Citation23-Citation25 Based on their involvement in transport, these proteins may play crucial roles in regulating exocytosis and vesicular transport in islet β-cells. Additionally, with the implicated functions of Slc2a10 (Glut10), a facilitated glucose transporter and Neurogenin3 in maintaining glucose homeostasis,Citation26-Citation31 our study suggest that increased levels of these genes potentially lead to the enhanced β-cell function seen in the Pdx1PB -Isl-1-Myc transgenic mice. Similar to an increase of Arx expression demonstrated previously by real-time PCR,Citation11 an increase in MafA expression was also detected (), although neither of these genes was detected in the microarray study, likely due to sensitivity issues with gene expression arrays.Citation32 We also attempted to confirm an increase in the expression of several other genes that may be involved in secretion, including glutamate receptor (Grin2c),Citation33 non voltage-gated sodium channel (Scnn1a),Citation34,Citation35 and sperm acrosome associated 1 (Spaca1),Citation36,Citation37 but were unable to validate upregulation of these genes. Taken together, our results suggest that Isl-1 may mediate increased β-cell functionality through a subset of genes involved in protein trafficking, metabolism, development, signal transduction, and transcriptional regulation.
Figure 3. Genome-wide gene expression analysis of 2-mo-old control and Pdx1PB-Isl-1-Myc islets. (A) Diagram summarizes functions of the 135 differentially expressed genes from the gene expression analysis. A 10% False Discovery Rate was used in this analysis. (B) Real-time PCR analysis confirmation of a subset of upregulated genes chosen from the microarray gene list. Bars represent the mean ± SEM. All changes were significant with p value < 0.05. (C) List of genes that are differentially expressed in the microarray study. (D) Additional genes evaluated by real-time PCR in islets of control and Pdx1PB-Isl-1-Myc mice.
Discussion
In this study, we investigate whether islet-specific Isl-1 overexpression in vivo can enhance β-cell function and cell proliferation. Although only about one third of β-cells activate the transgene,Citation11 the insulin secretory response in Pdx1PB-Isl-1-Myc mice was significantly improved. Additionally, an increase of Isl-1 expression in the islet α- or δ-cells might also alter glucose homeostasis in Pdx1PB-Isl-1-Myc mice. However, based on the small number of these non-β-cells expressing the Isl-1-Myc transgene,Citation11 we conclude that much of the improved glucose tolerance phenotype is due to Isl-1 overexpression in β cells.
A portion of our phenotype observed may be due to an increase in the expression of MafA, which is known to control genes implicated in insulin secretion.Citation38-Citation40 However, we have also identified ~135 differentially expressed genes from our microarray and were able to confirm an increase in expression in a subset of these genes by real-time PCR in islets of Pdx1PB-Isl-1-Myc mice. These genes are implicated to function in protein trafficking and metabolism, and may play important roles in β-cell biology. In this analysis, we were able to detect an increase in Slc2a10 (Glut10), a gene that has been implicated in glucose metabolism and type 2 diabetes,Citation26,Citation27 increased levels of SEC14L4, a factor that has been implicated to be involved in secretion in organs like salivary gland, prostate and pancreas,Citation24,Citation25 and a 35-fold increase in Afamin levels, a transporter for α-tocopherol in the CNS.Citation23 Additionally, P2rx7, a purinergic receptor with implicated function for regulating both endocrine and exocrine pancreas function and Caps2, a calcium binding protein were also increased.Citation18-Citation22 Although the precise function of these genes and the molecular mechanisms by which they impact β-cell physiology remain to be fully characterized, it is tempting to speculate that increased levels of some of these genes may have a positive impact on exocytosis, granule transport and secretion. Future studies manipulating these genes in either animal models or cell culture might provide further insights into the function of these genes in adult β-cells.
