
Abstract
Type 1 Diabetes is characterized by an absolute insulin deficiency due to the autoimmune destruction of insulin producing β-cells in the pancreatic islets. Akt1/Protein Kinase B is the direct downstream target of PI3 Kinase activation, and has shown potent anti-apoptotic and proliferation-inducing activities. This study was designed to explore whether gene transfer of constitutively active Akt1 (CA-Akt1) would promote β-cell survival and proliferation, thus be protective against experimental diabetes. In the study, a fiber-modified infectivity-enhanced adenoviral vector, Ad5RGDpK7, was used to deliver rat insulin promoter (RIP)-driven CA-Akt1 into β-cells. Our data showed this vector efficiently delivered CA-Akt1 into freshly isolated pancreatic islets, and promoted islet cell survival and β-cell proliferation in vitro. The therapeutic effect of the vector in vivo was assessed using streptozotocin (STZ)-induced diabetes mice. Two means of vector administration were explored: intravenous and intra-bile ductal injections. While direct vector administration into pancreas via bile-ductal injection resulted in local adverse effect, intravenous injection of the vectors offered therapeutic benefits. Further analysis suggests systemic vector administration caused endogenous Akt expression and activation in islets, which may be responsible, at least in part, for the protective effect of the infectivity-enhanced CA-Akt1 gene delivery vector. Taken together, our data suggest CA-Akt1 is effective in promoting β-cell survival and proliferation in vitro, but direct in vivo use is compromised by the efficacy of transgene delivery into β-cells. Nonetheless, the vector evoked the expression and activation of endogenous Akt in the islets, thus offering beneficial bystander effect against STZ-induced diabetes.
Keywords: :
Introduction
Diabetes mellitus is a common chronic disease affecting 1–2% of the population worldwide, with a trend of increasing incidence. Type 1 Diabetes (T1D) is a condition in which the insulin-producing β-cells in pancreatic islets are destroyed—primarily by a cell-mediated autoimmune response on β-cells, resulting in absolute insulin deficiency. The main treatment for T1D has been insulin therapy since its discovery in the 1920s. Indeed, exogenous insulin supplementation has transformed T1D from a fatal disease to one that is highly manageable. Nonetheless, insulin therapy is unable to effectively replicate the function of β-cells with regard to instant responses to fluctuations in blood glucose levels, and does not prevent the eventual development of diabetes-related complications. On this basis, a means to promote β-cell survival and proliferation is expected to have therapeutic benefits for T1D.
The serine/threonine kinase Akt, also called protein kinase B, is the direct downstream target of PI3 kinase (PI3K) that is activated by many growth factors and insulin, thus plays a key role in the control of cell cycle progression, cell survival, and regulation of processes that influence growth such as protein synthesis and glucose metabolism.Citation1-Citation6 Therefore, Akt is an appealing candidate for β-cell protection. More importantly, previous studies have shown the PI3K/Akt signaling pathway is highly relevant to T1D. For example, transgenic mice expressing constitutively active Akt (CA-Akt1) in β-cells show a significant increase in β-cell size and islet mass, better glucose tolerance, and are resistant to experimental diabetes.Citation5,Citation7 Mice lacking an isoform of Akt (Akt2), however, exhibit marked hyperglycemia and loss of pancreatic β-cells.Citation8 In addition, Akt1 activation via various stimuli has been shown to promote islet cell survival and proliferation.Citation1,Citation6,Citation9-Citation11 Expression of CA-Akt1 in human islets significantly improves the therapeutic outcome of islet transplantation in diabetic mice.Citation12 All of these observations suggest Akt1 plays critical roles in glucose metabolism and maintenance of pancreatic islet tissue, and thus may have therapeutic benefits for T1D.
Currently, three isoforms of Akt are identified, which include Akt1 (PKBα), Akt2 (PKBβ), and Akt3 (PKBγ). All three isoforms consist of a conserved domain structure: an N-terminal pleckstrin homology (PH) domain, a central kinase domain, and a carboxyl terminal regulatory domain. Although the three isoforms show broad tissue distribution, Akt1 is the most ubiquitously expressed and thoroughly studied. Activation of PI3K generates D3-phosphorylated phosphoinositides that bind Akt1 through the PH domain and induce the translocation of Akt1 to the plasma membrane. In the plasma membrane, Akt1 is phosphorylated at its two residues, Thr308 and Ser473.Citation13-Citation15 Thr308 is phosphorylated by phosphoinositide-dependent kinase-1 (PDK1), and Ser473 by mTOR complex 2 (mTORC2).Citation15,Citation16 There is also evidence showing Ser473 phosphorylation precedes and facilitates Thr308 phosphorylation by PDK1, and their phosphorylation leads to the activation of Akt1.Citation15,Citation16 On this basis, constitutively active forms of Akt1 (CA-Akt1) have been developed by directly targeting Akt1 to the plasma membrane, or by mimicking phosphorylation of the two sites. When direct targeting to the plasma membrane method is used, presence or absence of PH domain does not matter for the CA-Akt1. For therapeutic purpose, CA-Akt1 is advantageous because it does not require upstream stimuli.
To fully realize the therapeutic potential of CA-Akt1 for T1D, a highly efficient and specific gene delivery vector is critical. Both viral and nonviral vectors have been employed to deliver transgenes into islet cells ex vivo. These include adenoviral vectors (Ad), Adeno-associated vectors (AAV), liposomes, and a protein transduction domain (PTD)-based strategy.Citation12,Citation17-Citation22 Among them, human adenovirus serotype 5 (Ad5)-based vectors have been of high interest, attributable to their ability to infect islet cells with a relatively high level of efficiency. Previous studies and our own data have suggested a multiplicity of infection (MOI) of up to 500 viral particles per cell (VPs/cell) can be tolerated by islet cells.Citation12,Citation17 Nonetheless, we have also observed a reverse correlation between the amount of viruses used and the functionality of islets.Citation22 On this basis, it is clear that methods to increase the gene delivery efficacy of Ad5 to its target tissue will be beneficial.
Previous studies have shown genetic incorporation of extra targeting motifs into the capsid protein fiber can significantly improve Ad5 gene delivery efficacy.Citation23-Citation26 Two targeting motifs, RGD and polylysine (pK7), are of particular interest because their receptors, integrins for RGD epitope, and cell-surface proteins containing polyanion motifs such as heparin sulfate for the positively charged pK7 epitope, are widely expressed. We have shown incorporation of the RGD and pK7 motifs into Ad5 fiber significantly improves gene delivery efficiency into isolated human islets, which allows us to use fewer viral vectors for transgene delivery.Citation22,Citation26
This study was aimed to explore whether employment of this infectivity-enhanced vector, namely Ad5RGDpK7, could improve CA-Akt1 gene delivery into β-cells, and thus offer therapeutic benefits for T1D.
Results
Generation of infectivity-enhanced CA-Akt1 gene delivery vectors for pancreatic β-cells
Genetic modification of Ad5 fiber with two extra targeting motifs RGD and polylysine (pK7) has been shown to improve gene delivery efficacy for many cell types including pancreatic islet cells.Citation22,Citation26 We thus employed this infectivity-enhanced vector, Ad5RGDpK7, to deliver CA-Akt1 into β-cells. CA-Akt1 was obtained by constitutively targeting Akt1 to plasma membrane via N-terminal addition of a src myristoylation site. GFP was fused to the C-terminal end of Akt1 to facilitate the detection of transgene expression. To achieve β-cell specific gene delivery, the rat insulin promoter (RIP), was used to drive CA-Akt1 expression. The expression cassette RIP-CA-Akt1-GFP was incorporated into the deleted E1 (ΔE1) region of Ad5RGDpK7 genome. Since E1 region is essential for the initiation of Ad5 replication, the viral vector was replication deficient. The viral vector was subsequently rescued in 293 cells that stably express Ad-E1 genes, and the resultant vector was named Ad5RGDpK7.RIP-CA-Akt1 (). As negative targeting control, the CA-Akt1 expression cassette was incorporated into an E1-deleted unmodified Ad5 vector, resulting in the formation of Ad5.RIP-CA-Akt1 vector (). Other control vectors encoding RIP-driven reporters such as Ad5.RIP-Luc (for firefly luciferase) and Ad5.RIP-GFP were also constructed in a similar way ().
