Abstract
Recruitment of genes to the nuclear periphery upon transcriptional activation is a common phenomenon in Saccharomyces cerevisiae. We have recently identified DNA elements called gene recruitment sequences (GRSs) in the promoters of genes that are recruited to the nuclear periphery. These elements are necessary for peripheral targeting of genes. GRSs also function as DNA zip codes: they are sufficient to target an ectopic locus to the nuclear periphery. Targeting promotes full transcription and involves the interaction of promoters with the Nuclear Pore Complex (NPC). GRSs are widespread across the yeast genome, and are enriched in the promoters of genes induced by protein folding stress. Here, we place these observations in the context of the more global topic of genome organization and speculate about how the position of genes impacts their expression.
Introduction
The study of nuclear architecture and how it influences gene expression has long represented a fascinating challenge for cell biologists. Chromosomes fold and are non-randomly arranged within the nucleus.Citation1–Citation3 Therefore, individual genes on these chromosomes will have unique positions within the nucleus and may be exposed to distinct molecular environments. The positioning of genes in the nuclear space is also highly dynamic and can change rapidly in response to spatial and temporal cues.Citation4–Citation7 Repositioning of genes (and chromosomes) within the nucleus can therefore change inter-chromosomal and intra-chromosomal interactionsCitation8 and the association of genes with nuclear structures (reviewed in ref. Citation9). Changes in gene localization are often accompanied by changes in gene expression, suggesting that nuclear environment can influence transcription, perhaps by exploiting the heterogeneous distribution of trans-acting regulators within the nucleus.Citation10,Citation11 Here we discuss new conceptual and mechanistic insights into the molecular mechanisms that control the subnuclear positioning of genes in yeast.
Localization of Genes is Controlled by DNA Zip Codes
The sequence of genomes not only encodes proteins and binding sites that regulate gene expression, but also provides information that dictates gene positioning. In Drosophila, DNA sequences known as Scaffold Associated Regions (SARs) interact with the nuclear matrix.Citation12,Citation13 Additionally, microscopic analysis of Drosophila chromosome 2 revealed that specific loci serve as nuclear envelope association sites.Citation14 Genome-wide studies in humans and Drosophila have identified large, megabase scale domains that associate with the nuclear lamina. These lamin-associated domains or LADs, are enriched for Oct-1 binding sites, a protein known to localize to the nuclear lamina.Citation15 Taken together, these studies have suggested that the association of chromatin with specific nuclear landmarks is conferred or influenced by DNA sequence and that trans acting proteins with particular localization within the nucleus bind these sequences. Consistent with this idea, we have recently identified small, well-defined sequence elements from promoters that can function as “DNA zip codes”: elements that are necessary and sufficient to control the subnuclear localization of chromosomal loci.
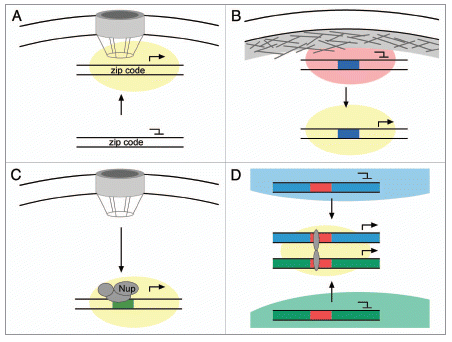
A number of genes in yeast localize in the nucleoplasm when transcriptionally repressed but move to the nuclear periphery upon activation.Citation16–Citation22 Relocalization to the nuclear periphery is associated with an interaction with the nuclear pore complex (NPC; ).Citation17 We found that the targeting of one such gene, INO1, to the nuclear periphery required two DNA elements within its promoter.Citation22 These elements are distinct from the previously characterized upstream activating elements and, although INO1 is targeted to the nuclear periphery upon activation, these targeting elements appear to function independently of these enhancer sequences.Citation22 We call these elements Gene Recruitment Sequences (GRSs).Citation22 The GRS elements are necessary for INO1 targeting to the nuclear periphery and sufficient to target an ectopic locus to the nuclear periphery. When removed from the INO1 promoter and integrated at a location that normally localizes in the nucleoplasm, they are sufficient to target this ectopic locus to the nuclear periphery. DNA zip codes such as the GRSs are small: insertion of the 8 base pair GRS I at an ectopic location is sufficient to confer peripheral localization.Citation22