Although the Notch/Neurogenin3 pathway has been shown to play critical roles in the maintenance of the pancreatic progenitor pool and in the differentiation into exocrine and endocrine lineages,Citation29,Citation41,Citation42 it has recently been shown to also play an important role in the regulation of β-cell maturation and function.Citation28 Therefore, a 2-fold increase in Neurogenin3 expression is likely mediating part of the enhanced β-cell function in the Pdx1PB-Isl-1-Myc mice. In addition, Neurogenin3 has also been implicated in regulating adult β-cell survival as a 4-fold increase in Neurogenin3 expression result in apoptosis.Citation30 Similarly, expression of Neurogenin3 in islets is negatively regulated by Musashi, a RNA binding protein with antiapoptotic roles in adult β-cells.Citation30,Citation31 Although it is not clear whether a modest increase in Neurogenin3 levels is physiologically relevant for β-cells, our observation fits with the current model that the adult β-cell is likely sensitive to Neurogenin3 levels and that differing levels of Neurogenin3 in β-cells result in activation of different gene programs (e.g., secretion vs. apoptosis).Citation28,Citation30,Citation31
A large network of islet transcriptional regulators is involved in the regulation of β cell development and function.Citation43 Recently, it has been demonstrated that stress-induced hyperglycemia could be prevented in mice overexpressing Pax4, a β-cell specific gene.Citation44 Furthermore, in contrast to what was shown in cell lines, increased expression of Nkx6.1 in mice, another key β-cell transcriptional regulator, had no impact on β-cell mass or glucose clearance.Citation45,Citation46 Taken together, these findings demonstrate the importance of using gain-of-function studies in vivo to understand the sufficiency of any islet transcription factors.
In summary, microarray analysis provides an ideal approach for comparing changes in gene expression on a large scale. We propose that using an Isl-1 overexpressing transgenic mouse model provides a novel platform for expanding our understanding of how a small increase in transcription factor expression can lead to a significant impact in insulin secretion in adult mice. We have identified a group of genes, many of which have not been fully characterized in islet β-cells, which may contribute significantly to the phenotype we observe. Although we did not find any significant Isl-1 binding within the genomic regions of some of these differentially expressed genes in our Isl-1 ChIP-Seq data set (unpublished data), this observation does not exclude direct binding of Isl-1 to these genes as regulatory domains of many mammalian genes can be far away from the gene itself. Alternatively, these genes might be direct targets of additional transcriptional regulator(s) yet to be identified using PCR-based transcription factor screens or RNA-Seq.Citation32 Furthermore, while Guo and colleagues had reported that Isl-1 can promote cell proliferation and attenuate cell death against oxidative stress in isolated rat islets and HIT-T15 cells,Citation16 we did not detect significant changes in cell proliferation or cell survival in our transgenic mouse model. The discrepancy between these findings might simply due to the high levels of Isl-1 expression achieved in the lentivirus-based in vitro model, which might lead to activation of different gene programs.Citation16
In conclusion, these novel findings provide the islet biology community with the gene expression profile of enhanced β-cell function. Future experiments designed to manipulate these genes in mouse models or cell cultures will lead to better understanding of how insulin secretion is regulated and how glucose homeostasis is maintained in adult animals.
Materials and Methods
Animals
Pdx1PB -Isl-1-Myc male mice used in this study were kept on a C57BL/6J background. Although only one Pdx1PB -Isl-1-Myc transgenic mouse line was characterized in our study, which we acknowledged it as a caveat, we are confident that the effects we observe in the Pdx1PB -Isl-1-Myc mice are due to the elevation of Isl-1 expression as Arx and MafA, two genes that are downregulated in Isl-1 deficient pancreata,Citation10,Citation11 were increased the islets of the Pdx1PB -Isl-1-Myc transgenic mice. Animal experiments were approved by the Children’s Hospital of Philadelphia’s Institutional Animal Care and Use Committee.
Glucose and insulin measurement
Blood glucose levels were measured from tail vein blood using an automatic glucometer (One Touch Ultra; LifeScan). Plasma was separated from whole blood using heparinized tubes (BD Microtainer, BD, 365958) and total pancreatic proteins were extracted with acid-ethanol. Insulin protein levels were measured using ELISA kit (Mercodia, 10-1113-01).
Immunohistochemistry and immunofluorescence
Pancreata from control and mutant mice (n > 3/group) were removed, weighed, fixed in 4% PFA and embedded in paraffin. For α- and β-cell mass measurements, slides with the maximum footprint (three slides per animal) were subjected to immunohistochemical staining for glucagon or insulin. Hormone positive areas were measured using Aperio software. Cell mass was obtained by hormone-positive area percentage times the weight of the pancreas. The following primary antibodies were used: insulin (1:1000; Linco, 1013), glucagon (1:3000; Linco, Cat. No. 4030-01F) and Ki 67 (1:1000; Leica, NCL-Ki67p). Cy3-conjugated secondary antibody (1:600) was from Jackson ImmunoResearch.
