
Abstract
Diabetes mellitus (DM) is a great public health problem, which attacks part of the world population, being characterized by an imbalance in body glucose homeostasis. Physical exercise is pointed as a protective agent and is also recommended to people with DM. As pancreatic islets present an important role in glucose homeostasis, we aim to study the role of physical exercise (chronic adaptations and acute responses) in pancreatic islets functionality in Wistar male rats. First, animals were divided into two groups: sedentary (S) and aerobic trained (T). At the end of 8 weeks, half of them (S and T) were submitted to an acute exercise session (exercise until exhaustion), being subdivided as acute sedentary (AS) and acute trained (AT). After the experimental period, periepididymal, retroperitoneal and subcutaneous fat pads, blood, soleus muscle and pancreatic islets were collected and prepared for further analysis. From the pancreatic islets, total insulin content, insulin secretion stimulated by glucose, leucine, arginine and carbachol were analyzed. Our results pointed that body adiposity and glucose homeostasis improved with chronic physical exercise. In addition, total insulin content was reduced in group AT, insulin secretion stimulated by glucose was reduced in trained groups (T and AT) and insulin secretion stimulated by carbachol was increased in group AT. There were no significant differences in insulin secretion stimulated by arginine and leucine. We identified a possible modulating action on insulin secretion, probably related to the association of chronic adaptation with an acute response on cholinergic activity in pancreatic islets.
Introduction
Diabetes mellitus is a disease characterized by chronically above normal values of glycemia. Diabetes mellitus can be caused by autoimmune destruction of insulin-producing and secreting cells, the β-pancreatic cells, and it is called type I diabetes mellitus (DM1). It also can be named type II diabetes mellitus (DM2), which is characterized by reduced insulin action, called insulin resistance, compensatory hyperinsulinemia, overload of β-pancreatic cells, followed by increased β-cells apoptosis.Citation1-Citation5
Type 2 diabetes mellitus is a disease that attacks a major part of the world population. Data from the World Health Organization (WHO) indicate that about 10% of the Brazilian population can be characterized as diabetic. In addition, it is estimated that currently about 250 million people can be considered diabetic, and by 2025 this number may increase to around 380 million people.Citation6
The incidence of DM2 presents an inverse relationship with physical fitness levels. Shaibi et al.Citation7 observed that the cardiorespiratory fitness and daily physical activity levels are reduced in young people with DM2. In addition, Krause et al.Citation8 also observed an inverse relationship between cardiorespiratory fitness and physical exercise with the prevalence of DM2 in women. Grundy et al.Citation9 performed an evaluation of cardiorespiratory fitness and the presence of metabolic diseases in about 60,000 men and 22,000 women, and concluded that the reduction of cardiorespiratory fitness is related to higher instances of obesity, dyslipidemia and DM2.
Due to the relationship between physical exercise and the improvement in glycemic homeostasis, some researchers sought to delineate a relationship between physical exercise and insulin secretion by pancreatic cells. Therefore, an improvement in insulin secretion capacity was observed in isolated pancreatic islets, stimulated with high glucose concentration, from rats submitted to swimming training for 8 weeksCitation10 and in the secretion stimulated by lower glucose concentration in type I diabetic rats.Citation11 It was also identified that 8 weeks of treadmill running activated growth pathways and cell survival in pancreatic islets,Citation12 increased anaplerotic enzyme activityCitation13 and the mediator agent responsible for higher insulin secretory capacity induced by exercise training would be interleucine 6 (IL-6).Citation14
However, most studies indicate adaptations resulting from chronic physical exercise and the stimulation of insulin secretion induced only by its main secretagogue, glucose. This way, our study sought to establish the relationship between chronic (long-term) and acute (short-term) actions of physical exercise on the secretory insulin capacity of isolated pancreatic islets stimulated by different secretagogues.