The localization of transcriptionally active genes to the nuclear periphery is a common phenomenon. The hsp70 gene cluster in Drosophila physically interacts with the NPC and localizes to the nuclear periphery.Citation23 In mouse tissue culture cells, some genes that are derepressed by treatment with the histone deacetylase inhibitor TSA also relocalize to the nuclear periphery and physically interact with the NPC.Citation24 Furthermore, in male flies, the entire X chromosome localizes at the nuclear periphery and interacts with the NPC.Citation25 In S. cerevisiae where this phenomenon is best characterized, hundreds of active genes are thought to interact with the NPC.Citation16–Citation20,Citation22
Gene localization has been studied using DNA-FISHCitation26,Citation27 or variations of the GFP-Lac repressor/Lac operator system,Citation28 which allow the visualization of a specific genetic locus with respect to nuclear landmarks. Genome-wide chromatin immunoprecipitation experiments with NPC proteins has defined a subset of the yeastCitation16,Citation17 and fly genomeCitation29–Citation31 that interacts with the NPC and, in some cases is localized at the nuclear periphery (see below for exceptions). Among the genes that physically associate with the nuclear pore complex in yeast, we found an enrichment for genes with GRS I zip codes in their promoters.Citation22 Furthermore, the GRS I element, integrated at an ectopic locus, is sufficient to confer a physical interaction with the nuclear pore complex.Citation22 Therefore, we have argued that GRS-mediated targeting of genes to the nuclear periphery occurs through interaction of their promoters with the NPC.
We believe that current estimates for the number of genes that interact with the NPC and localize to the nuclear periphery may represent a conservative lower limit. The identification of NPC-associated genes by ChIP was performed only under a few conditionsCitation16,Citation17 and did not include many of the inducible genes that have been shown to be targeted to the nuclear periphery. We have identified ∼100 yeast genes with perfect GRS I elements in their promoters, but we imagine that there are a number of variant forms of this element that would expand this list. Additionally, two redundant zip codes, GRS I and GRS II, control the peripheral targeting of INO1. These elements are functionally redundant but are not similar DNA sequences. Therefore, some genes may be targeted by only GRS I (as is the case for the yeast TSA2 gene), some genes may be targeted by only GRS II and some genes may be targeted by redundant mechanisms.Citation22 Therefore, until we better understand the mechanisms controlling gene localization to the nuclear periphery, it is difficult to know how broadly these mechanisms are employed.
Two redundant DNA zip codes target transcriptionally active INO1 to the nuclear pore complex.Citation22 When INO1 is repressed, the gene does not immediately return to the nucleoplasm. INO1 remains at the nuclear periphery after being repressed, a phenomenon known as transcriptional memory.Citation21 While at the nuclear periphery, genes in the “memory state” are primed for reactivation. Retention of INO1 at the nuclear periphery is maintained through 3 to 4 cell division and is observed in the daughters, grand daughters and great-grand daughters of the cells that previously expressed the gene.Citation21 This inherited, epigenetic retention of INO1 at the nuclear periphery involves a different targeting mechanism than the active gene, requiring the histone variant H2A.ZCitation21 and a distinct interaction with the NPC (Light et al. submitted). Furthermore, whereas targeting of active INO1 promotes a higher level of transcriptionCitation22 (see below), retention of genes after repression promotes a faster rate of reactivation.Citation21 Therefore there are several (and perhaps many) mechanisms by which genes can interact with the NPC and localize to the nuclear periphery and some genes use more than one.