Figure 1. Generation and verification of CA-Akt1 gene delivery vectors. (A) Diagram of the vectors used in this study. Luc, firefly luciferase; RIP, Rat Insulin Promoter. GFP was fused to CA-Akt1 at its C-terminal end. The infectivity-enhanced vector contained two targeting motifs, RGD and pK7, in the capsid protein fiber. All transgenes were incorporated into the deleted E1 (ΔE1) region of Ad5-based vectors. (B) western blotting analysis confirmed CA-Akt1 gene delivery in human islets in vitro. Anti-β-actin staining was included to indicate equal sample loading. C-20 detected total Akt expression, and P-Ser473 and P-Thr308 detected phosphorylated Akt at corresponding sites. The arrow marks CA-Akt1 that was fused to GFP, and the asterisk (*) marks endogenous Akt. The lower bands (# marked) appear to be degraded fusion protein since they were only detected in Ad5.RIP-CA-Akt1 and Ad5RGDpK7.RIP-CA-Akt1 infected islets. (C) Immunofluorescence staining showing CA-Akt1 expression in islet cells. The human islets infected with CA-Akt1 expressing vectors were stained with anti-GFP antibody, followed by corresponding FITC-conjugated secondary antibody.
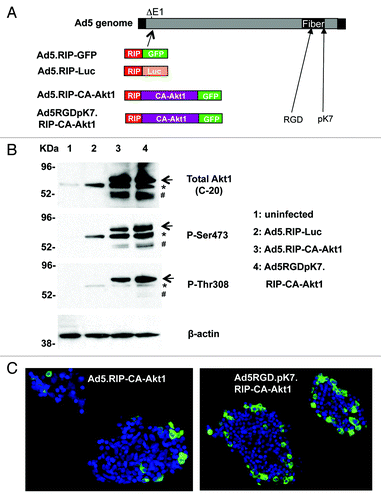
Next we examined the gene delivery efficiency mediated by the vectors using freshly isolated human islets. The islets were infected with the viruses at an MOI of 250 VPs/cell. Two days later, the islets were either lysed for western blotting assay () or processed for immunofluorescence staining () to detect Akt1 gene expression. As shown in , CA-Akt1 was successfully delivered into human islets by both Ad5 and Ad5RGDpK7, while the latter showed higher gene delivery efficiency. Staining with antibodies recognizing the phosphorylated Akt1 at either Ser473 or Thr308 showed that the transgene was active. Of note, endogenous Akt appeared to be induced by Ad5 vector infection alone, which was phosphorylated at site Ser473, and to a less degree at Thr308, consistent with our previous observation.Citation6 Of note, it has been shown phosphorylation of Ser473 precedes and facilitates that of Thr308.Citation16 The observation that P-Ser473 staining showed stronger signal than P-Thr308 staining indicates endogenous Akt might not be fully activated by Ad5 infection.
Immunofluorescence staining of the human islets with GFP confirmed CA-Akt1 expression and higher gene delivery efficacy that was mediated by the infectivity-enhanced vector (). Nonetheless, both vectors showed more gene delivery in the peripheral area of human islets, suggesting their penetration into the islet core was limited when applied in culture. Of note, in human islets, β-cells and non-β-cells are intermingled. Therefore, β-cell specific expression of GFP could be detected in the periphery of the islets.
CA-Akt1 expression improved the survival of islet cells in vitro
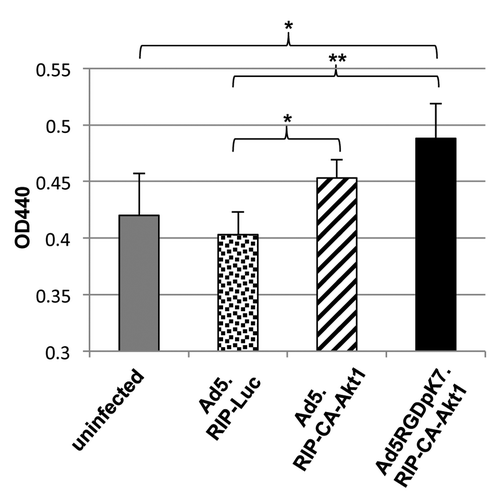
Previous studies have shown CA-Akt1 has strong protective effect on islet cells.Citation5,Citation7,Citation12 To examine whether CA-Akt1 delivered by the vectors was functional in islet cells, we performed a WST-1 based viability assay (). WST-1 is a tetrazolium salt, and it can be cleaved by mitochondrial dehydrogenase that results in the production of formazan, a dye whose intensity can be measured at OD440. Increased activity of mitochondrial dehydrogenase represents more viable cells. As shown in , CA-Akt1 expression in rat islets increased cell viability compared with control Ad5.RIP-Luc infection. More efficient CA-Akt1 gene delivery mediated by Ad5RGDpK7 showed better cell protection than unmodified Ad5, but the difference did not appear to be very significant (p = 0.0548). Compared with uninfected islets, Ad5.RIP-CA-Akt1 did not show significant cell protection (p = 0.108), while Ad5RGDpK7.RIP-CA-Akt1 did (p = 0.019), although it is not as significant as compared with Ad5.RIP-Luc (p = 0.0007). All of these may be explained by the negative effect of Ad5 infection on cell viability. Indeed, Ad5.RIP-Luc-infected islets showed lower viability than uninfected islets, although the difference was not significant (p = 0.388). Therefore, the cell viability measured here is the collective effect of CA-Akt1-mediated survival minus toxic effect of viral infection. More infectivity mediated by Ad5RGDpK7.RIP-CA-Akt1 might have resulted in more toxicity. Similar results were obtained for both rat and human islets. Taken together, these data suggest CA-Akt1 delivered by the vectors was functional in promoting cell survival.
Figure 2. CA-Akt1 expression improved islet cell survival in vitro. In the experiments, freshly isolated rat islets were infected with the vectors at a MOI of 250 VPs/cell. Three days later, islet cell viability was assessed using a WST-1 based assay kit. Higher OD440 readings mean more viable cells. The * marks p < 0.05, and ** indicates p < 0.005. The error bars represent standard deviation (n = 5). Note: Ad5.RIP-Luc infected cells showed decreased survival compared with uninfected controls, indicating there was some toxicity related to Ad5 infection, although the difference was not significant.
CA-Akt1 expression promoted islet cell proliferation in vitro
Another feature of CA-Akt1 is promoting cell proliferation. To examine whether this was true for our vectors, freshly isolated human islets were infected with the vectors, and BrdU incorporation assays were performed. BrdU, a thymidine analog, can be incorporated into the genome during DNA replication, thus is widely used as a proliferation marker. Following viral infection, the islets were cultured in BrdU-containing media for 4 d. The islets were then processed to measure BrdU incorporation using an immunochemical assay kit (). The human islets infected with CA-Akt1 expressing vectors showed significantly more BrdU incorporation than control Ad5.RIP-Luc infected ones, and the infectivity-enhanced vector Ad5RGDpK7.RIP-CA-Akt1 resulted in significantly more BrdU incorporation than Ad5.RIP-CA-Akt1 (), suggesting CA-Akt1 expression promoted islet cell proliferation.
Figure 3. CA-Akt1 expression promoted β-cell proliferation in human islets in vitro. Human islets were infected with the specified vectors, and cultured in BrdU-containing media for 4 d. (A) Total BrdU incorporation in each group of islets as assessed with a cell-based ELISA kit (n = 5). The error bars represent standard deviation. (B) BrdU incorporation at cellular level was assessed by co-staining the islets with anti-GFP (green) and anti-BrdU (red) antibodies. Nuclei were stained with Hoechst. The arrows mark the BrdU+/GFP+ cells. More significant BrdU incorporation was detected in Ad5RGDpK7.RIP-CA-Akt1 infected islets.