Results
Training efficacy
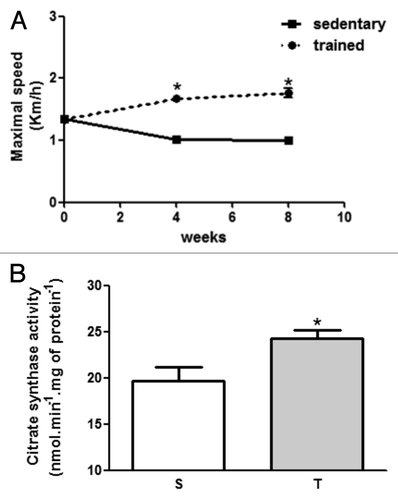
First of all, we investigated if 8 weeks of aerobic exercise was effective enough to improve aerobic capacity compared with sedentary rats. With this purpose, we performed a maximal velocity test until exhaustion, and the measurement of citrate synthase activity in soleus muscle. As observed in , trained rats (T and AT) presented an improvement in maximal running velocity during the test (S-1.0 ± 0.05 vs. T-1.76 ± 0.07 Km/h, p < 0.05). In addition, citrate synthase activity also increased in trained rats (S-19.6 ± 1.5 vs. T-24.3 ± 0.9nmol.min−1.mg of protein−1, p < 0.05).
Figure 1. (A) Maximal velocity test performed at day 0, after 4 and 8 weeks of training protocol. (B) Citrate synthase activity of sedentary and trained rats in soleus muscle. *p < 0.05 in comparison with sedentary group.
Body weight and adiposity
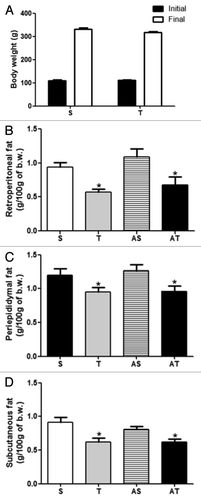
shows initial and final body weight and adiposity of sedentary and trained rats. Eight weeks of aerobic exercise did not reduce final body weight (Fig. 8A). In addition, we measured visceral and subcutaneous fat (Fig. 8B–D), and we observed an improvement of adiposity in trained rats. Trained rats (T and AT) presented reduced fat deposition in retroperitoneal, periepididymal and subcutaneous depots in relation to sedentary rats (S and AS). Acute exercise sessions did not influence this parameter (S vs. AS and T vs. AT).
Figure 2. (A) Initial and final body weight, (B) retroperitoneal adipose tissue, (C) periepididymal adipose tissue, and (D) subcutaneous adipose tissue of sedentary (S) and trained (T) rats submitted (AS and AT) or not to acute exercise session.*p < 0.05 in comparison with S and AS groups.
Serum glucose and insulin
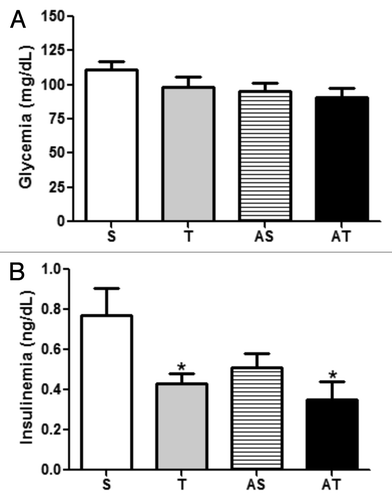
indicates the effects of chronic and acute exercise sessions on glucose and insulin circulating levels. Glycemia did not present statistical differences among groups. In contrast, aerobic exercise reduced insulin in chronic trained groups (T and AT) in comparison with sedentary groups (S and AS). Acute exercise sessions did not change insulin circulating levels in both sedentary and trained groups when compared with chronic trained groups (S vs. AS and T vs. AT).
Figure 3. (A) Glycemia, and; (B) insulinemia of sedentary (S), trained (T), sedentary that performed acute exercise session (AS), and trained rats that performed acute exercise session (AT).*p < 0.05 in comparison with S group.
Insulin content
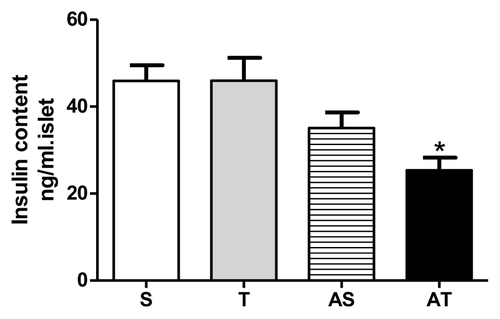
In accordance with our previous data of reduced insulinemia in trained rats, we started to investigate if training status influenced pancreatic islets. As observed in , after acute exercise, insulin content was reduced in AT group in relation to groups that did not perform acute exercise (S and T groups).
