Abstract
5-Methylcytosine (5-mC) is an epigenetic modification associated with gene repression. Recent studies demonstrated that 5-mC can be enzymatically oxidised into 5-hydroxymethylcytosine and further into 5-formylcytosine (5-fC) and 5-carboxylcytsine (5-caC). 5-caC has been found in embryonic stem cells and in mouse pre-implantation embryos but no detectable levels of this modification have been reported for somatic tissues to date. Whereas it has been suggested that 5-caC can serve as an intermediate in the process of active demethylation, the function of this form of modified cytosine remains obscure. Here we show that 5-caC is immunochemically detectable in somatic cells of axolotl ovary. We demonstrate that both 5-hmC and 5-caC are localized to the euchromatin in the nuclei of axolotl follicular cells with similar patterns of spatial distribution. Our results suggest that 5-carboxylcytosine may play a distinct functional role in certain biological contexts.
Results and Discussion
DNA methylation is an epigenetic modification of vertebrate genomes linked with a repressive chromatin state.Citation1,Citation2 Tet (Ten-11 translocation) proteins (Tet1/2/3) can enzymatically oxidise 5-methylcytosine (5-mC) generating another stable cytosine modification, 5-hydroxymethylcytosine (5-hmC), which differs from 5-mC in its genomic and tissue distribution as well as (likely) biological function.Citation3-Citation6 Recent reports demonstrated that 5-hmC can by oxidised further forming 5-formylcytosine (5-fC) and 5-carboxylcytosine (5-caC).Citation7,Citation8 Since both 5-fC and 5-caC can be recognized and removed from the DNA by the components of base excision repair machinery, a model according to which these oxidised forms of 5-methylcytosine may serve as intermediates in active demethylation process has been suggestedCitation8 but it is still unclear if this model is being employed during vertebrate development and differentiation. Thus, it has been reported that 5-methylcytosine is converted to 5-fC and 5-caC in mouse zygotes and both these modifications exhibit replication-dependent dilution during mouse pre-implantation development, which suggests that they are relatively stable and may play distinct biological roles in early mouse embryos.Citation9 Although significant levels of both 5-fC and 5-caC have been found in ESCs,Citation7,Citation8 no detectable levels of 5-carboxylcytosine have been reported for mouse somatic tissues.
Here we assessed the tissue distribution of 5-carboxylcytosine in adult amphibian (axolotl, Ambystoma mexicanum) tissues and demonstrate that 5-caC is immunochemically detectable in the somatic cells of axolotl ovary and localized to euchromatin in their nuclei.
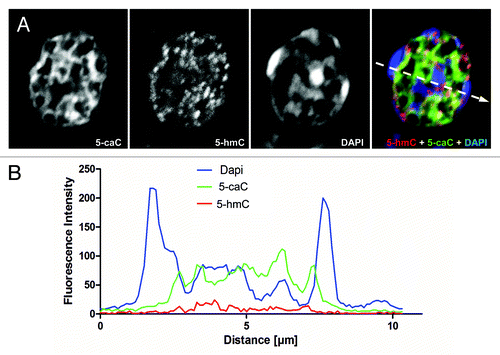
We previously showed that tissue distribution of 5-hydroxymethylcytosine is conserved between amphibians and mammals.Citation10 Although it can be detected in most of the adult axolotl organs, neuronal cells are significantly enriched with 5-hmC compared with the rest of tissues in amphibians.Citation10 Employing the same highly sensitive immunochemical staining procedure, based on the use of peroxidase-conjugated secondary antibody coupled with a tyramide signal enhancement system, we performed co-immunostaining of 5-caC and 5-hmC on the tissues of young axolotl adults (). Using-5-caC antibody (active motif), which is able to specifically recognize 5-caC but not 5-mC, 5-hmC or 5-fC according to the latest report,Citation9 we could not detect any 5-caC signal in axolotl neural tube, muscle or connective tissue ( and data not shown) . On the other hand, we detected specific 5-carboxylcytosine staining evident in mouse ESCs, and showed that this is eliminated in the presence of 5-caC- but not 5-mC-, 5-hmC-containing or unmodified DNA (Fig. S1), confirming antibody specificity. Similar to our previous results, the immunostaining using mouse monoclonal anti-5-hmC antibody produced non-negligible signal in all the tested tissues (). These results are in line with the previously published data on biochemical detection of 5-caC, where this modification was