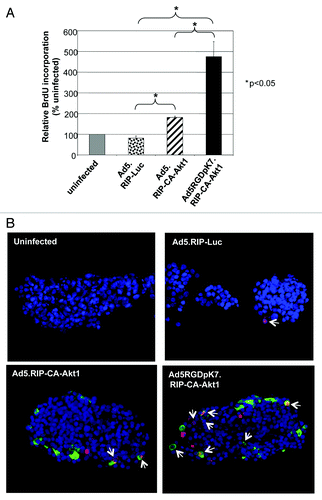
The immunochemical assay measures total BrdU incorporation in each sample. It did not distinguish whether BrdU was incorporated into β-cells, other islet cell types, or non-islet cells that were co-purified with islets in the samples. To confirm whether CA-Akt1 expression promoted BrdU incorporation in the target cells, immunofluorescence staining of the human islets was performed. Anti-BrdU, anti-GFP, and nuclei staining were used to co-stain the islet samples. Since GFP was fused to CA-Akt1 and was driven by β-cell specific promoter RIP, GFP+ cells represented β-cells expressing the transgene CA-Akt1. As shown in , we clearly detected BrdU+/GFP+ cells in islets infected with CA-Akt1 expressing vectors. More such cells were detected in Ad5RGDpK7.RIP-CA-Akt1 group than in Ad5.RIP-CA-Akt1 group (). Of note, BrdU+ cells could also be detected in control Ad5.RIP-Luc infected islets, although at much less frequency. This is in agreement with the activation of endogenous Akt by Ad5 infection observed in this study () and previous work.Citation6 Taken together, these data demonstrated CA-Akt1 expression in β-cells promoted its proliferation, and more efficient CA-Akt1 expression resulted in more significant β-cell proliferation.
Assessment of the therapeutic effect of CA-Akt1 following direct vector administration into pancreas
After the vectors were validated in vitro, we then examined whether CA-Akt1 gene delivery in vivo had any therapeutic effect for experimental T1D. In order to maximize vector administration to pancreatic islets, we directly injected the vectors into pancreas through the common bile duct of diabetic mice. Previous studies have shown such technique can allow Ad5 vectors to efficiently infect pancreatic tissue.Citation27 In this study, mouse T1D models were established with multiple-low doses (MLD) of STZ, and the vectors were injected into the pancreas through the common bile duct toward the pancreas. Three days later, the pancreas were fixed and examined for CA-Akt1 gene expression. As shown in , GFP+ β-cells were detectable, suggesting CA-Akt1 was delivered into β-cells. Nonetheless, we would like to point out that such cells were not prominent, and most islets (> 90%) did not contain GFP+ cells. This is not unexpected because CA-Akt1 was driven by RIP promoter, thus could only be expressed in cells with active insulin promoter such as β-cells, and it was difficult for the vectors to penetrate into the islets. Indeed, Taniguchi et al. has shown bile ductal administration of Ad5 vectors mediates efficient gene expression mainly in exocrine and peripheral ductal cells in the pancreas.Citation27
Figure 4. Effect of CA-Akt1 gene delivery on MLD STZ-induced T1D following intra-bile ductal injection. The CA-Akt1 expressing vectors and controls were injected into the pancreas of diabetic mice via common bile duct. Seven mice were used in each group. (A) Detection of CA-Akt1 gene expression in pancreatic islets using anti-GFP and anti-insulin co-staining. GFP+/Insulin+ cells were detected, although in very few islets (less than 10%) and often in small clusters close to ducts. (B and C) Glucose tolerance test at 1 week (B) and 6 weeks (C) after vector administration. Following overnight fasting, 1g/Kg bodyweight of glucose was administered into the peritoneal cavity of the mice, and their blood glucose levels were monitored for 2 h. At 1 week post injection, Ad5RGDpK7.RIP-CA-Akt1 treated mice showed the worst glucose tolerance, but they recovered to a level similar to normal mice by 6 weeks.
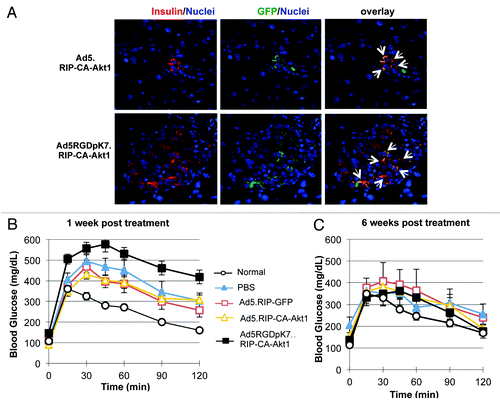
To examine whether the vectors had any therapeutic benefits, we performed glucose tolerance test to evaluate the blood glucose control of the T1D mice at weekly intervals. Surprisingly, one week after treatment, the mice that received Ad5RGDpK7.RIP-CA-Akt1 showed worse glucose tolerance than the control groups (media, Ad5.RIP-GFP and Ad5.RIP-CA-Akt1) (). At 6 weeks, however, these mice had similar glucose tolerance, if not better (difference not significant), compared with the other groups (), suggesting the mice recovered from T1D. The reason of the early adverse effect of the infectivity-enhanced vector is not clear, but may be attributable to the more severe toxicity due to its higher infectivity. Indeed, transgene expression was detected in pancreatic lymph nodes (Fig. S1), suggesting immune cells were involved in the action of CA-Akt1 expressing vectors in pancreas following bile-ductal injection.
Reversal of T1D in these MLD STZ-treated mice argued for β-cell regeneration. To investigate whether this was the case, we added BrdU into the mice’s drinking water following bile ductal injection of the vectors. As shown in , BrdU labeled β-cells were readily detectable in all MLD STZ-treated mice regardless of subsequent viral vector or media injections, suggesting β-cell proliferation occurred in response to MLD STZ-mediated β-cell injury in these mice. Ad5.RIP-CA-Akt1 and Ad5RGDpK7.RIP-CA-Akt1 treated mice showed higher percentages of BrdU+ β-cells than media or Ad5.RIP-GFP-treated mice, although the differences were not very significant (). Nonetheless, the trend indicates CA-Akt1 promoted β-cell proliferation, although marginally, and this could explain Ad5RGDpK7.RIP-CA-Akt1 treated mice recovered at a faster speed than media or Ad5.RIP-GFP groups (). Normal mice also showed BrdU incorporation in β-cells, but with significantly less frequency, and that is consistent with physiological β-cell turnover.Citation28
Figure 5. BrdU incorporation showed β-cell regeneration in MLD STZ-induced mice following intra-bile ductal injection of viral vectors. Following bile ductal injection of the vectors, BrdU-containing drinking water was provided to the mice. Six weeks later, the mice were sacrificed, and their pancreas were sectioned and processed for immunofluorescence staining using anti-insulin (red) and anti-BrdU (green) antibodies. (A) Representative images for each group. BrdU+ β-cells were detected in all groups, suggesting β-cell regeneration/proliferation played a role in the recovery of MLD STZ-induced diabetes. (B) The percentages of BrdU+ β-cells in each treatment group. (C) The differences in the percentage of BrdU+ β-cells between different groups were evaluated. p < 0.05 is defined as statistically significant. The p values were obtained using two-tail t-test between the compared groups. The symbol * stands for p < 0.05, ** for p < 0.005, and *** for p < 0.001.
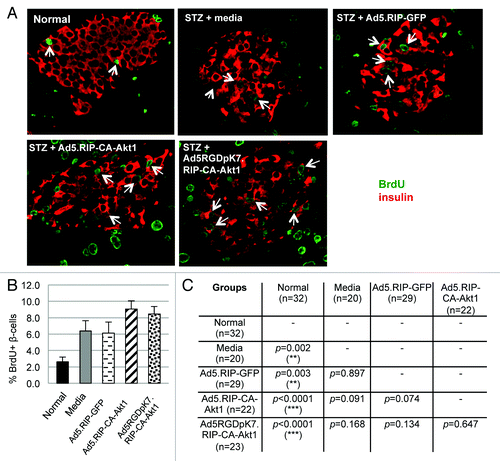
Taken together, these data suggest regeneration of β-cells was significantly induced by MLD-STZ mediated β-cell injury, which allowed the mice to recover after STZ was cleared. CA-Akt1 expression delivered by either Ad5 or Ad5RGDpK7 further promoted β-cell proliferation. However, Ad5RGDpK7.RIP-CA-Akt1 treatment did not show beneficial effect during the early days following direct injection into the pancreas, which is probably because the higher infectivity resulted in more severe toxicity.