Figure 4. Insulin content of pancreatic islets in sedentary (S), trained (T), sedentary rats that performed acute exercise session (AS), and trained rats that performed acute exercise session (AT).* p < 0.05 in comparison with non-acute exercised groups (S and T).
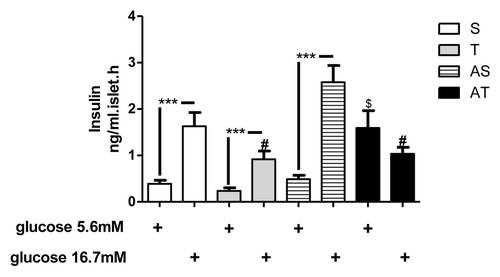
Glucose induced insulin secretion
After differences observed in insulinemia and islets insulin content, we aimed to determine the effects of training status and/or acute session of exercise on glucose-stimulated insulin secretion in isolated islets. As demonstrated in , aerobic training (8 weeks) resulted in a reduction of insulin secretion induced by high glucose concentration (16.7 mM) (S and AS vs. T and AT, p < 0.05). In addition, acute exercise resulted in elevated insulin secretion stimulated by low glucose in AT group (p < 0.05). No difference was found between low and high glucose in AT group.
Figure 5. Glucose-induced insulin secretion of pancreatic islets of sedentary (S), trained (T), sedentary rats that performed acute exercise session (AS), and trained rats that performed acute exercise session (AT). Islets were incubated with low (5.6 mM) and high (16.7 mM) glucose concentration.***p < 0.001 between low vs. high glucose. #p < 0.05 in comparison with sedentary groups in high glucose stimulus. $p < 0.05 in comparison with other groups in low glucose stimulus.
Insulin secretion stimulated by other agents
In addition to glucose-induced insulin secretion, we incubated pancreatic islets with other insulin secretagogues. First of all, we incubated islets of different groups with a depolarizing agent, arginine, which stimulates Ca2+ influx and insulin granules exocytosis independently of ATP concentration. As observed in , 8 weeks of aerobic training and/or acute exercise did not change insulin secretion stimulated by arginine 10 mM.
Figure 6. Insulin secretion stimulated by other secretagogues. (A) Arginine-induced insulin secretion; (B) Leucine-induced insulin secretion; (C) Carbachol-induced insulin secretion in sedentary (S), trained (T), sedentary rats that performed acute exercise session (AS), and trained rats (AT) that performed acute exercise session. *p < 0.05 in comparison with other groups.
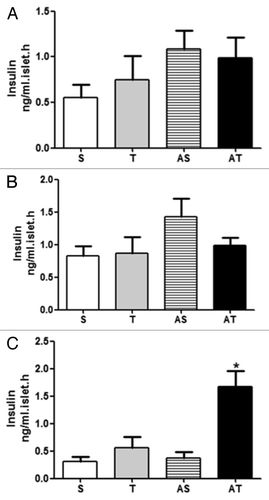
Leucine is a potent insulin secretagogue. The main mechanism for leucine-stimulated insulin secretion involves allosteric modulation of glutamate dehydrogenase. We also incubated islets from different groups with leucine 10 mM, and we did not observe any influence of chronic and acute exercise in leucine-induced insulin secretion ().
Other mechanisms by which insulin could be secreted involve cholinergic activation of muscarinic receptors in pancreatic islets. This activation results in PIP3 and DAG formation, increasing cytoplasmic Ca2+ concentration and elevating PKC activity. Carbachol is an agonist of muscarinic receptor. As demonstrated in , rats exercised that performed acute exercise for 8 weeks presented higher carbachol-induced insulin secretion (p < 0.05).
Discussion
Due to the beneficial effects of physical exercise on glycemic homeostasis, we sought to investigate its effects on insulin secretory capacity in isolated pancreatic islets. However, because physical exercise can be subdivided into chronic effects (adaptations from long-term exercise training) and acute effects (response occurred during and/or after a single session of exercise), we aimed to identify the function of pancreatic islets under these conditions.