Peripheral Localization and Gene Expression
Both active and repressed loci localize at the nuclear periphery and localization affects these expression states.Citation32 Tethering of telomeres to the nuclear envelope plays a role in the establishment of transcriptional silencing of subtelomeric genes in yeastCitation33 and the tethering of genes to the nuclear lamina in mammals is often sufficient to promote repression ().Citation34 This has led to the suggestion that the nuclear periphery is a complex environment with sub-compartments that allow active and silenced regions of the genome to be in close proximity. Consistent with this proposal, interaction with the NPC is sufficient to establish boundaries between these silenced and active chromatin domains.Citation35,Citation36
Genes that relocalize to the nuclear periphery upon activation require targeting for full expression. Mutation of the GRS zip codes in the INO1 and TSA2 gene promoters causes a loss of peripheral localization and poor transcription.Citation22 Transcription correlates with gene relocalization and reaches full levels once targeting to the periphery is complete.Citation21,Citation37,Citation38 Artificially tethering of INO1 to the nuclear periphery enhances the rate of transcriptional activationCitation21 and tethering of the HXK1 gene to the nuclear periphery promotes more robust transcription.Citation38 Likewise, tethering nucleoporins of the Nup84 subcomplex to reporter genes leads to constitutive activation, suggesting that localization at the NPC is sufficient to promote transcription.Citation39 The hsp70 gene cluster localizes at the nuclear periphery, interacts with the NPC and expression of these genes depends on proteins that interact with the NPC.Citation23 The localization of the male X chromosome in Drosophila to the nuclear periphery involves interaction with the NPC and this is important for interaction with MSL dosage compensation factors, suggesting a correlation between peripheral localization and increased transcription.Citation25 Thus many genes require localization to the nuclear periphery for full activation.
Simply targeting genes to the nuclear periphery is usually not sufficient for full transcription. For the yeast genes we have examined, a specific interaction of the gene promoter with the NPC is important to promote transcription.Citation18,Citation22 Using a version of the INO1 gene where the GRS I sequence had been mutated, we were able to partially rescue the defect in transcription by reintroduction of the GRS I 5′ of the gene. Introduction of the GRS I sequence 3′ of the gene led to peripheral targeting but did not improve transcription.Citation22 This suggests that the interaction of the promoter with the NPC is important for transcription. Both GRS elements that have been studied in detail have been found in gene promoters (INO1 and TSA2). In fact, the positioning of the GRS I elements with respect to their transcriptional start sites is very similar in these two promoters. Genes co-regulated with INO1 most often contain GRS I elements located within 775 base pairs upstream of the open reading frame, suggesting that the positioning of the sequence may influence transcription of the gene.Citation22
During development, many genes move away from the nuclear periphery and towards the interior of the nucleus when induced (). Repositioning of the activated gene towards the nuclear interior has been documented for IgH (immunoglobulin heavy chain) in committed B lymphocytes,Citation4 Mash1 in neuronal cells,Citation6 β-globin in erythocytesCitation7 CFTR in adenocarcinoma cellsCitation5 and several tissue-specific promoters in C. elegans.Citation40 A progressive increase in expression of these genes is seen as they move away from the nuclear periphery.Citation7 Therefore, even though the nuclear periphery serves as a platform for increased gene expression in yeast, developmentally regulated genes seem to move to more interior positions, perhaps to interact with ‘transcription factories’. This change might also reflect escape from lamin-mediated repression ().Citation34 Furthermore, recent studies in Drosophila have demonstrated that NPC proteins interact with active genes both at the nuclear periphery and in the nucleoplasm ().Citation29–Citation31 The interaction with these nucleoporins is required for proper expression of many of these genes. Therefore, movement of some genes to a specific nuclear landmark may not be required to interact with proteins that are concentrated at the NPC.