Glucose tolerance test
Following a 16 h fast, animals (n = 10/group) were injected with glucose (2 mg/g body weight) in sterile PBS. Blood glucose levels were monitored at 0, 15, 30, 60, 90 and 120 min after injection.
Islet static incubation
Islets were isolated using the standard collagenase procedure.Citation47 About 50 islets from each animal were cultured with islet medium (RPMI 1640, 10%FBS, 1x penicillin/streptomycin, 1 x glutamine, 5 mM Hepes, 5 mM Glucose). After overnight incubation at 5% CO2, 37°C, the islets were stimulated with low glucose (2.5 mM), then high glucose (16 mM) for 30 min sequentially. Mouse Insulin ELISA was performed to measure insulin levels. Four animals from each group were used for this assay. All values were normalized by total islet insulin content.
Microarray and quantitative real-time PCR
Total RNA from isolated islets was extracted in TRIzol (Invitrogen, 15596-026) and then purified using RNeasy Mini Kit (Qiagen, 74104) according to the manufacturer’s instructions. Four pairs of control and transgenic islets were purity matched,Citation48 and hybridized to Agilient Gene expression Array for dual color expression analysis by the DRC Functional Genomics Core of the Penn Institute for Diabetes, Obesity and Metabolism. For real-time PCR, approximately 1 μg RNA was reverse transcribed into cDNA. Real-time PCR reactions were performed using the Brilliant SYBR Green PCR Master Mix (Stratagene, 600828) on Mx3005 Multiplex Quantitative PCR System (Stratagene). All reactions were performed in triplicate with reference dye normalization and medium values were used for analysis. Primer sequences are available upon request.
Acknowledgments
We thank the members of the Molecular Pathology and Imaging Core in the Center for Molecular Studies in Digestive and Liver Diseases (P30-DK050306) for sample processing and Dr Jonathan Schug and the members of the DRC Functional Genomics and Radioimmunoassay/Biomarkers Cores of the Penn Institute for Diabetes, Obesity and Metabolism (DERC: P30-DK19525) for conducting microarray and RIA experiments. We are grateful to Dr Klaus Kaestner and Dr Christine Reid for the critical reading of the manuscript and Dr Changhong Li for the advice on static incubation studies. C.L.M. was supported by NIH-DK078606 and JDRF-2-2007-730.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
References
- Keller MP, Choi Y, Wang P, Davis DB, Rabaglia ME, Oler AT, et al. A gene expression network model of type 2 diabetes links cell cycle regulation in islets with diabetes susceptibility. Genome Res 2008; 18:706 - 16; http://dx.doi.org/10.1101/gr.074914.107; PMID: 18347327
- Rankin MM, Kushner JA. Aging induces a distinct gene expression program in mouse islets. Islets 2010; 2:345 - 52; http://dx.doi.org/10.4161/isl.2.6.13376; PMID: 21099336
- Rankin MM, Kushner JA. Adaptive beta-cell proliferation is severely restricted with advanced age. Diabetes 2009; 58:1365 - 72; http://dx.doi.org/10.2337/db08-1198; PMID: 19265026
- Teta M, Long SY, Wartschow LM, Rankin MM, Kushner JA. Very slow turnover of beta-cells in aged adult mice. Diabetes 2005; 54:2557 - 67; http://dx.doi.org/10.2337/diabetes.54.9.2557; PMID: 16123343
- Tschen SI, Dhawan S, Gurlo T, Bhushan A. Age-dependent decline in beta-cell proliferation restricts the capacity of beta-cell regeneration in mice. Diabetes 2009; 58:1312 - 20; http://dx.doi.org/10.2337/db08-1651; PMID: 19228811