First of all, one of the difficulties faced when dealing with animal models of exercise is to demonstrate that the experimental protocol was effective to improve physical capacity in trained groups. In this way, we performed an incremental velocity test before, after 4 weeks and at the end (8 weeks) of training protocol. As one can see, our trained group presented increased running capacity until fatigue compared with the sedentary group. In addition, we analyzed the enzyme activity of citrate synthase, which was also higher in trained groups. This enzyme is the first enzyme acting in Krebs cycle and it is well known that its activity is increased in aerobic training groups.Citation15
Afterward, we sought to analyze body weight and adiposity on different groups, since there is a direct relationship between the increase of adiposity and malfunction of pancreatic β-cells.Citation16,Citation17 As observed, trained groups presented reduced adiposity compared with sedentary groups with what could reflect positively on glycemic homeostasis and/or malfunction of pancreatic islets. The lack of significant differences between groups T and AT and S and AS seems to be directly related to chronic effects.
Focusing on glycemic homeostasis, it was not changed by chronic exercise or by acute exercise. However, an important effect of 8 weeks of physical exercise on insulinemia was observed. Once again, acute exercise did not interfere in this variable. Reduced insulinemia in response to an exercise program has been observed in other studies.Citation18,Citation19
Due to the change in insulinemia in trained groups, we sought to identify what occurred with pancreatic islets after 8 weeks of physical training and/or an acute exercise session. First of all, we tried to identify what occurred with total insulin content in pancreatic islets of different groups. As one can observe, there was no difference between trained and sedentary groups, independently of the performance of an acute exercise session. However, the association of acute exercise with chronic effects of physical exercise depressed total insulin content in pancreatic islets from group AT. Our results were different from those observed previously in literature. Oliveira et al.Citation10 observed that trained groups, independent of performing acute physical activity, presented greater insulin content in their pancreatic islets. Huang et al.Citation11 also observed an increase in total insulin content in diabetic type I rats. This difference between our and other studies can be due to the pattern of exercise and/or exhaustion protocol adopted during training and acute exercise session, respectively.
After the results obtained with total insulin content, we sought to identify the secretor insulin pattern in response to other secretagogues. Glucose is the main insulin secretagogue.Citation20 Due to the role of physical exercise in improving insulin sensibility in several peripheral tissues (among them skeletal muscle,Citation21-Citation24 which is the major tissue responsible for uptake of glucose from the bloodCitation25), it was expected that the pancreatic islets from trained rats secrete lower levels of insulin when stimulated with glucose. As it was observed, our initial hypothesis was confirmed and it corroborates data from previous studies.Citation10-Citation13 However, even with acute exercise session improving insulin sensibility of peripheral tissues,Citation26 we did not observe significant differences in insulin response stimulated by high concentrations of glucose in animals that performed acute exercise session. It demonstrates that changes in pancreatic islets in response to physical exercise require a longer time to take place.
In addition, we tried to investigate insulin secretion pathway induced by arginine (a depolarizing agent) and by leucine (which acts like an allosteric agent of glutamate dehydrogenase). No other previous study has incubated pancreatic islets from trained animals with these secretagogues. This way, we were the first to demonstrate that the effects of physical exercise on insulin secretory capacity is not mediated by post-mitochondrial mechanisms and does not depend on the depression of enzyme activity of glutamate dehydrogenase.
Besides insulin secretion stimulated by glucose and other nutrients, neural and hormonal signals can also control this action.Citation20,Citation27 Acetylcholine, for example, when released from cholinergic terminals, binds to muscarinic receptors in pancreatic islets, potentiating nutrient-induced insulin secretion by activating phospholipase C, generating diacylglycerol (DAG) and inositol-1,4,5 triphosphate (IP3). DAG activates PKC, increasing the efficiency of cytosolic Ca2+ on exocytosis of insulin granules; while IP3 releases Ca2+ from endoplasmic reticulum.Citation27,Citation28 We observed that trained animals, submitted to an acute exercise session, increase their insulin secretion stimulated by the cholinergic agent carbachol. Our group was the first to observe this fact. This way, we hypothesized that exercised animals present an adaptive mechanism of pancreatic islets that favors the activation of this pathway right after acute exhaustive exercise sessions to induce glucose uptake in skeletal muscle, restoring glycogen stores that were depleted during muscle work. However, additional studies are necessary to verify this previous information.