In yeast, genes containing GRS I elements are transcriptionally co-regulated. Many of these genes are induced by heat shock and ER stress conditions.Citation22 This raises the interesting possibility that localization to the nuclear periphery in yeast may serve as a meeting point for coordinately expressed genes to colocalize and collectively benefit from the peripheral environment. This resembles a phenomenon known as ‘gene kissing’ where two or more loci on different chromosomes spatially colocalize with each other and with RNA polymerase II foci in the nucleus upon co-induction ().Citation41 Though ‘gene kissing’ has not been documented in yeast, it has been reported in T lymphocytes and erythroid cells. Using chromosome conformation capture (3C) the interferon-γ gene (Ifng) on mouse chromosome 10 was shown to physically interact with the TH2 cytokine gene locus located on chromosome 11 in naive T lymphocytes.Citation8 Furthermore, mutations at the chromosome 11 locus that led to a loss of kissing affected the expression of both genes following stimulation of naïve T lymphocytes.Citation8 Similarly, the active globin genes in erythroid cells interact with many co-regulated genes in transcription factories.Citation41 Consistent with the idea that DNA elements are important for regulating gene positioning, the colocalization of the globin genes with coregulated genes requires a transcription factor responsible for their induction ().Citation41 Therefore, it is possible that genes not only relocalize within the nucleus, but that there are mechanisms to bring together co-regulated genes either at an existing subnuclear body or as a way to create a new subnuclear compartment.
Why is the nuclear periphery in yeast beneficial for transcription of GRS I targeted genes? The simplest possibility is that the periphery presents a high local concentration of transcription factors, initiation factors and/or chromatin modifying enzymes important for the transcription of these genes. This is perhaps analogous to the high concentration of the Sir proteins at the nuclear envelope that help maintain and propagate silenced regions of the genome at the nuclear periphery.Citation42–Citation44 Previous work has shown that components of the SAGA histone acetyltransferase complex are required for the peripheral localization of the INO1 and GAL1 genes.Citation21,Citation45–Citation47 The SAGA complex has been suggested to physically associate with the nuclear pore complex through a bridging interaction between the Sac3 and Sus1 proteins.Citation48 Acetylation by the catalytic subunit of SAGA, the Gcn5 protein, promotes transcription initiation of the INO1 and GAL1 genes, both of which are targeted to the nuclear periphery upon activation.Citation49,Citation50 Therefore, localization to the nuclear periphery may help overcome rate-limiting steps in transcription initiation by providing increased access to SAGA components. Though this is an attractive model for the INO1 and GAL1 genes, it does not explain the role of NPC interaction in the transcription of TSA2, a gene not regulated at the level of chromatin. Therefore, localization to the nuclear periphery may fulfill a more basic requirement for transcription such as the binding of a transcription factor, the rate at which RNA polymerase II is recruited to the promoter or the rate at which RNA polymerase is recycled. Peripheral association may function at multiple steps to promote transcription initiation, elongation, termination or even post-transcriptional mRNA processing. Proximity to the nuclear pore complex may also allow improved mRNA export through a mechanism reminiscent of the ‘gene gating’ hypothesis.Citation51
Conclusions
Studies of nuclear architecture have long suggested the importance of three-dimensional positioning in establishing and maintaining gene expression states. Nuclear positioning of DNA has also been implicated in DNA repair and maintenance of genome stability, cancer and aging.Citation52 Therefore, to better understand the regulation of genomes and cellular changes that result as a consequence of natural development or aging, it is important to understand where loci are positioned in the nucleus. The GRS I element is able to function in an organism approximately one billion years diverged from the budding yeast Saccharomyces cerevisiae in which it was discovered.Citation22 Therefore, movement of DNA to the nuclear periphery represents an ancient mechanism that could be shared by other eukaryotic organisms. Understanding the fundamental principles that govern DNA movement and localization in the simple yeast model system may lay the foundation for the study of how genes and chromosomes are localized in more complex metazoan nuclei. The contribution of DNA sequence to gene positioning is only half the puzzle. The challenge that lies ahead is to identify proteins that recognize these sequences to cause the dynamic behavior of genes. Insight into the mechanism of gene targeting may also help highlight how localization of genes to particular nuclear environments impacts gene expression and how this is managed at the level of the genome.