- Ahlgren U, Pfaff SL, Jessell TM, Edlund T, Edlund H. Independent requirement for ISL1 in formation of pancreatic mesenchyme and islet cells. Nature 1997; 385:257 - 60; http://dx.doi.org/10.1038/385257a0; PMID: 9000074
- May CL. The role of Islet-1 in the endocrine pancreas: Lessons from pancreas specific Islet-1 deficient mice. Islets 2010; 2:121 - 3; http://dx.doi.org/10.4161/isl.2.2.10908; PMID: 21099304
- Thor S, Ericson J, Brännström T, Edlund T. The homeodomain LIM protein Isl-1 is expressed in subsets of neurons and endocrine cells in the adult rat. Neuron 1991; 7:881 - 9; http://dx.doi.org/10.1016/0896-6273(91)90334-V; PMID: 1764243
- Wang M, Drucker DJ. The LIM domain homeobox gene isl-1 is a positive regulator of islet cell-specific proglucagon gene transcription. J Biol Chem 1995; 270:12646 - 52; http://dx.doi.org/10.1074/jbc.270.21.12646; PMID: 7759514
- Du A, Hunter CS, Murray J, Noble D, Cai CL, Evans SM, et al. Islet-1 is required for the maturation, proliferation, and survival of the endocrine pancreas. Diabetes 2009; 58:2059 - 69; http://dx.doi.org/10.2337/db08-0987; PMID: 19502415
- Liu J, Hunter CS, Du A, Ediger B, Walp E, Murray J, et al. Islet-1 regulates Arx transcription during pancreatic islet alpha-cell development. J Biol Chem 2011; 286:15352 - 60; http://dx.doi.org/10.1074/jbc.M111.231670; PMID: 21388963
- Gannon M, Ray MK, Van Zee K, Rausa F, Costa RH, Wright CV. Persistent expression of HNF6 in islet endocrine cells causes disrupted islet architecture and loss of beta cell function. Development 2000; 127:2883 - 95; PMID: 10851133
- Wu KL, Gannon M, Peshavaria M, Offield MF, Henderson E, Ray M, et al. Hepatocyte nuclear factor 3beta is involved in pancreatic beta-cell-specific transcription of the pdx-1 gene. Mol Cell Biol 1997; 17:6002 - 13; PMID: 9315659
- Cherksey B, Altszuler N. Tolbutamide and glyburide differ in effectiveness to displace alpha- and beta-adrenergic radioligands in pancreatic islet cells and membranes. Diabetes 1984; 33:499 - 503; http://dx.doi.org/10.2337/diabetes.33.5.499; PMID: 6327436
- Gorus FK, Schuit FC, In’t Veld PA, Gepts W, Pipeleers DG. Interaction of sulfonylureas with pancreatic beta-cells. A study with glyburide. Diabetes 1988; 37:1090 - 5; http://dx.doi.org/10.2337/diabetes.37.8.1090; PMID: 2839387
- Guo T, Wang W, Zhang H, Liu Y, Chen P, Ma K, et al. ISL1 promotes pancreatic islet cell proliferation. PLoS One 2011; 6:e22387; http://dx.doi.org/10.1371/journal.pone.0022387; PMID: 21829621
- Rieck S, White P, Schug J, Fox AJ, Smirnova O, Gao N, et al. The transcriptional response of the islet to pregnancy in mice. Mol Endocrinol 2009; 23:1702 - 12; http://dx.doi.org/10.1210/me.2009-0144; PMID: 19574445
- Ohtani M, Ohura K, Oka T. Involvement of P2X receptors in the regulation of insulin secretion, proliferation and survival in mouse pancreatic β-cells. Cell Physiol Biochem 2011; 28:355 - 66; http://dx.doi.org/10.1159/000331752; PMID: 21865744
- Amisten S, Meidute-Abaraviciene S, Tan C, Olde B, Lundquist I, Salehi A, et al. ADP mediates inhibition of insulin secretion by activation of P2Y13 receptors in mice. Diabetologia 2010; 53:1927 - 34; http://dx.doi.org/10.1007/s00125-010-1807-8; PMID: 20526761
- Novak I. Purinergic receptors in the endocrine and exocrine pancreas. Purinergic Signal 2008; 4:237 - 53; http://dx.doi.org/10.1007/s11302-007-9087-6; PMID: 18368520