From our data, we conclude that physical exercise is an important tool to control glycemic homeostasis, acting directly on insulin secretory capacity stimulated by its major secretagogue, the glucose. We also demonstrated that in animals chronically trained (8 weeks), an acute exhaustive exercise session increases insulin secretory capacity stimulated by a cholinergic agent.
Methods
Animals
Male Wistar rats, 8 weeks old were used. Animals were divided into two groups: the sedentary (S) did not perform physical exercise, and the trained group (T) performed treadmill running for 8 weeks. After 8 weeks of physical training, groups S and T were subdivided into a group that performed an acute exercise session until exhaustion (AS and AT) and those that did not perform this acute exercise session (S and T). Animals were housed in groups of four animals per cage, with full access to food and water, in a 12 h light: 12 h darkness cycle (lights on at 0600 h). All procedures were in agreement with the Animal Experimentation Ethics Committee of the Institute of Biomedical Sciences of the University of São Paulo and were conducted in accordance with the institutional and national guidelines for the care and use of animals.
Physical training
During the first week, all animals were submitted to an adaptation period. After this period, animals from sedentary and trained groups completed an incremental velocity test until exhaustion. This test was used to identify training adaptations, and it was reapplied after 4 weeks of physical training and at the end of experimental protocol (8 weeks). Other criteria to determine the improvement in physical fitness was the enzyme activity of citrate synthase on the soleus muscle, as previously published.Citation29
Training protocol was composed of velocities ranging from 60% to 70% of maximal velocity reached in the incremental test, for 60 min/day, 5 d/week, during 8 weeks during dark cycle, which correspond to their active period. Acute exercise session correspond to an incremental running protocol until exhaustion performed previous the sacrifice of the animals.
Tissue collection and plasma analysis
At the end of the experimental period, animals were weighted, anesthetized and sacrificed for tissue collection. Periepididymal, retroperitoneal and subcutaneous fat pads (as an adiposity indicator), blood (glycaemia and insulinemia dosage) and pancreas (for isolation of pancreatic islets) were collected. Glycemia and insulinemia were measured by glucose oxidase kit and the radioimmunoassay method, respectively.
Isolation of pancreatic islets and incubations with different secretagogues
Islets of Langerhans were isolated from rats by pancreas digestion with collagenase as previously described by Lacy e Kostianovsky (1967). Groups of five islets were preincubated in 0.5 mL of Krebs-Henseleit (KH) (Na+ 139 mM, K+ 5 mM, Ca2+ 1 mM, Mg2+ 1 mM, Cl- 124 mM, HCO3- 24 mM), and 0.2% of albumin, with a glucose concentration of 5.6 mM, at 37°C, for 30 min. After this period, the islets were preincubated in the following solutions: KH with glucose 5.6 mM, KH with glucose 16.7 mM, KH with glucose 2.8 mM + arginin 20 mM, KH with leucine 20 mM, and KH with glucose 11.1 mM + carbachol 100 mM. The incubation period was for 60 min, and afterward, the solution was collected and insulin was measured by radioimmunoassay. In addition, groups of five islets were also collected and put into KH, and sonicated for the measurement of the insulin content.
Statistical Analysis
All data were presented as mean ± SEM. An analysis of variance (ANOVA) was used to determine differences among groups. All statistical tests were conducted using GraphPad Prism 5.0 (GraphPad software). A p < 0.05 as used to indicate statistical differences.
Acknowledgments
We thank Sandra Andreotti and Luciene Ribeiro for technical support. These studies were supported by grants and fellowships from the Foundation for Research Support of the State of Sao Paulo (FAPESP) (Proc. No. 2008/51544-2), the Coordination for the Improvement of Higher Level Personnel (CAPES) and the National Council for Scientific and Technological Development (CNPq).
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Notes
† These authors contributed equally to this work.