Abbreviations
| SARs | = | scaffold associated regions |
| LAD | = | lamin associated domains |
| NPC | = | nuclear pore complex |
| GRS | = | gene recruitment sequence |
| TSA | = | trichostatin A |
| SAGA | = | Spt5-Ada2-Gcn5 acetyltransferase |
Figures and Tables
Figure 1 Gene relocalization within the nucleus associated with changes in expression. (A) Genes in yeast, and perhaps other organisms, associate with the nuclear pore complex upon activation. Interaction of the INO1 and TSA2 genes with the NPC requires DNA zip codes in their promoters and promotes their expression, perhaps because of a high local concentration of factors that promote transcription (yellow cloud). (B) Genes that are induced during development often relocalize away from the nuclear periphery upon activation. Interaction with the nuclear lamina is associated with transcriptional repression (red cloud). It is possible that cis-acting DNA elements within such genes (blue box) might either target them to the nuclear lamina when repressed or target them to an internal site when activated. (C) Some Drosophila genes interact with NPC proteins such as Nup153, Nup98, Sec13 and Mtor in the nucleoplasm and this promotes their expression. It is possible that cis-acting DNA elements (green box) control the interaction with NPC proteins specifically within the nucleoplasm. (D) Co-regulated genes from different chromosomes can cluster together upon activation, a phenomenon known as “gene kissing”. These genes often relocalize from within chromosome territories to the inter-chromosomal space. Colocalization of genes at “transcription factories” requires transcriptional regulators (grey ovals) that bind to cis-acting DNA elements (red boxes).
Acknowledgements
The authors are supported by the Searle Leadership Fund at Northwestern University, National Institutes of Health grant GM080484 and a W.M. Keck Young Scholar in Medical Research Award.
Extra View to:
References
- Rabl C. Gegenbaur C. über Zeillteilung. Morphol Jahrbuch 1885; 214 - 330
- Boveri T. Die blastermerenkerne von Ascaris megalocephala und die theorie der chromosomenindividualität. Arch Zellforschung 1909; 3:81
- Cremer T, Cremer M, Dietzel S, Muller S, Solovei I, Fakan S. Chromosome territories—a functional nuclear landscape. Curr Opin Cell Biol 2006; 18:307 - 316
- Kosak ST, Skok JA, Medina KL, Riblet R, Le Beau MM, Fisher AG, et al. Subnuclear compartmentalization of immunoglobulin loci during lymphocyte development. Science 2002; 296:158 - 162
- Zink D, Amaral MD, Englmann A, Lang S, Clarke LA, Rudolph C, et al. Transcription-dependent spatial arrangements of CFTR and adjacent genes in human cell nuclei. J Cell Biol 2004; 166:815 - 825
- Williams RR, Azuara V, Perry P, Sauer S, Dvorkina M, Jorgensen H, et al. Neural induction promotes large-scale chromatin reorganisation of the Mash1 locus. J Cell Sci 2006; 119:132 - 140
- Ragoczy T, Bender MA, Telling A, Byron R, Groudine M. The locus control region is required for association of the murine beta-globin locus with engaged transcription factories during erythroid maturation. Genes Dev 2006; 20:1447 - 1457
- Lee GR, Spilianakis CG, Flavell RA. Hypersensitive site 7 of the TH2 locus control region is essential for expressing TH2 cytokine genes and for long-range intrachromosomal interactions. Nat Immunol 2005; 6:42 - 48
- Misteli T. Beyond the sequence: cellular organization of genome function. Cell 2007; 128:787 - 800
- Spector DL. SnapShot: Cellular bodies. Cell 2006; 127:1071
- Takizawa T, Meaburn KJ, Misteli T. The meaning of gene positioning. Cell 2008; 135:9 - 13
- Paulson JR, Laemmli UK. The structure of histone-depleted metaphase chromosomes. Cell 1977; 12:817 - 828