- Yuasa HJ, Nakatomi A, Suzuki T, Yazawa M. Genomic structure of the sponge, Halichondria okadai calcyphosine gene. Gene 2002; 298:21 - 7; http://dx.doi.org/10.1016/S0378-1119(02)00920-4; PMID: 12406572
- Wang S, Chen JZ, Zhang Z, Huang Q, Gu S, Ying K, et al. Cloning, characterization, and expression of calcyphosine 2, a novel human gene encoding an EF-hand Ca(2+)-binding protein. Biochem Biophys Res Commun 2002; 291:414 - 20; http://dx.doi.org/10.1006/bbrc.2002.6461; PMID: 11846421
- Kratzer I, Bernhart E, Wintersperger A, Hammer A, Waltl S, Malle E, et al. Afamin is synthesized by cerebrovascular endothelial cells and mediates alpha-tocopherol transport across an in vitro model of the blood-brain barrier. J Neurochem 2009; 108:707 - 18; http://dx.doi.org/10.1111/j.1471-4159.2008.05796.x; PMID: 19046407
- Kempna P, Ricciarelli R, Azzi A, Zingg JM. Alternative splicing and gene polymorphism of the human TAP3/SEC14L4 gene. Mol Biol Rep 2010; 37:3503 - 8; http://dx.doi.org/10.1007/s11033-009-9943-2; PMID: 20012369
- Zingg JM, Kempna P, Paris M, Reiter E, Villacorta L, Cipollone R, et al. Characterization of three human sec14p-like proteins: alpha-tocopherol transport activity and expression pattern in tissues. Biochimie 2008; 90:1703 - 15; http://dx.doi.org/10.1016/j.biochi.2008.07.008; PMID: 18725265
- McVie-Wylie AJ, Lamson DR, Chen YT. Molecular cloning of a novel member of the GLUT family of transporters, SLC2a10 (GLUT10), localized on chromosome 20q13.1: a candidate gene for NIDDM susceptibility. Genomics 2001; 72:113 - 7; http://dx.doi.org/10.1006/geno.2000.6457; PMID: 11247674
- Dawson PA, Mychaleckyj JC, Fossey SC, Mihic SJ, Craddock AL, Bowden DW. Sequence and functional analysis of GLUT10: a glucose transporter in the Type 2 diabetes-linked region of chromosome 20q12-13.1. Mol Genet Metab 2001; 74:186 - 99; http://dx.doi.org/10.1006/mgme.2001.3212; PMID: 11592815
- Wang S, Jensen JN, Seymour PA, Hsu W, Dor Y, Sander M, et al. Sustained Neurog3 expression in hormone-expressing islet cells is required for endocrine maturation and function. Proc Natl Acad Sci U S A 2009; 106:9715 - 20; http://dx.doi.org/10.1073/pnas.0904247106; PMID: 19487660
- Gradwohl G, Dierich A, LeMeur M, Guillemot F. neurogenin3 is required for the development of the four endocrine cell lineages of the pancreas. Proc Natl Acad Sci U S A 2000; 97:1607 - 11; http://dx.doi.org/10.1073/pnas.97.4.1607; PMID: 10677506
- Dror V, Nguyen V, Walia P, Kalynyak TB, Hill JA, Johnson JD. Notch signalling suppresses apoptosis in adult human and mouse pancreatic islet cells. Diabetologia 2007; 50:2504 - 15; http://dx.doi.org/10.1007/s00125-007-0835-5; PMID: 17922104
- Szabat M, Kalynyak TB, Lim GE, Chu KY, Yang YH, Asadi A, et al. Musashi expression in β-cells coordinates insulin expression, apoptosis and proliferation in response to endoplasmic reticulum stress in diabetes. Cell Death Dis 2011; 2:e232; http://dx.doi.org/10.1038/cddis.2011.119; PMID: 22113197
- Gupta RK, Rosen ED, Spiegelman BM. . Identifying novel transcriptional components controlling energy metabolism. Cell Metab 2011; 14:739 - 45; http://dx.doi.org/10.1016/j.cmet.2011.11.007
- Gammelsaeter R, Coppola T, Marcaggi P, Storm-Mathisen J, Chaudhry FA, Attwell D, et al. A role for glutamate transporters in the regulation of insulin secretion. PLoS One 2011; 6:e22960; http://dx.doi.org/10.1371/journal.pone.0022960; PMID: 21853059
