
Abstract
A full understanding of embryonic endocrine pancreas development will be key to the establishment of islet replacement strategies. In particular, it is important to identify molecular pathways that establish the correct balance of specific endocrine pancreatic islet cell types. Recently, our work in the zebrafish has revealed that the correct ratio of α and β cell fates depends on the homeodomain transcription factor Mnx1 (Hb9); in the absence of functional Mnx1, β cell precursors give rise to α cells.1 Our study suggests that mnx1 may function in β cell precursors to suppress the α cell fate. Here we consider how Mnx1 may interact with other endocrine-specific transcription factors to specify β cells. Our work emphasizes the vital importance of Mnx1 for β cell development, and suggests that identifying Mnx1 transcriptional targets in β cell precursors may provide important new information of direct relevance to stem cell-based protocols to cure diabetes.
Introduction
Recent studies in the zebrafish model have clarified many details of pancreas development in this system, while emphasizing conservation of developmental mechanisms across vertebrates (reviewed in ref. Citation2). Zebrafish development is rapid; by the end of the first day of development the dorsal pancreatic bud that gives rise to the primary islet is already morphologically apparent.Citation3 During the second day of development, a secondary ventral bud forms, rapidly merging with the dorsal bud to form the nascent pancreas, which is located on the right hand side of the larval fish.Citation3 The developing zebrafish pancreas expresses a very similar complement of developmental control genes to its mammalian equivalent (reviewed in ref. Citation2). Understanding how common endocrine progenitors give rise to distinct endocrine cell types is a topic of great interest. Although our understanding of this process remains incomplete, it is clear that transcription factors play key roles. In particular, the homeodomain transcription factors Nkx2.2, Nkx6.1 and Nkx6.2 have been shown to regulate the development of α and β cell lineages in both mice and zebrafish.Citation4-Citation8 Nkx2.2−/− mutant mice and zebrafish Nkx2.2a morphants (embryos in which antisense morpholinos are used to knockdown gene function) display similar endocrine pancreas phenotypes; while both β and α cell numbers are severely reduced, the number of ghrelin-producing epsilon cells is increased.Citation7,Citation9 Nkx6.1−/− mutant mice have decreased numbers of β cells, and Nkx6.1 functions downstream of Nkx2.2 in the pathway leading to β cell fate.Citation5 By contrast, Nkx6.2−/− mutant mice display no pancreatic defects.Citation6 However, Nkx6.1/Nkx6.2 compound mutant mice have decreased numbers of α cells and show a further decrease in the number of β cells when compared with single Nkx6.1 mutants.Citation6 These findings reveal roles for Nkx6 genes in both α and β cell development. In zebrafish, single knockdowns of nkx6.1 or nkx6.2 result in a decrease in α cell number, however simultaneous knockdown suggests that β cell development requires the function of both genes.Citation8
Interactions between Mnx1 and Nkx transcription factors may determine β cell fate
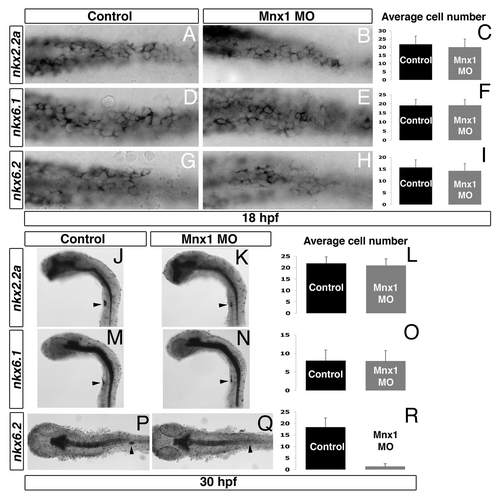
Our published data suggest that Mnx1 is a key determinant of β cell fate; using a zebrafish line carrying a Mnx1GFP reporter transgene, we have demonstrated that in the absence of functional Mnx1 protein, the Mnx1GFP-expressing β cell progenitors give rise to α cells.Citation1 To better understand the pathway from endocrine precursor to β cell fate, we have analyzed how Mnx1 knockdown might alter the expression of the nkx genes. We analyzed Mnx1 morphants at the stage when endocrine precursors are specified (18 h post fertilization; hpf) and at a stage when they have differentiated (30 hpf). Comparison of total number of nkx2.2a-, nkx6.1- and nkx6.2-expressing cells in control and Mnx1 morphants at 18 hpf indicated that knockdown of mnx1 does not alter the number of cells expressing these genes in endocrine precursors (), consistent with our previous finding that endocrine precursors are specified normally in Mnx1 morphants.Citation1 At 30 hpf, the total number of nkx2.2a- and nkx6.1-expressing cells were again similar in controls and Mnx1 morphants (). However, the number of nkx6.2-expressing cells was severely reduced in Mnx1 morphants (). As nkx6.2 expression has been shown to largely localize to the β cells by 24 hpf, this loss is not unexpected.Citation8 Recent work in the zebrafish has suggested that the dorsal bud-derived β cells do not expand further and may be less crucial for blood sugar homeostasis than the later developing ventral bud-derived β cells,Citation10 a developmental pattern similar to that of mammals. We found that the decreased expression of nkx6.2 in the developing islet is maintained through to at least 72 hpf (data not shown), by which stage ventral bud-derived cells have begun to contribute to the pancreas. These data confirm our previous findings that Mnx1 is important for β cell fates both in dorsal and ventral bud-derived β cells.Citation1 It has previously been suggested that zebrafish nkx6.1 and nkx6.2 can compensate for one another, despite their gradually segregating expression domains, such that either is sufficient to support β cell differentiation.Citation8 However, in our experimental situation, we find that although expression of nkx6.1 is retained in Mnx1 morphants, this is not sufficient to support β cell differentiation, implying that other critical factors are also missing. We can conclude that other yet-to-be-identified downstream targets of Mnx1 must also play important roles in establishing the β cell fate.