Systemic CA-Akt1 gene delivery provided therapeutic benefits for the treatment of experimental T1D
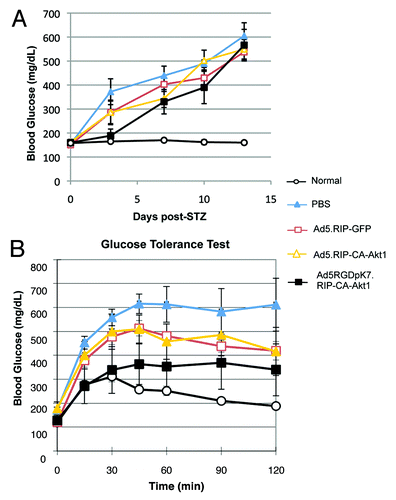
Since direct injection of Ad5 vectors into pancreas had undesirable effect, we next examined whether systemic vector administration had any benefits for T1D treatment. It is well known that intravenously injected unmodified Ad5 vectors are mainly taken up by liver, with little gene expression in other organs. Our data suggest Ad5RGDpK7 had much less liver infection than unmodified Ad5, while infection in other organs including pancreas did not change (Fig. S2). Therefore, to examine whether low level of CA-Akt1 gene delivery had any effect on the protection of T1D, we injected PBS or 1010 VPs of Ad5.RIP-GFP, Ad5.RIP-CA-Akt1, or Ad5RGDpK7.RIP-CA-Akt1 into each mouse via intravenous injection. A single high dose of STZ was used to induce T1D. As shown in , the mice that received Ad5RGDpK7.RIP-CA-Akt1 treatment showed better blood glucose control than the other treatment groups, especially in the first 7 d after STZ induction, while the mice that were not treated by any vectors (PBS group) developed hyperglycemia after 2 d (blood glucose > 300 mg/dL) (). In agreement with this, the mice that received Ad5RGDpK7.RIP-CA-Akt1 showed better glucose tolerance than other groups (). The unmodified Ad5.RIP-CA-Akt1 vector did not show difference from non-therapeutic control vector Ad5.RIP-GFP with regard to random blood glucose levels and glucose tolerance, but both groups showed some beneficial effect compared with PBS.
Figure 6. Intravenous injection of Ad5RGDpK7.RIP-CA-Akt1 improved blood glucose control of T1D mice. Six to seven mice were used in each group. (A) Random blood glucose levels of T1D mice that were treated with CA-Akt1 expressing vectors and controls. (B) Glucose tolerance test for the mice treated with different vectors one week after the vector administration. Ad5RGDpK7.RIP-CA-Akt1 treated mice showed better glucose tolerance than other groups although it was not as good as normal.
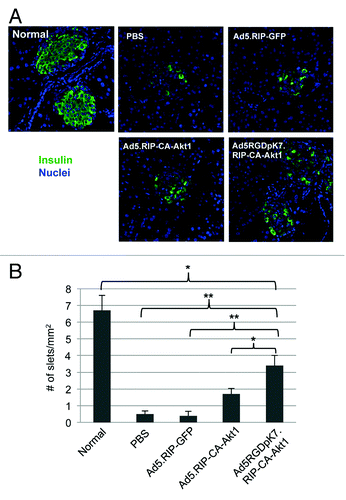
After the mice were sacrificed and pancreatic islets assessed by anti-insulin fluorescence staining, we found treatment with CA-Akt1 expressing vectors allowed more islets remaining after STZ injection (). In normal pancreatic slices, we could easily detect 6–7 islets /mm2, and all of them showed predominant insulin-producing β-cells. For STZ-induced diabetic mice, very few islets were identifiable by insulin staining after PBS or Ad5.RIP-GFP treatment. In contrast, Ad5RGDpK7.RIP-CA-Akt1 treated mice showed more identifiable islets (). Taken together, systemic CA-Akt1 gene delivery mediated by the infectivity-enhanced vector appeared to have protective effect for the pancreatic islets in vivo.
Figure 7. Evaluation of pancreatic islets in the diabetic mice that were treated with intravenously injected CA-Akt1 expressing vectors and controls. (A) Representative islet images in different treatment groups. The insulin-producing β-cells in each islet were much fewer in the STZ-induced diabetic mice than in normal ones. (B) The number of identifiable islets in each group of mice (n = 5). Ad5RGDpK7.RIP-CA-Akt1 treated diabetic mice had significantly more islets remaining than PBS, Ad5.RIP-GFP, and Ad5.RIP-CA-Akt1 treated mice, but still significantly less than normal mice. The * indicates p < 0.05, and ** indicates p < 0.005.
β-cell regeneration following systemic vector administration
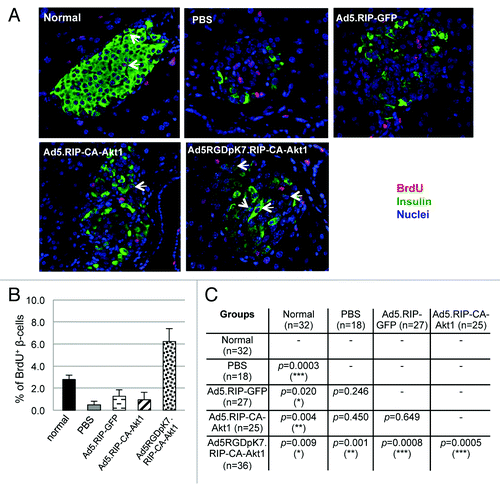
We next examined whether β-cell regeneration in STZ-induced diabetic mice was improved by intravenous injection of the CA-Akt1 expressing vectors. Using BrdU in vivo labeling, we found very few BrdU+ β-cells in PBS, Ad5.RIP-GFP, or Ad5.RIP-CA-Akt1 treated diabetic mice, which were significantly less than normal mice in which spontaneous β-cell proliferation occurred under normal physiological condition (). Ad5RGDpK7.RIP-CA-Akt1 treated mice showed significantly more β-cell proliferation than normal and all of the other treatment groups (). These data support the beneficial effect of Ad5RGDpK7.RIP-CA-Akt1 when administered systemically through intravenous injection into the single high-dose of STZ-induced diabetic mouse models. Of note, most of the BrdU+ cells in the islets of diabetic mice were non-β cells (), which is consistent with our previous studies showing treatment with a single high dose of STZ leads to rapid α-cell and δ-cell regeneration in adult mice.Citation29 Regeneration of β-cells, in contrast, seemed to be overwhelmed by the β-cell destruction induced by high-dose of STZ and subsequent severe hyperglycemia.
Figure 8. β-cell regeneration in diabetic mice that were treated with CA-Akt1 expressing or control vectors via intravenous injection. Following vector injection and a single diabetes-inducing dose of STZ, BrdU-containing drinking water was provided to the mice. Two weeks later, the mice were sacrificed, and their pancreas were sectioned and processed for immunofluorescence staining using anti-insulin (green) and anti-BrdU (red) antibodies (A). The percentages of BrdU+ β-cells in each group were quantified (B) and the differences between different groups were evaluated using two-tail t-test (C). p < 0.05 is defined as statistically significant. *p < 0.05; **p < 0.005; ***p < 0.001.
Activation of endogenous Akt in pancreatic islets
Because our data showed that exogenous delivery of CA-Akt1 with Ad5RGDpK7 in vivo had beneficial effect for STZ-induced T1D, and also because our previous studies have shown Ad5 infection of isolated islets activates endogenous Akt,Citation6 we investigated whether endogenous Akt was activated in the in vivo experimental settings above. Following intravenous injection of the vectors and STZ-mediated diabetes induction, the pancreatic tissues were processed for immunofluorescence staining with Akt1 antibodies. We discovered that both C-20 and P-Ser473 showed positive staining in the mouse islet cells (, and data not shown), suggesting endogenous Akt was induced and activated. Of note, since all three Akt isoforms (Akt1, Akt2 and Akt3) are highly homologous, we could not rule out antibody cross-reactivity. So to be precise, Akt was used here instead of Akt1.