detectable in mouse ESCs but not in the adult mouse tissues.Citation7,Citation8 In contrast the immunostaining of ovarian tissue revealed relatively strong signal for both 5-hmC and 5-caC (). Notably, most of the somatic cells in developing follicles and cell nests, but not the surrounding cells of ovarian theca, exhibited strong 5-caC staining (). As we found that 5-caC is detectable in follicular cells of axolotl we decided to examine if the nuclear distribution of this mark matches that of 5-hydroxymethylcytosine in these cells. Unlike 5-methylcytosine, usually enriched in condensed DAPI-bright transcriptionally-silent chromatin, 5-hydroxymethylcytosine has been reported to be predominantly associated with euchromatic regions such as gene bodies and enhancer elements.Citation4 Since the anti-5-mC antibody we routinely use is mouse monoclonal (clone 33D3) and, therefore, it is technically impossible to co-localize all three epigenetic marks (5-mC, 5-hmC and 5-caC) on the same sample, we co-stained ovarian tissue with 5-hmC and 5-caC antibodies in mild denaturing conditions (2 M HCl), allowing us to visualize dense heterochromatic DNA regions with DAPI. Analysis of these immunostaining experiments, imaged using confocal microscopy, revealed that both 5-hmC and 5-caC are predominantly localized to gene-rich euchromatic regions but not to the DAPI-bright regions of condensed chromatin (; ). Quantification of the immunostaining showed that whereas the 2.5XD plots for 5-hmC and 5-caC signals exhibit similar profiles (), they both are strikingly different from that of DAPI staining (). Indeed, the signal profiling across the heterochromatic and euchromatic regions of the nucleus demonstrated that 5-carboxylcytosine signal intensity is highest in the regions of low or moderate DAPI signal, while the intensities of both 5-hmC and 5-caC signals are very weak in heterochromatic areas with strong DAPI staining (). At the same time, although both 5-hmC and 5-caC are localized to the euchromatin, their signal profiles are not absolutely identical (). Thus 5-hmC staining seems to be more punctate than that of 5-caC and certain 5-hydroxymethylcytosine enriched regions do not exhibit enrichment similar to the 5-caC content ().
Figure 1. 5-Carboxylcytosine is immunochemically detectable in the somatic cells of axolotl ovary. (A) 5-hmC and 5-caC immunostaining of young adult axolotl ovary and neural tube tissue (indicated). Developing follicle is marked with white and cell-nest with yellow arrows. (B) Immunohistochemical detection of 5-hmC and 5-caC in axolotl ovary and surrounding tissues. Immunostaining for 5-hmC, 5-caC, DAPI and 5-hmC/5-caC merge views are shown. The morphological border of ovarian theca is marked with a dotted line. Examples of non-specific (non-nuclear) staining produced by anti-5-caC antibody in ovarian theca are marked with white arrow. Scale bars are 20 μm.
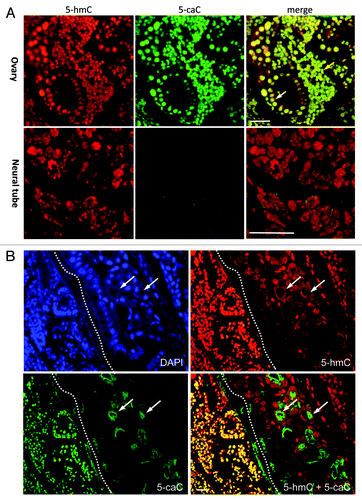
Figure 2. Both 5-hydroxymethylcytosine and 5-carboxylcytosine are distributed to the euchromatic regions in the nuclei of axolotl follicular cells. (A) Confocal image of a representative field of axolotl ovary stained with 5-hmC, 5-caC and DAPI. Merge view is shown. An inset shows the cells used for the analysis presented in (B-E) under higher magnification. Scale bar is 10 μm. (B-E) 2.5XD plot of 5-hmC, 5-caC and DAPI signal intensities of the inset in (A). Merge view (B) and single channel images (C, D and E) are shown.
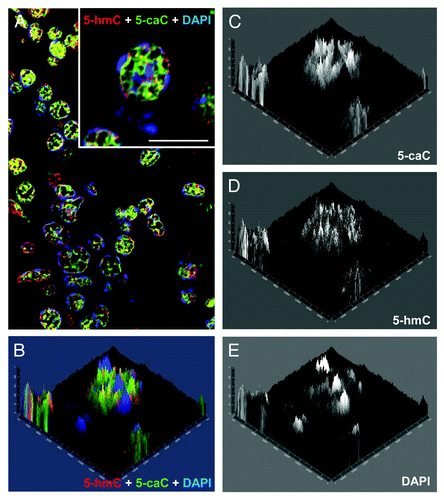
Figure 3. The patterns of nuclear distribution of 5-hydroxymethylcytosine and 5-carboxylcytosine are similar but not identical. (A) Immunochemical detection of 5-hmC and 5-caC in a single nucleus of axolotl follicular cell co-stained with DAPI using confocal microscopy. Merge view and single channel images (indicated) are shown. The position of the region used for the generation of the signal intensities profile plot shown in (B) is marked with a dashed arrow. (B) The profile of DAPI, 5-hmC and 5-caC signal intensities (indicated) across heterochromatic and euchromatic regions of the follicular cell nucleus presented in (A).