- Ernst SJ, Aguilar-Bryan L, Noebels JL. Sodium channel beta1 regulatory subunit deficiency reduces pancreatic islet glucose-stimulated insulin and glucagon secretion. Endocrinology 2009; 150:1132 - 9; http://dx.doi.org/10.1210/en.2008-0991; PMID: 18988673
- Swayne LA, Mezghrani A, Lory P, Nargeot J, Monteil A. The NALCN ion channel is a new actor in pancreatic β-cell physiology. Islets 2010; 2:54 - 6; http://dx.doi.org/10.4161/isl.2.1.10522; PMID: 21099296
- Mandal A, Klotz KL, Shetty J, Jayes FL, Wolkowicz MJ, Bolling LC, et al. SLLP1, a unique, intra-acrosomal, non-bacteriolytic, c lysozyme-like protein of human spermatozoa. Biol Reprod 2003; 68:1525 - 37; http://dx.doi.org/10.1095/biolreprod.102.010108; PMID: 12606493
- Abou-Haila A, Tulsiani DR. Mammalian sperm acrosome: formation, contents, and function. Arch Biochem Biophys 2000; 379:173 - 82; http://dx.doi.org/10.1006/abbi.2000.1880; PMID: 10898932
- Matsuoka TA, Zhao L, Artner I, Jarrett HW, Friedman D, Means A, et al. Members of the large Maf transcription family regulate insulin gene transcription in islet beta cells. Mol Cell Biol 2003; 23:6049 - 62; http://dx.doi.org/10.1128/MCB.23.17.6049-6062.2003; PMID: 12917329
- Wang H, Brun T, Kataoka K, Sharma AJ, Wollheim CB. MAFA controls genes implicated in insulin biosynthesis and secretion. Diabetologia 2007; 50:348 - 58; http://dx.doi.org/10.1007/s00125-006-0490-2; PMID: 17149590
- Zhang C, Moriguchi T, Kajihara M, Esaki R, Harada A, Shimohata H, et al. MafA is a key regulator of glucose-stimulated insulin secretion. Mol Cell Biol 2005; 25:4969 - 76; http://dx.doi.org/10.1128/MCB.25.12.4969-4976.2005; PMID: 15923615
- Apelqvist A, Li H, Sommer L, Beatus P, Anderson DJ, Honjo T, et al. Notch signalling controls pancreatic cell differentiation. Nature 1999; 400:877 - 81; http://dx.doi.org/10.1038/23716; PMID: 10476967
- Schwitzgebel VM, Scheel DW, Conners JR, Kalamaras J, Lee JE, Anderson DJ, et al. Expression of neurogenin3 reveals an islet cell precursor population in the pancreas. Development 2000; 127:3533 - 42; PMID: 10903178
- Oliver-Krasinski JM, Stoffers DA. On the origin of the beta cell. Genes Dev 2008; 22:1998 - 2021; http://dx.doi.org/10.1101/gad.1670808; PMID: 18676806
- Hu He KH, Lorenzo PI, Brun T, Jimenez Moreno CM, Aeberhard D, Vallejo Ortega J, et al. In vivo conditional Pax4 overexpression in mature islet β-cells prevents stress-induced hyperglycemia in mice. Diabetes 2011; 60:1705 - 15; http://dx.doi.org/10.2337/db10-1102; PMID: 21521872
- Schaffer AE, Yang AJ, Thorel F, Herrera PL, Sander M. Transgenic overexpression of the transcription factor Nkx6.1 in β-cells of mice does not increase β-cell proliferation, β-cell mass, or improve glucose clearance. Mol Endocrinol 2011; 25:1904 - 14; http://dx.doi.org/10.1210/me.2011-1010; PMID: 21964593
- Schisler JC, Fueger PT, Babu DA, Hohmeier HE, Tessem JS, Lu D, et al. Stimulation of human and rat islet beta-cell proliferation with retention of function by the homeodomain transcription factor Nkx6.1. Mol Cell Biol 2008; 28:3465 - 76; http://dx.doi.org/10.1128/MCB.01791-07; PMID: 18347054
- Gupta RK, Vatamaniuk MZ, Lee CS, Flaschen RC, Fulmer JT, Matschinsky FM, et al. The MODY1 gene HNF-4alpha regulates selected genes involved in insulin secretion. J Clin Invest 2005; 115:1006 - 15; PMID: 15761495
- Lantz KA, Vatamaniuk MZ, Brestelli JE, Friedman JR, Matschinsky FM, Kaestner KH. Foxa2 regulates multiple pathways of insulin secretion. J Clin Invest 2004; 114:512 - 20; PMID: 15314688