Figure 1. Whole mount in situ hybridization at 18 hpf (A,B,D,E,G,H) and 30 hpf (J,K,M,N,P,Q). Control (A,D,G,J,M,P) and Mnx1 morphants (B,E,H,K,N,Q). In situ hybridization for nkx2.2a (A,B,J,K), nkx6.1 (D,E,M,N) and nkx6.2 (G,H,P,Q). Endocrine pancreas anlagen and dorsal endocrine bud (arrow) are shown in ventral (A,B,D,E,G,H,P,Q) and lateral (J,K,M,N) views. Mean (± s.d.) number of cells expressing nkx2.2a (C,L), nkx6.1 (F,O) and nkx6.2 (I,R) markers. Results are from minimum of two independent experiments with a minimum of 20 embryos per group.
As determination of cell fate choice is regulated by multiple factors, it is likely that Mnx1 also functions in concert with other endocrine-specific transcription factors to determine β cell fate. Our previous work suggested that Mnx1 suppresses the α cell fate in the β cell lineage, possibly by downregulating the transcription of arx.Citation1 In mammals, another important regulator of Arx is the Nkx2.2 transcription factor; interestingly, Nkx2.2 can function either as an activator or as a repressor of transcription in the developing endocrine pancreas.Citation11-Citation14 Nkx2.2 may achieve this dual role by interacting with different cofactors in different cell types. However, epigenetic changes in the target DNA sequence also influence whether Nkx2.2 functions as an activator vs. as a repressor. Specifically, it has recently been shown that Nkx2.2 maintains β cell identity by binding to the hypermethylated Arx promoter to repress Arx transcription, whereas in α cells, the Arx promoter is hypomethylated and, thus, activated by Nkx2.2.Citation15,Citation16 As both Mnx1 and Nkx2.2 play important roles in suppressing α cell fate, it will be interesting to test whether these two transcription factors cooperate in the β cell lineage, for example by functioning in concert to negatively regulate Arx.
Conclusions
Here we have considered the relationships between mnx1 and nkx transcription factor encoding genes during the establishment of β cell fate in the zebrafish. Our published work suggests that Mnx1 functions primarily as a repressor of transcription to block the α cell fate.Citation1 However, as mnx1 expression is dynamic,Citation1,Citation17-Citation19 with early broad expression in the pancreatic endoderm replaced with β cell-specific expression by 24 hpf, it will ultimately be important to analyze mnx1 function at different developmental stages to gain a more complete understanding of its various roles. Moreover, we suggest that it will be important to identify the direct targets of Mnx1 in the β cell precursors, as these may provide new insights into the appropriate strategies to generate β cells in vitro for use in replacement therapies.
Acknowledgments
We thank Dr. Graeme Bell and Dr. Christopher J. Rhodes for helpful comments on the manuscript. This work was supported by the National Institute of Health (grant DK064973 to V.E.P.) and a Juvenile Diabetes Research Foundation (JDRF) fellowship to G.D.