Figure 9. Endogenous Akt activation in pancreatic islets of diabetic mice that were treated with different vectors via intravenous injection. Two weeks after intravenous injection, the mice were sacrificed, and their pancreas were sectioned and processed for immunofluorescence staining using anti-insulin (green) and anti-P-Ser473 (red) antibodies. Note: phosphorylated Akt (P-Ser473) was detected in non-β-cells in normal mice, but appeared in β-cells following Ad5RGDpK7.RIP-CA-Akt1 treated diabetic mice (arrows).
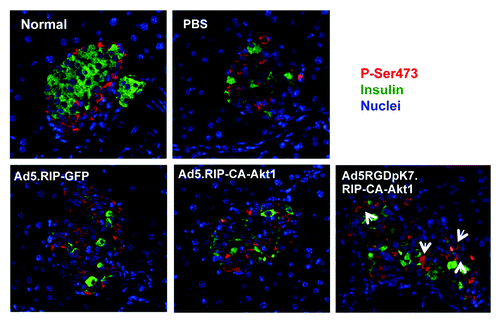
Interestingly, normal islets showed constitutive expression of endogenous Akt as well, but exclusively in non-β-cells. In STZ-injected mice, with PBS, Ad5.RIP-GFP, and Ad5.RIP-CA-Akt1 treatment, endogenous Akt expression and activation expanded in the islets, but remained in non-β-cells. In contrast, Akt activation appeared in β-cells after Ad5RGDpK7.RIP-CA-Akt1 treatment (). The active Akt did not appear to be exogenous transgene CA-Akt1 because GFP-staining was negative in these islets (data not shown). Therefore, these data suggest the protective effect of Ad5RGDpK7.RIP-CA-Akt1 was attributable to its activation of endogenous Akt in β-cells which promoted β-cell proliferation and survival following STZ-induced β-cell destruction.
Discussion
In this study, we explored the therapeutic utility of CA-Akt1 for the treatment of T1D. In order to deliver CA-Akt1 into β-cells, we generated an infectivity-enhanced adenoviral vector that mediated β-cell specific CA-Akt1 gene expression, Ad5RGDpK7.RIP-CA-Akt1. This vector showed enhanced gene delivery into β-cells, and improved cell survival and proliferation in vitro as assessed in primary islet cultures. However, the effect was more complicated in vivo. Direct intra-pancreas injection of the vectors via the common bile duct did not improve the survival and proliferation of β-cells. In fact, the treatment had opposite effect in the first two weeks, although it was recovered later. The early adverse effect could be explained by the more severe toxic effect due to higher vector infectivity. When administered systemically (i.v.), this vector showed protective effect against STZ-induced T1D. However, the protection appeared to have resulted from indirect effect, which was mediated by the induction of endogenous Akt in β-cells.
Therapeutic potential of CA-Akt1 for T1D has been implicated in two studies involving β-cell specific CA-Akt1 transgenic mice.Citation5,Citation7 Islet hypertrophy, hyperplasia and hyperinsulinaemia are the major phenotypes of the mice. As a result, the mice were resistant to STZ-induced diabetes. In contrast to CA-Akt1 transgenic expression in prostate or T lymphocytes where tumorigenesis is observed,Citation30-Citation32 no malignant transformation in pancreas is found in both pancreatic CA-Akt1 transgenic lines,Citation5,Citation7,Citation32 suggesting the activation of Akt1 in pancreas alone is not sufficient to cause transformation.Citation32 In fact, Akt appears to be expressed and active in normal mouse islets (), supporting the safety of using CA-Akt1 for islet cell protection.
Gene delivery into pancreatic islets has been a very difficult task in the gene therapy field. In order for the vectors to reach pancreas at high concentration, we adopted intra-bile ductal injection to directly introduce the vectors to the pancreas, which has also been employed by other investigators.Citation27 This strategy is similar to the clinical procedure, Endoscopic Retrograde Cholangiopancreatography (ERCP), except it is more invasive—requiring survival surgery for the mice. Using an adenoviral vector carrying CMV promoter-driven LacZ reporter, Taniquchi et al. has shown intra-common bile ductal injection leads to robust LacZ expression in pancreas, but mainly in the exocrine cells and the peripheral ducts.Citation27 In this current study, we found GFP+ (that is, CA-Akt1+) cells in pancreatic tissues were sparse, and even fewer for GFP+/insulin+ cells. The difference between our observation and Taniquchi study is apparently due to the promoters. In our system, CA-Akt1 or control GFP was driven by β-cell specific RIP promoter, so the transgene would only be expressed in β-cells. And both studies suggest penetration of the vectors into the islets was not efficient following bile ductal injection.
Interestingly, three days following intra-bile ductal injection, most GFP+ cells were found in the peri- and intra-pancreatic lymph nodes (Fig. S1). We could not find any references showing insulin promoter is active in pancreatic lymph nodes. The strong GFP expression cannot simply be explained by leaky promoter either. One could speculate that the cells with active insulin promoter (β-cells or certain precursor cells) that were infected by the vectors (Ad5.RIP-GFP, Ad5.RIP-CA-Akt1, Ad5RGDpK7.RIP-CA-Akt1) were marked as foreigners/intruders, and were transferred to the lymph nodes for immune destruction. This may also explain why there were so few GFP+/insulin+ cells in the pancreatic tissues. However, more in-depth studies are required to define the cause of the transgene expression in pancreatic lymph nodes following bile-ductal vector administration.
Another intriguing discovery made in this study is the induction and activation of endogenous Akt in islet cells following STZ and adenoviral vector injections. Akt is a critical player in mechanisms involving cell survival and proliferation. Therefore it is not surprising that its activation or upregulation is often observed in response to cellular stress, which is to exert self-protection. Previous studies have demonstrated adenoviral infection induces endogenous Akt expression and activation in different systems including primary islet culture.Citation6,Citation33,Citation34 In this study, we discovered that endogenous Akt in pancreatic islets could be induced in vivo by adenoviral vector injection, and it became more prominent with the use of the infectivity-enhanced vector, Ad5RGDpK7.RIP-CA-Akt1. In addition, we found the level of endogenous Akt activation correlated with the islet survival and blood glucose control of each mouse (unpublished data). Therefore, it is highly likely that this bystander effect was responsible, at least in part, for the therapeutic benefits observed in the systemic vector administration model. More intriguingly, the data implicate that it was the activation of endogenous Akt, but not the delivered CA-Akt1, that appears to be mainly responsible for the protective effect. This may be explained by the extent of endogenous Akt activation, which was much more than the amount of CA-Akt1 that was delivered into pancreatic β-cells, thus masked CA-Akt1 effect. Indeed, we were unable to detect CA-Akt1 expression by anti-GFP staining after i.v. injections (data not shown). This also explains why Ad5.RIP-GFP and Ad5.RIP-CA-Akt1 had similar effect in vivo—they activated endogenous Akt to a similar extent. The more significant endogenous Akt activation evoked by Ad5RGDpK7.RIP-CA- Akt1 could account for its better protection for the single-dose STZ-induced diabetes.
It should be noted that since the three Akt isoforms (Akt1, Akt2 and Akt3) are functionally and structurally conserved,Citation3,Citation4 thus there could be antibody cross-reaction. For example, for the phosphorylated Akt1 at site Ser473, it has corresponding counterparts in Akt2 and Akt3 (Ser474 and Ser472, respectively).Citation4 Therefore, the Akt we detected in this study does not necessarily represent Akt1. Indeed, it has been shown that all three Akt isoforms can be expressed in islet cells such as α-, β- and δ-cells.Citation35 The isoforms and their relative abundance in the islet cells following STZ and adenoviral vector treatment remain to be investigated.
Despite direct in vivo use of Ad5RGDpK7.RIP-CA-Akt1 appeared to be limited, this vector showed remarkable efficacy in vitro. The gene delivery efficiency was higher. The islets survived better, and showed more β-cell proliferation. Therefore, it may be used in combination with other therapies such as islet transplantation. Indeed, Rao et al. has shown CA-Akt1 gene delivery into human islets in vitro with unmodified Ad5, driven by CMV promoter—thus not β-cell specific, can improve the therapeutic efficacy of islet transplantation.Citation12 Ad5RGDpK7.RIP-CA-Akt1 is expected to offer similar benefits in this type of settings, which will be investigated in details in the future.