- Amati B, Gasser SM. Drosophila scaffold-attached regions bind nuclear scaffolds and can function as ARS elements in both budding and fission yeasts. Mol Cell Biol 1990; 10:5442 - 5454
- Marshall WF, Dernburg AF, Harmon B, Agard DA, Sedat JW. Specific interactions of chromatin with the nuclear envelope: positional determination within the nucleus in Drosophila melanogaster. Mol Biol Cell 1996; 7:825 - 842
- Guelen L, Pagie L, Brasset E, Meuleman W, Faza MB, Talhout W, et al. Domain organization of human chromosomes revealed by mapping of nuclear lamina interactions. Nature 2008; 453:948 - 951
- Casolari JM, Brown CR, Drubin DA, Rando OJ, Silver PA. Developmentally induced changes in transcriptional program alter spatial organization across chromosomes. Genes Dev 2005; 19:1188 - 1198
- Casolari JM, Brown CR, Komili S, West J, Hieronymus H, Silver PA. Genome-wide localization of the nuclear transport machinery couples transcriptional status and nuclear organization. Cell 2004; 117:427 - 439
- Brickner JH, Walter P. Gene recruitment of the activated INO1 locus to the nuclear membrane. PLoS Biol 2004; 2:342
- Dieppois G, Iglesias N, Stutz F. Cotranscriptional recruitment to the mRNA export receptor Mex67p contributes to nuclear pore anchoring of activated genes. Mol Cell Biol 2006; 26:7858 - 7870
- Sarma NJ, Haley TM, Barbara KE, Buford TD, Willis KA, Santangelo GM. Glucose-responsive regulators of gene expression in Saccharomyces cerevisiae function at the nuclear periphery via a reverse recruitment mechanism. Genetics 2007; 175:1127 - 1135
- Brickner DG, Cajigas IC, Fondufe-Mittendorf Y, Ahmed S, Lee PC, Widom J, et al. H2A.Z-mediated localization of genes at the nuclear periphery confers epigenetic memory of previous transcriptional state. PLoS Biol 2007; 5:e81
- Ahmed S, Brickner DG, Light WH, McDonough M, Froyshteter AB, Volpe T, et al. DNA zip codes control an ancient mechanism for targeting genes to the nuclear periphery. Nat Cell Biol 2010; 12:111 - 118
- Kurshakova MM, Krasnov AN, Kopytova DV, Shidlovskii YV, Nikolenko JV, Nabirochkina EN, et al. SAGA and a novel Drosophila export complex anchor efficient transcription and mRNA export to NPC. EMBO J 2007; 26:4956 - 4965
- Brown CR, Kennedy CJ, Delmar VA, Forbes DJ, Silver PA. Global histone acetylation induces functional genomic reorganization at mammalian nuclear pore complexes. Genes Dev 2008; 22:627 - 639
- Mendjan S, Taipale M, Kind J, Holz H, Gebhardt P, Schelder M, et al. Nuclear pore components are involved in the transcriptional regulation of dosage compensation in Drosophila. Mol Cell 2006; 21:811 - 823
- Dernburg AF, Sedat JW. Mapping three-dimensional chromosome architecture in situ. Methods Cell Biol 1998; 53:187 - 233
- Gotta M, Laroche T, Gasser SM. Analysis of nuclear organization in Saccharomyces cerevisiae. Methods Enzymol 1999; 304:663 - 672
- Belmont AS. Visualizing chromosome dynamics with GFP. Trends Cell Biol 2001; 11:250 - 257
- Capelson M, Liang Y, Schulte R, Mair W, Wagner U, Hetzer MW. Chromatin-bound nuclear pore components regulate gene expression in higher eukaryotes. Cell 2010; 140:372 - 383
- Vaquerizas JM, Suyama R, Kind J, Miura K, Luscombe NM, Akhtar A. Nuclear pore proteins nup153 and megator define transcriptionally active regions in the Drosophila genome. PLoS Genet 2010; 6:1000846
- Kalverda B, Pickersgill H, Shloma VV, Fornerod M. Nucleoporins directly stimulate expression of developmental and cell cycle genes inside the nucleoplasm. Cell 2010; 140:360 - 371
- Akhtar A, Gasser SM. The nuclear envelope and transcriptional control. Nat Rev Genet 2007; 8:507 - 517