References
- Dalgin G, Ward AB, Hao T, Beattie CE, Nechiporuk A, Prince VE. Zebrafish mnx1 controls cell fate choice in the developing endocrine pancreas. Development 2011; 138:4597 - 608; http://dx.doi.org/10.1242/dev.067736; PMID: 21989909
- Kinkel MD, Prince VE. On the diabetic menu: zebrafish as a model for pancreas development and function. Bioessays 2009; 31:139 - 52; http://dx.doi.org/10.1002/bies.200800123; PMID: 19204986
- Field HA, Dong PDS, Beis D, Stainier DYR. Formation of the digestive system in zebrafish. II. Pancreas morphogenesis. Dev Biol 2003; 261:197 - 208; http://dx.doi.org/10.1016/S0012-1606(03)00308-7; PMID: 12941629
- Sussel L, Kalamaras J, Hartigan-O’Connor DJ, Meneses JJ, Pedersen RA, Rubenstein JL, et al. Mice lacking the homeodomain transcription factor Nkx2.2 have diabetes due to arrested differentiation of pancreatic beta cells. Development 1998; 125:2213 - 21; PMID: 9584121
- Sander M, Sussel L, Conners J, Scheel D, Kalamaras J, Dela Cruz F, et al. Homeobox gene Nkx6.1 lies downstream of Nkx2.2 in the major pathway of beta-cell formation in the pancreas. Development 2000; 127:5533 - 40; PMID: 11076772
- Henseleit KD, Nelson SB, Kuhlbrodt K, Hennings JC, Ericson J, Sander M. NKX6 transcription factor activity is required for α- and β-cell development in the pancreas. Development 2005; 132:3139 - 49; http://dx.doi.org/10.1242/dev.01875; PMID: 15944193
- Pauls S, Zecchin E, Tiso N, Bortolussi M, Argenton F. Function and regulation of zebrafish nkx2.2a during development of pancreatic islet and ducts. Dev Biol 2007; 304:875 - 90; http://dx.doi.org/10.1016/j.ydbio.2007.01.024; PMID: 17335795
- Binot A-C, Manfroid I, Flasse L, Winandy M, Motte P, Martial JA, et al. Nkx6.1 and nkx6.2 regulate α- and β-cell formation in zebrafish by acting on pancreatic endocrine progenitor cells. Dev Biol 2010; 340:397 - 407; http://dx.doi.org/10.1016/j.ydbio.2010.01.025; PMID: 20122912
- Prado CL, Pugh-Bernard AE, Elghazi L, Sosa-Pineda B, Sussel L. Ghrelin cells replace insulin-producing beta cells in two mouse models of pancreas development. Proc Natl Acad Sci USA 2004; 101:2924 - 9; http://dx.doi.org/10.1073/pnas.0308604100; PMID: 14970313
- Hesselson D, Anderson RM, Beinat M, Stainier DYR. Distinct populations of quiescent and proliferative pancreatic beta-cells identified by HOTcre mediated labeling. Proc Natl Acad Sci USA 2009; 106:14896 - 901; http://dx.doi.org/10.1073/pnas.0906348106; PMID: 19706417
- Raum JC, Gerrish K, Artner I, Henderson E, Guo M, Sussel L, et al. FoxA2, Nkx2.2, and PDX-1 regulate islet beta-cell-specific mafA expression through conserved sequences located between base pairs -8118 and -7750 upstream from the transcription start site. Mol Cell Biol 2006; 26:5735 - 43; http://dx.doi.org/10.1128/MCB.00249-06; PMID: 16847327
- Doyle MJ, Loomis ZL, Sussel L. Nkx2.2-repressor activity is sufficient to specify alpha-cells and a small number of beta-cells in the pancreatic islet. Development 2007; 134:515 - 23; http://dx.doi.org/10.1242/dev.02763; PMID: 17202186
- Doyle MJ, Sussel L. Nkx2.2 regulates beta-cell function in the mature islet. Diabetes 2007; 56:1999 - 2007; http://dx.doi.org/10.2337/db06-1766; PMID: 17456846
- Anderson KR, Torres CA, Solomon K, Becker TC, Newgard CB, Wright CV, et al. Cooperative transcriptional regulation of the essential pancreatic islet gene NeuroD1 (beta2) by Nkx2.2 and neurogenin 3. J Biol Chem 2009; 284:31236 - 48; http://dx.doi.org/10.1074/jbc.M109.048694; PMID: 19759004
- Papizan JB, Singer RA, Tschen S-I, Dhawan S, Friel JM, Hipkens SB, et al. Nkx2.2 repressor complex regulates islet β-cell specification and prevents β-to-α-cell reprogramming. Genes Dev 2011; 25:2291 - 305; http://dx.doi.org/10.1101/gad.173039.111; PMID: 22056672
- Dhawan S, Georgia S, Tschen SI, Fan G, Bhushan A. Pancreatic β cell identity is maintained by DNA methylation-mediated repression of Arx. Dev Cell 2011; 20:419 - 29; http://dx.doi.org/10.1016/j.devcel.2011.03.012; PMID: 21497756
- Harrison KA, Thaler J, Pfaff SL, Gu H, Kehrl JH. Pancreas dorsal lobe agenesis and abnormal islets of Langerhans in Hlxb9-deficient mice. Nat Genet 1999; 23:71 - 5; http://dx.doi.org/10.1038/12674; PMID: 10471502
- Li H, Arber S, Jessell TM, Edlund H. Selective agenesis of the dorsal pancreas in mice lacking homeobox gene Hlxb9. Nat Genet 1999; 23:67 - 70; http://dx.doi.org/10.1038/12669; PMID: 10471501
- Wendik B, Maier E, Meyer D. Zebrafish mnx genes in endocrine and exocrine pancreas formation. Dev Biol 2004; 268:372 - 83; http://dx.doi.org/10.1016/j.ydbio.2003.12.026; PMID: 15063174