References
- Eizirick DL, Cnop M. ER stress in pancreatic beta cells: the thin red line between adaptation and failure. Sci Signal 2010; 3:1 - 4; PMID: 20940426
- Eizirik DL, Colli ML, Ortis F. The role of inflammation in insulitis and beta-cell loss in type 1 diabetes. Nat Rev Endocrinol 2009; 5:219 - 26; http://dx.doi.org/10.1038/nrendo.2009.21; PMID: 19352320
- Guillausseau PJ, Meas T, Virally M, Laloi-Michelin M, Médeau V, Kevorkian JP. Abnormalities in insulin secretion in type 2 diabetes mellitus. Diabetes Metab 2008; 34:Suppl 2 S43 - 8; http://dx.doi.org/10.1016/S1262-3636(08)73394-9; PMID: 18640585
- Kahn SE. The relative contributions of insulin resistance and beta-cell dysfunction to the pathophysiology of Type 2 diabetes. Diabetologia 2003; 46:3 - 19; PMID: 12637977
- Poitout V, Robertson RP. Minireview: Secondary beta-cell failure in type 2 diabetes--a convergence of glucotoxicity and lipotoxicity. Endocrinology 2002; 143:339 - 42; http://dx.doi.org/10.1210/en.143.2.339; PMID: 11796484
- Hegarty BD, Turner N, Cooney GJ, Kraegen EW. Insulin resistance and fuel homeostasis: the role of AMP-activated protein kinase. Acta Physiol (Oxf) 2009; 196:129 - 45; http://dx.doi.org/10.1111/j.1748-1716.2009.01968.x; PMID: 19245658
- Shaibi GQ, Faulkner MS, Weigensberg MJ, Fritschi C, Goran MI. Cardiorespiratory fitness and physical activity in youth with type 2 diabetes. Pediatr Diabetes 2008; 9:460 - 3; http://dx.doi.org/10.1111/j.1399-5448.2008.00407.x; PMID: 18503498
- Krause MP, Hallage T, Gama MPR, Goss FL, Robertson R, da Silva SG. Association of adiposity, cardiorespiratory fitness and exercise practice with the prevalence of type 2 diabetes in Brazilian elderly women. Int J Med Sci 2007; 4:288 - 92; http://dx.doi.org/10.7150/ijms.4.288; PMID: 18071583
- Grundy SM, Barlow CE, Farrell SW, Vega GL, Haskell WL. Cardiorespiratory fitness and metabolic risk. Am J Cardiol 2012; 109:988 - 93; http://dx.doi.org/10.1016/j.amjcard.2011.11.031; PMID: 22221951
- Oliveira CAM, Paiva MF, Mota CAS, Ribeiro C, Leme JACA, Luciano E, et al. Exercise at anaerobic threshold intensity and insulin secretion by isolated pancreatic islets of rats. Islets 2010; 2:240 - 6; http://dx.doi.org/10.4161/isl.2.4.12266; PMID: 21099318
- Huang HH, Farmer K, Windscheffel J, Yost K, Power M, Wright DE, et al. Exercise increases insulin content and basal secretion in pancreatic islets in type 1 diabetic mice. Exp Diabetes Res 2011; 2011:481427; http://dx.doi.org/10.1155/2011/481427; PMID: 21912535
- Calegari VC, Abrantes JL, Silveira LR, Paula FM, Costa JM Jr., Rafacho A, et al. Endurance training stimulates growth and survival pathways and the redox balance in rat pancreatic islets. J Appl Physiol 2012; 112:711 - 8; http://dx.doi.org/10.1152/japplphysiol.00318.2011; PMID: 22174407
- Zoppi CC, Calegari VC, Silveira LR, Carneiro EM, Boschero AC. Exercise training enhances rat pancreatic islets anaplerotic enzymes content despite reduced insulin secretion. Eur J Appl Physiol 2011; 111:2369 - 74; http://dx.doi.org/10.1007/s00421-011-1842-2; PMID: 21287194
- Ellingsgaard H, Hauselmann I, Schuler B, Habib AM, Baggio LL, Meier DT, et al. Interleukin-6 enhances insulin secretion by increasing glucagon-like peptide-1 secretion from L cells and alpha cells. Nat Med 2011; 17:1481 - 9; http://dx.doi.org/10.1038/nm.2513; PMID: 22037645
- Duscha BD, Annex BH, Johnson JL, Huffman K, Houmard J, Kraus WE. Exercise dose response in muscle. Int J Sports Med 2012; 33:218 - 23; http://dx.doi.org/10.1055/s-0031-1291323; PMID: 22261824