Materials and Methods
Antibodies
Goat polyclonal anti-human Akt1 antibody C-20 (1:200) was purchased from Santa Cruz Biotechnology. Rabbit monoclonal antibodies anti-β-actin (1:1000), and anti-phosphorylated Akt1 at sites Thr308 (P-Thr308, 1:200) and Ser473 (P-Ser473, 1:200) were purchased from Cell Signaling Technology. Mouse anti-green fluorescent protein (GFP) 11E5 (1:500) and 3E6 (1:500) monoclonal antibodies were purchased from Invitrogen. Guinea pig anti-human insulin polyclonal antibody (1:100) and mouse anti-BrdU monoclonal antibody (1:500) were purchased from Millipore Corp. The Texas-Red (TR), FITC and horse radish peroxidase (HRP)-conjugated secondary antibodies, which include anti-mouse, anti-goat, anti-guinea pig, and anti-rabbit antibodies, were purchased from Jackson ImmunoResearch Laboratories Inc.The nuclear dye Hoechst 33342 was obtained from Sigma-Aldrich Corp., and used at a final concentration of 2 µg/ml.
Generation of adenoviral vectors
In order to generate Ad5-based gene delivery vectors, first a shuttle vector carrying the transgene expression cassette was constructed. Briefly, CA-Akt1 was generated by incorporation of a src myristoylation site at the N-terminus of human Akt1 so that it was always targeted to membrane and thus was constitutively active. GFP was fused to its C-terminal end to facilitate detection of gene expression. Rat insulin promoter (RIP) was used to drive CA-Akt1 expression. With conventional cloning procedures, the expression cassette RIP-CA-Akt1-GFP was transferred into pShuttle vector (Agilent Technologies) that would allow subsequent incorporation into the deleted E1 region of adenoviral genome via homologous recombination. The resultant plasmid was named pShuttle.RIP-CA-Akt1. The shuttle vectors pShuttle.RIP-GFP and pShuttle.RIP-Luc were constructed similarly.
Construction of Ad5 genome containing extra targeting motifs RGD and pK7 in fiber gene was described previously.Citation26 Plasmid containing this genome, namely pAd5RGDpK7, was used as backbone to generate infectivity-enhanced CA-Akt1 gene delivery vector. Unmodified pAd5 was used as backbone for control vectors.
Incorporation of the transgene expression cassette into adenoviral genome was achieved by homologous recombination between the pShuttle vectors and the backbone plasmids. Specifically, the pShuttle vectors were linearized with Pme I digestion, and the backbones pAd5 or pAd5RGDpK7 were linearized by Pac I digestions. After the fragments were purified by phenol extraction, they were co-transformed into bacteria BJ5183 via electroporation with a molar ratio of 1:10 (backbone: shuttle). Recombinants were screened using various enzyme digestions and PCR analysis after DNA mini-prep from the colonies. After the positive clones were identified, their DNA was transferred into DH10B for stable DNA amplification and purification.
In order to rescue the resultant viruses, the recombinant plasmids were digested with Pac I (to release Ad5 genome from the plasmid), purified, and transfected into 293 cells that stably express Ad-E1 proteins. After the viruses were formed, and complete CPE reached, they were amplified in a large number of 293 cells and purified by double CsCl gradient centrifugation as described previously.Citation36,Citation37
Islet isolation and culture
Freshly isolated human islets (≥ 95% viability and ≥ 90% purity) were obtained from the Integrated Islet Distribution Program (IIDP) that was funded by National Institute of Diabetes and Digestive and Kidney Diseases (NIDDK) with support from Juvenile Diabetes Research Foundation (JDRF). The islets were cultured in complete Miami media supplemented with 10% fetal bovine serum (FBS) and 20 mg/L ciprofloxacin hydrochloride, at 37°C, in a humidified incubator containing 5% CO2.
Rat islets were isolated from Sprague-Dawley rats (~250 g) as following. Under anesthesia with intraperitoneal injection of Ketamine/Xylazine, each rat underwent midline incision, and then its common bile duct was cannulated with tubing oriented toward the pancreas. Dissociation enzyme solution containing 0.2 mg/ml of Liberase RI and 0.1 mg/ml of DNase I (Roche Applied Science) in Hank’s buffered saline solution (HBSS) was injected into the pancreas via the common bile duct at a dosage of 1 ml per 100 g body weight. The distended pancreas was dissected, cut into small pieces, transferred to a 50-ml conical tube containing 10 ml of the dissociation enzyme solution, and incubated in a 37°C waterbath with gentle agitation for ~20 min. Then 5 ml of quenching buffer (10% FBS and 0.1 mg/ml DNase I in HBSS) was added into the tube, and the tissue suspension was filtered through a 500 µm Netwell filter into a clean tube. To purify the islets, the tissue suspension was first centrifuged at 1200 rpm at 4°C for 5 min. After removal of supernatant, the tissue was washed twice with quenching buffer, and Eurocollins solution containing 20% FBS. Then the tissue was re-suspended in Euroficoll 1.037, and loaded on top of Euroficoll 1.108 in a clean tube for gradient centrifugation. The islets were separated into the interface of the two layers after centrifugation at 1500 rpm, 4°C for 10 min without brake. The purified islets were collected from the interface, washed twice with culture media. After a small aliquot was taken for quantification, and the remaining islets were re-suspended in complete culture media and cultured as described above.
Ad5 infection of islets
Freshly isolated human or rat islets were distributed in ultra-low attachment culture flasks or plates with a density of 500 IEQ/ml. Ad5 vectors equivalent to a multiplicity of infection (MOI) of 250 viral particles per cell (VPs/cell) were used to infect the islets, of which 1 IEQ = 2000 cells were used in the calculations. Uninfected islets were processed in parallel as a control. The culture media were refreshed 24 h later, and every other day thereafter.
BrdU-based proliferation assay
To detect cell proliferation, immediately following adenoviral vector infection of human islets, BrdU was added into the culture media at a final concentration of 25 μg/ml. The islets were cultured for 4 d with media (containing BrdU) refreshed every other day. BrdU incorporation was then assessed by either immunofluorescence staining or immunochemical assays. Immunofluorescence staining was performed as described below. Immunochemical assessment, a cell-based ELISA assay, of BrdU incorporation was performed using BrdU Cell Proliferation kit following the manufacturer’s instruction (EMD Milliopore). Briefly, the islets (cultured in 96-well plates, in suspension) were spun down using a microplate centrifuge, and fixed with fixing solution for 30 min. After washing, the islets were incubated with anti-BrdU monoclonal antibody, and then with Peroxidase-conjugated secondary antibody. Following washes with washing buffer and final water wash, peroxidase substrate was added into each well. The reaction was stopped 30 min later with stop solution. The plate was read at OD450nm using PowerWaveHT 340 plate reader (BioTek). The higher OD reading indicates more BrdU incorporation in the sample.
Cell viability assay
Cell viability was measured using a WST-1 based assay kit following the manufacturer’s protocol (Millipore). Briefly, the islets infected with different viruses were cultured in 96-well plates (30 IEQ/100 µl/well, 5 wells per infection group). Three days after viral infection, 10 µl WST-1 was added into each well, and continued to culture for 4 h. OD440nm was then measured with PowerWaveHT 340 plate reader (BioTek). Higher readings mean more viable cells.
Tissue preparation and immunofluorescence staining
Both OCT-embedded (for isolated human islets) and paraffin-embedded (mouse pancreas) tissue slices were used in this study. For isolated human islets, following infection and culture, the islets were collected by centrifugation at 1000 rpm for 5 min, washed with PBS twice, and then fixed in 4% formaldehyde for 1 h at room temperature. After washing with PBS, the islets were incubated in 20% sucrose overnight at 4°C. The islets were then spun down, and re-suspended in a 1:1 mixture of 20% sucrose and OCT. Next, the islets were transferred into specimen molds and frozen on dry ice immediately. They were stored at -70°C until being sectioned into 7 μm slices. Immunofluorescence staining of the slices with anti-GFP, anti-BrdU antibodies and nuclear dye Hoechest was performed as described previously.Citation29
For pancreatic tissues, after the mice were sacrificed, their pancreas were dissected and fixed in 10% buffered formalin solution overnight, and then processed for paraffin-embedding and sectioning with standard procedures. Deparaffinization, antigen retrieval, and immunofluorescence staining were performed as previously described.Citation29
Western blotting assays
Freshly isolated human islets were infected with Ad5.RIP-Luc, Ad5.RIP-CA-Akt1, or Ad5RGDpK7.RIP-CA-Akt1 at MOI of 250 VPs/cell. Uninfected islets were used as control. Two days after infection, the islets were collected, washed with phosphate-buffered saline (PBS), and lysed in Reporter Lysis Buffer (Promega) containing 1% SDS. Following one freeze/thaw cycle, SDS sample buffer were added into cell lysates, and the samples boiled for 5 min. After cooling down on ice, the cell lysates were processed for SDS-PAGE and western blotting assays following standard procedures.