- Andrulis ED, Neiman AM, Zappulla DC, Sternglanz R. Perinuclear localization of chromatin facilitates transcriptional silencing. Nature 1998; 394:592 - 595
- Reddy KL, Zullo JM, Bertolino E, Singh H. Transcriptional repression mediated by repositioning of genes to the nuclear lamina. Nature 2008; 452:243 - 247
- Ishii K, Arib G, Lin C, Van Houwe G, Laemmli UK. Chromatin boundaries in budding yeast: the nuclear pore connection. Cell 2002; 109:551 - 562
- Dilworth DJ, Suprapto A, Padovan JC, Chait BT, Wozniak RW, Rout MP, et al. Nup2p dynamically associates with the distal regions of the yeast nuclear pore complex. J Cell Biol 2001; 153:1465 - 1478
- Schmid M, Arib G, Laemmli C, Nishikawa J, Durussel T, Laemmli UK. Nup-PI: the nucleopore-promoter interaction of genes in yeast. Mol Cell 2006; 21:379 - 391
- Taddei A, Van Houwe G, Hediger F, Kalck V, Cubizolles F, Schober H, et al. Nuclear pore association confers optimal expression levels for an inducible yeast gene. Nature 2006; 441:774 - 778
- Menon BB, Sarma NJ, Pasula S, Deminoff SJ, Willis KA, Barbara KE, et al. Reverse recruitment: the Nup84 nuclear pore subcomplex mediates Rap1/Gcr1/Gcr2 transcriptional activation. Proc Natl Acad Sci USA 2005; 102:5749 - 5754
- Meister P, Towbin BD, Pike BL, Ponti A, Gasser SM. The spatial dynamics of tissue-specific promoters during C. elegans development. Genes Dev 2010; 24:766 - 782
- Schoenfelder S, Sexton T, Chakalova L, Cope NF, Horton A, Andrews S, et al. Preferential associations between co-regulated genes reveal a transcriptional interactome in erythroid cells. Nat Genet 2010; 42:53 - 61
- Gotta M, Gasser SM. Nuclear organization and transcriptional silencing in yeast. Experientia 1996; 52:1136 - 1147
- Maillet L, Boscheron C, Gotta M, Marcand S, Gilson E, Gasser SM. Evidence for silencing compartments within the yeast nucleus: a role for telomere proximity and Sir protein concentration in silencer-mediated repression. Genes Dev 1996; 10:1796 - 1811
- Taddei A, Van Houwe G, Nagai S, Erb I, van Nimwegen E, Gasser SM. The functional importance of telomere clustering: global changes in gene expression result from SIR factor dispersion. Genome Res 2009; 19:611 - 625
- Cabal GG, Genovesio A, Rodriguez-Navarro S, Zimmer C, Gadal O, Lesne A, et al. SAGA interacting factors confine sub-diffusion of transcribed genes to the nuclear envelope. Nature 2006; 441:770 - 773
- Luthra R, Kerr SC, Harreman MT, Apponi LH, Fasken MB, Ramineni S, et al. Actively transcribed GAL genes can be physically linked to the nuclear pore by the SAGA chromatin modifying complex. J Biol Chem 2007; 282:3042 - 3049
- Chekanova JA, Abruzzi KC, Rosbash M, Belostotsky DA. Sus1, Sac3 and Thp1 mediate post-transcriptional tethering of active genes to the nuclear rim as well as to non-nascent mRNP. RNA 2008; 14:66 - 77
- Rodriguez-Navarro S, Fischer T, Luo MJ, Antunez O, Brettschneider S, Lechner J, et al. Sus1, a functional component of the SAGA histone acetylase complex and the nuclear pore-associated mRNA export machinery. Cell 2004; 116:75 - 86
- Bhaumik SR, Green MR. Differential requirement of SAGA components for recruitment of TATA-box-binding protein to promoters in vivo. Mol Cell Biol 2002; 22:7365 - 7371
- Dietz M, Heyken WT, Hoppen J, Geburtig S, Schuller HJ. TFIIB and subunits of the SAGA complex are involved in transcriptional activation of phospholipid biosynthetic genes by the regulatory protein Ino2 in the yeast Saccharomyces cerevisiae. Mol Microbiol 2003; 48:1119 - 1130
- Blobel G. Gene gating: a hypothesis. Proc Natl Acad Sci USA 1985; 82:8527 - 8529
- Misteli T, Soutoglou E. The emerging role of nuclear architecture in DNA repair and genome maintenance. Nat Rev Mol Cell Biol 2009; 10:243 - 254