- Chentouf M, Dubois G, Jahannaut C, Castex F, Lajoix AD, Gross R, et al. Excessive food intake, obesity and inflammation process in Zucker fa/fa rat pancreatic islets. PLoS One 2011; 6:e22954; http://dx.doi.org/10.1371/journal.pone.0022954; PMID: 21826222
- Ferrannini E, Camastra S, Gastaldelli A, Maria Sironi A, Natali A, Muscelli E, et al. beta-cell function in obesity: effects of weight loss. Diabetes 2004; 53:Suppl 3 S26 - 33; http://dx.doi.org/10.2337/diabetes.53.suppl_3.S26; PMID: 15561918
- Zorba E, Cengiz T, Karacabey K. Exercise training improves body composition, blood lipid profile and serum insulin levels in obese children. J Sports Med Phys Fitness 2011; 51:664 - 9; PMID: 22212270
- Wieczorek-Baranowska A, Nowak A, Michalak E, Karolkiewicz J, Pospieszna B, Rutkowski R, et al. Effect of aerobic exercise on insulin, insulin-like growth factor-1 and insulin-like growth factor binding protein-3 in overweight and obese postmenopausal women. J Sports Med Phys Fitness 2011; 51:525 - 32; PMID: 21904293
- Seino S, Shibasaki T, Minami K. Dynamics of insulin secretion and the clinical implications for obesity and diabetes. J Clin Invest 2011; 121:2118 - 25; http://dx.doi.org/10.1172/JCI45680; PMID: 21633180
- Lambernd S, Taube A, Schober A, Platzbecker B, Görgens SW, Schlich R, et al. Contractile activity of human skeletal muscle cells prevents insulin resistance by inhibiting pro-inflammatory signalling pathways. Diabetologia 2012; 55:1128 - 39; http://dx.doi.org/10.1007/s00125-012-2454-z; PMID: 22282161
- Hawley JA, Lessard SJ. Exercise training-induced improvements in insulin action. Acta Physiol (Oxf) 2008; 192:127 - 35; http://dx.doi.org/10.1111/j.1748-1716.2007.01783.x; PMID: 18171435
- Zierath JR. Invited review: Exercise training-induced changes in insulin signaling in skeletal muscle. J Appl Physiol 2002; 93:773 - 81; PMID: 12133891
- Ryder JW, Chibalin AV, Zierath JR. Intracellular mechanisms underlying increases in glucose uptake in response to insulin or exercise in skeletal muscle. Acta Physiol Scand 2001; 171:249 - 57; http://dx.doi.org/10.1046/j.1365-201x.2001.00827.x; PMID: 11412137
- Bouzakri K, Koistinen HA, Zierath JR. Molecular mechanisms of skeletal muscle insulin resistance in type 2 diabetes. Curr Diabetes Rev 2005; 1:167 - 74; http://dx.doi.org/10.2174/1573399054022785; PMID: 18220592
- Xiao Y, Sharma N, Arias EB, Castorena CM, Cartee GD. A persistent increase in insulin-stimulated glucose uptake by both fast-twitch and slow-twitch skeletal muscles after a single exercise session by old rats. Age (Dordr) 2012; In press http://dx.doi.org/10.1007/s11357-012-9383-0; PMID: 22286902
- Batista TM, Ribeiro RA, Amaral AG, de Oliveira CA, Boschero AC, Carneiro EM. Taurine supplementation restores glucose and carbachol-induced insulin secretion in islets from low-protein diet rats: involvement of Ach-M3R, Synt 1 and SNAP-25 proteins. J Nutr Biochem 2012; 23:306 - 12; http://dx.doi.org/10.1016/j.jnutbio.2010.12.012; PMID: 21543213
- Gilon P, Henquin JC. Mechanisms and physiological significance of the cholinergic control of pancreatic beta-cell function. Endocr Rev 2001; 22:565 - 604; http://dx.doi.org/10.1210/er.22.5.565; PMID: 11588141
- Leandro CG, Levada AC, Hirabara SM, Manhães-de-Castro R, De-Castro CB, Curi R, et al. A program of moderate physical training for Wistar rats based on maximal oxygen consumption. J Strength Cond Res 2007; 21:751 - 6; PMID: 17685693