Establishment of mouse diabetes models
C57BL/6 mice (Jackson Labs) and NOD-SCID mice (Frederick Cancer Research) were used in this study. All animal procedures followed the guidelines set by the Institutional Animal Care and Use Committee at both the University of Alabama at Birmingham and Tulane University. The animal protocol approval numbers are 100308682 and 4254, respectively.
To establish diabetes mouse models with multiple-low doses of streptozotocin (STZ), 8–10 weeks old mice were treated with 40 mg/kg bodyweight of STZ for 5 consecutive days via intra-peritoneal (i.p.) injections. Four hours following each STZ injection, 100 µl of 20% glucose were administered into each mouse to prevent hypoglycemia caused by sudden release of large amounts of insulin due to STZ-induced β-cell destruction. Their blood glucose was monitored twice a week using AlphaTRAK Blood Glucose Monitoring System (Abbott Laboratories).
Establishment of diabetes models with a single-dose of STZ were described in our previous study.Citation29 Briefly, the mice (8–10 weeks old) were fasted overnight, and then injected with 130 mg/kg bodyweight of STZ. Four hours later, the mice received 200 µl of 20% glucose via i.p. injection to prevent hypoglycemia. Their blood glucose was monitored as described above.
Bile ductal administration of Ad5 vectors
A survival bile-duct injection protocol was developed by modifying what Taniquchi et al. described before.Citation27 Specifically, C57BL/6 mice were under anesthesia with ketamine + xylazine. Following a midline incision (< 2 cm long), the common bile duct was exposed by carefully displacing intestines. For injection, a 31G syringe containing 5 × 109 VPs diluted in 250 ul of RPMI basal media was inserted into the bile duct toward the pancreas. After injection, the intestines were carefully placed back in position. The peritoneal cavity and the skin were closed with absorbable PDS*II sutures (Ethicon, Inc.). Then the mice were placed on heating pads during recovery, and standard care procedures were followed. Buprenex (0.1 mg/kg body weight) was administered subcutaneously after the mice regained consciousness, and every 12 h thereafter as needed.
Quantification of islet cells
Quantification of BrdU+ β-cells were performed as described previously.Citation29 Briefly, following immunofluorescence staining, the numbers of BrdU+ β-cells and total β-cells in each islet were counted from the islet microscopy images stored in Adobe Photoshop. 20–35 representative islets from five mice were counted. Islets with < 10 cells were excluded.
Statistical analysis
All statistical analyses were performed using GraphPad Prism biostatistics software (GraphPad Software, Inc.). The data were expressed as mean, and the error bars represent standard error of the mean (SEM) unless stated otherwise. p < 0.05 was considered statistically significant.
Additional material
Download Zip (970.2 KB)Acknowledgments
The authors would like to thank Tie Han for his technical support, and Dr Juan L. Contreras and his technical personnel Stacie Bryant, Hughston Head and Brandon Moore for their assistance on rat islet isolation. The authors would also like to acknowledge several core facilities in the University of Alabama at Birmingham (UAB) and Tulane University. These include UAB High Resolution Imaging Facility, UAB Center for Metabolic Bone Disease Core Laboratory, UAB Animal Physiology Core in the Diabetes Research and Training Center, the COBRE core facility in Tulane Hypertension and Renal Center of Excellence and Tulane Histology Research Laboratory.
This research was supported by the Juvenile Diabetes Research Foundation grant 27–2009–378 and the National Institute of Diabetes and Digestive and Kidney Diseases grant R01DK081463 (HW). J.B.P. was supported by grants 09GRNT2160024 from the American Heart Association Greater Southeast Affiliate and R01DK072154 from the National Institutes of Health.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Supplemental Material
Supplemental materials may be found here:
http://www.landesbioscience.com/journals/islets/article/22721/
Notes
† These authors contributed equally to this work.
References
- Aikin R, Rosenberg L, Maysinger D. Phosphatidylinositol 3-kinase signaling to Akt mediates survival in isolated canine islets of Langerhans. Biochem Biophys Res Commun 2000; 277:455 - 61; http://dx.doi.org/10.1006/bbrc.2000.3664; PMID: 11032744
- Bos JL. A target for phosphoinositide 3-kinase: Akt/PKB. Trends Biochem Sci 1995; 20:441 - 2; http://dx.doi.org/10.1016/S0968-0004(00)89097-0; PMID: 8578585
- Brazil DP, Yang ZZ, Hemmings BA. Advances in protein kinase B signalling: AKTion on multiple fronts. Trends Biochem Sci 2004; 29:233 - 42; http://dx.doi.org/10.1016/j.tibs.2004.03.006; PMID: 15130559
- Hanada M, Feng J, Hemmings BA. Structure, regulation and function of PKB/AKT–a major therapeutic target. Biochim Biophys Acta 2004; 1697:3-16.
- Tuttle RL, Gill NS, Pugh W, Lee JP, Koeberlein B, Furth EE, et al. Regulation of pancreatic beta-cell growth and survival by the serine/threonine protein kinase Akt1/PKBalpha. Nat Med 2001; 7:1133 - 7; http://dx.doi.org/10.1038/nm1001-1133; PMID: 11590437
- Icyuz M, Bryant SM, Fortinberry HK, Molakandov K, Siegal GP, Contreras JL, et al. Adenovirus infection activates akt1 and induces cell proliferation in pancreatic islets1. Transplantation 2009; 87:821 - 4; http://dx.doi.org/10.1097/TP.0b013e318199c686; PMID: 19300183
- Bernal-Mizrachi E, Wen W, Stahlhut S, Welling CM, Permutt MA. Islet beta cell expression of constitutively active Akt1/PKB alpha induces striking hypertrophy, hyperplasia, and hyperinsulinemia. J Clin Invest 2001; 108:1631 - 8; PMID: 11733558
- Garofalo RS, Orena SJ, Rafidi K, Torchia AJ, Stock JL, Hildebrandt AL, et al. Severe diabetes, age-dependent loss of adipose tissue, and mild growth deficiency in mice lacking Akt2/PKB beta. J Clin Invest 2003; 112:197 - 208; PMID: 12843127
- Contreras JL, Smyth CA, Bilbao G, Young CJ, Thompson JA, Eckhoff DE. Simvastatin induces activation of the serine-threonine protein kinase AKT and increases survival of isolated human pancreatic islets. Transplantation 2002; 74:1063 - 9; http://dx.doi.org/10.1097/00007890-200210270-00001; PMID: 12438947
- Maeda H, Rajesh KG, Maeda H, Suzuki R, Sasaguri S. Epidermal growth factor and insulin inhibit cell death in pancreatic beta cells by activation of PI3-kinase/AKT signaling pathway under oxidative stress. Transplant Proc 2004; 36:1163 - 5; http://dx.doi.org/10.1016/j.transproceed.2004.04.018; PMID: 15194403
- Srinivasan S, Bernal-Mizrachi E, Ohsugi M, Permutt MA. Glucose promotes pancreatic islet beta-cell survival through a PI 3-kinase/Akt-signaling pathway. Am J Physiol Endocrinol Metab 2002; 283:E784 - 93; PMID: 12217896
- Rao P, Roccisana J, Takane KK, Bottino R, Zhao A, Trucco M, et al. Gene transfer of constitutively active Akt markedly improves human islet transplant outcomes in diabetic severe combined immunodeficient mice. Diabetes 2005; 54:1664 - 75; http://dx.doi.org/10.2337/diabetes.54.6.1664; PMID: 15919787
- Alessi DR, Andjelkovic M, Caudwell B, Cron P, Morrice N, Cohen P, et al. Mechanism of activation of protein kinase B by insulin and IGF-1. EMBO J 1996; 15:6541 - 51; PMID: 8978681
- Kohn AD, Takeuchi F, Roth RA. Akt, a pleckstrin homology domain containing kinase, is activated primarily by phosphorylation. J Biol Chem 1996; 271:21920 - 6; http://dx.doi.org/10.1074/jbc.271.36.21920; PMID: 8702995
- Scheid MP, Woodgett JR. Unravelling the activation mechanisms of protein kinase B/Akt. FEBS Lett 2003; 546:108 - 12; http://dx.doi.org/10.1016/S0014-5793(03)00562-3; PMID: 12829245
- Sarbassov DD, Guertin DA, Ali SM, Sabatini DM. Phosphorylation and regulation of Akt/PKB by the rictor-mTOR complex. Science 2005; 307:1098 - 101; http://dx.doi.org/10.1126/science.1106148; PMID: 15718470
- Contreras JL, Bilbao G, Smyth CA, Jiang XL, Eckhoff DE, Jenkins SM, et al. Cytoprotection of pancreatic islets before and soon after transplantation by gene transfer of the anti-apoptotic Bcl-2 gene. Transplantation 2001; 71:1015 - 23; http://dx.doi.org/10.1097/00007890-200104270-00001; PMID: 11374395
- Garcia-Ocana A, Takane KK, Reddy VT, Lopez-Talavera JC, Vasavada RC, Stewart AF. Adenovirus-mediated hepatocyte growth factor expression in mouse islets improves pancreatic islet transplant performance and reduces beta cell death. J Biol Chem 2003; 278:343 - 51; http://dx.doi.org/10.1074/jbc.M207848200; PMID: 12403787
- Kapturczak MH, Flotte T, Atkinson MA. Adeno-associated virus (AAV) as a vehicle for therapeutic gene delivery: improvements in vector design and viral production enhance potential to prolong graft survival in pancreatic islet cell transplantation for the reversal of type 1 diabetes. Curr Mol Med 2001; 1:245 - 58; http://dx.doi.org/10.2174/1566524013363979; PMID: 11899074
- Mendoza V, Klein D, Ichii H, Ribeiro MM, Ricordi C, Hankeln T, et al. Protection of islets in culture by delivery of oxygen binding neuroglobin via protein transduction. Transplant Proc 2005; 37:237 - 40; http://dx.doi.org/10.1016/j.transproceed.2004.12.270; PMID: 15808606
- Ribeiro MM, Klein D, Pileggi A, Molano RD, Fraker C, Ricordi C, et al. Heme oxygenase-1 fused to a TAT peptide transduces and protects pancreatic beta-cells. Biochem Biophys Res Commun 2003; 305:876 - 81; http://dx.doi.org/10.1016/S0006-291X(03)00856-8; PMID: 12767912
- Contreras JL, Wu H, Smyth CA, Eckstein CP, Young CJ, Seki T, et al. Double genetic modification of adenovirus fiber with RGD polylysine motifs significantly enhances gene transfer to isolated human pancreatic islets. Transplantation 2003; 76:252 - 61; http://dx.doi.org/10.1097/01.TP.0000066361.02042.CA; PMID: 12865820
- Dmitriev I, Krasnykh V, Miller CR, Wang M, Kashentseva E, Mikheeva G, et al. An adenovirus vector with genetically modified fibers demonstrates expanded tropism via utilization of a coxsackievirus and adenovirus receptor-independent cell entry mechanism. J Virol 1998; 72:9706 - 13; PMID: 9811704
- Han T, Tang Y, Ugai H, Perry LE, Siegal GP, Contreras JL, et al. Genetic incorporation of the protein transduction domain of Tat into Ad5 fiber enhances gene transfer efficacy. Virol J 2007; 4:103; http://dx.doi.org/10.1186/1743-422X-4-103; PMID: 17958897
- Wickham TJ, Roelvink PW, Brough DE, Kovesdi I. Adenovirus targeted to heparan-containing receptors increases its gene delivery efficiency to multiple cell types. Nat Biotechnol 1996; 14:1570 - 3; http://dx.doi.org/10.1038/nbt1196-1570; PMID: 9634823
- Wu H, Seki T, Dmitriev I, Uil T, Kashentseva E, Han T, et al. Double modification of adenovirus fiber with RGD and polylysine motifs improves coxsackievirus-adenovirus receptor-independent gene transfer efficiency. Hum Gene Ther 2002; 13:1647 - 53; http://dx.doi.org/10.1089/10430340260201734; PMID: 12228019
- Taniguchi H, Yamato E, Tashiro F, Ikegami H, Ogihara T, Miyazaki J. beta-cell neogenesis induced by adenovirus-mediated gene delivery of transcription factor pdx-1 into mouse pancreas. Gene Ther 2003; 10:15 - 23; http://dx.doi.org/10.1038/sj.gt.3301846; PMID: 12525833
- Dor Y, Brown J, Martinez OI, Melton DA. Adult pancreatic beta-cells are formed by self-duplication rather than stem-cell differentiation. Nature 2004; 429:41 - 6; http://dx.doi.org/10.1038/nature02520; PMID: 15129273
- Zhang Y, Zhang Y, Bone RN, Cui W, Peng JB, Siegal GP, et al. Regeneration of pancreatic non-β endocrine cells in adult mice following a single diabetes-inducing dose of streptozotocin. PLoS One 2012; 7:e36675; http://dx.doi.org/10.1371/journal.pone.0036675; PMID: 22586489
- Jones RG, Parsons M, Bonnard M, Chan VS, Yeh WC, Woodgett JR, et al. Protein kinase B regulates T lymphocyte survival, nuclear factor kappaB activation, and Bcl-X(L) levels in vivo. J Exp Med 2000; 191:1721 - 34; http://dx.doi.org/10.1084/jem.191.10.1721; PMID: 10811865
- Majumder PK, Yeh JJ, George DJ, Febbo PG, Kum J, Xue Q, et al. Prostate intraepithelial neoplasia induced by prostate restricted Akt activation: the MPAKT model. Proc Natl Acad Sci U S A 2003; 100:7841 - 6; http://dx.doi.org/10.1073/pnas.1232229100; PMID: 12799464
- Yang ZZ, Tschopp O, Baudry A, Dümmler B, Hynx D, Hemmings BA. Physiological functions of protein kinase B/Akt. Biochem Soc Trans 2004; 32:350 - 4; http://dx.doi.org/10.1042/BST0320350; PMID: 15046607
- Liu Q, White LR, Clark SA, Heffner DJ, Winston BW, Tibbles LA, et al. Akt/protein kinase B activation by adenovirus vectors contributes to NFkappaB-dependent CXCL10 expression. J Virol 2005; 79:14507 - 15; http://dx.doi.org/10.1128/JVI.79.23.14507-14515.2005; PMID: 16282450
- Miller-Jensen K, Janes KA, Wong YL, Griffith LG, Lauffenburger DA. Adenoviral vector saturates Akt pro-survival signaling and blocks insulin-mediated rescue of tumor necrosis-factor-induced apoptosis. J Cell Sci 2006; 119:3788 - 98; http://dx.doi.org/10.1242/jcs.03102; PMID: 16940353
- Muller D, Huang GC, Amiel S, Jones PM, Persaud SJ. Gene expression heterogeneity in human islet endocrine cells in vitro: the insulin signalling cascade. Diabetologia 2007; 50:1239 - 42; http://dx.doi.org/10.1007/s00125-007-0671-7; PMID: 17440705
- Wu H, Dmitriev I, Kashentseva E, Seki T, Wang M, Curiel DT. Construction and characterization of adenovirus serotype 5 packaged by serotype 3 hexon. J Virol 2002; 76:12775 - 82; http://dx.doi.org/10.1128/JVI.76.24.12775-12782.2002; PMID: 12438602
- Wu H, Curiel DT. Fiber-modified adenoviruses for targeted gene therapy. Methods Mol Biol 2008; 434:113 - 32; PMID: 18470642