Our data show that 5-carboxylcytosine is immunochemically detectable in the somatic cells of amphibian ovary, where both 5-hmC and 5-caC are localized mainly to gene-rich euchromatic regions with similar patterns of nuclear distribution. As the oxidation of 5-hydroxymethylcytosine to 5-carboxylcytosine is taking place in mammalian zygotesCitation9 where Tet3 protein is likely responsible for this conversion,Citation11 it would be interesting to check if a Tet3 homolog is required for the generation of 5-caC in amphibian follicular cells. Since, according to our data, most of the somatic cells in the ovary possess similar levels of 5-carboxylcytosine, were this modification to indicate the presence of active demethylation, our results would imply an extremely dynamic demethylation process, with constant cycles of 5-methylcytosine removal from DNA followed by re-methylation. Such genome wide demethylation taking place at the actively transcribed euchromatic regions of follicular cells’ DNA would most likely contribute to switching on and off the transcription of a number of genes active at different phases of amphibian oocyte development.
Since according to the recent reports, 5-caC reduces the rate and substrate specificity of RNA polymerase II transcription,Citation12 and 5-formylcytosine distribution is associated with transcription in mouse ESCs,Citation13 an alternative explanation of our results would suggest that 5-carboxylcytosine is not always an intermediate in demethylation but may instead play a distinct functional role in transcription regulation in some biological contexts.
Materials and Methods
ES cell culture
Mouse HM1 ESCs were maintained on gelatine-coated dishes in Glasgow's minimum essential medium (GIBCO), supplemented with 15% fetal bovine serum, 55 μM β-mercaptoethanol (GIBCO), 2 mM l-glutamine, 0.1 mM MEM non-essential amino acids, 5 000 units ml−1 penicillin/streptomycin and 1 000 units ml−1 of murine LIF (Chemicon) under feeder-free conditions.
Immunohistochemistry and imaging
Paraffin embedded formaldehyde fixed sections of axolotl adult tissues were used for immunohistochemistry. Tissues were fixed in 4% formaldehyde for 12 h. Tissue sections were de-waxed according to standard procedures and permeabilised for 15 min with PBS containing 0.5% Triton X-100. For 5-hmC and 5-mC staining, permeabilised tissue sections were incubated in 2 N HCl for 1 h at 37°C and then neutralised in 100 mM TRIS-HCl (pH 8.5) for 10 min, followed by a standard immunostaining protocol. Anti-5-hmC mouse monoclonal (Active Motif, 1:5000 dilution) and anti-5-caC rabbit polyclonal (Active Motif, 1:3000 dilution) primary antibodies were used. Peroxidase-conjugated anti-rabbit secondary antibody (Dako) and the tyramide signal enhancement system (Perkin Elmer) were employed for 5-caC detection. 5-hmC was visualized using 555-conjugated secondary antibody (Alexafluor). Control staining without primary antibody produced no detectable signal. Images were acquired using a Nikon ECLIPSE 90i immunofluorescence microscope and Volocity software.
Immunostaining competition experiment
Immunostaining competition experiment was performed as describedCitation6 using 5-caC DNA standard (Active Motif) and PCR-produced 100-bp DNA fragments, where all the cytosines were replaced with either 5-hmC or 5-mC. The sequence of the PCR fragment used is available upon request.
Confocal microscopy
Images (400 nm optical sections) were acquired with a Zeiss LSM 700 AxioObserver confocal microscope using a Plan-Apochromat 63×/1.40 Oil DIC M27 objective. Images were then processed using Image J and Adobe Photoshop software. 2.5xD images were generated using Zeiss LSM 700 imaging software.
Additional material
Download Zip (50.5 KB)Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
We thank Lorrain Young (University of Nottingham) for support. We would like to acknowledge Denise Christie, Advanced Microscopy Unit, School of Biomedical Sciences, University of Nottingham for her contribution to the preparation of histological samples used in this study.
References
- Bird A. DNA methylation patterns and epigenetic memory. Genes Dev 2002; 16:6 - 21; http://dx.doi.org/10.1101/gad.947102; PMID: 11782440
- Reik W, Dean W, Walter J. Epigenetic reprogramming in mammalian development. Science 2001; 293:1089 - 93; http://dx.doi.org/10.1126/science.1063443; PMID: 11498579
- Ito S, D’Alessio AC, Taranova OV, Hong K, Sowers LC, Zhang Y. Role of Tet proteins in 5mC to 5hmC conversion, ES-cell self-renewal and inner cell mass specification. Nature 2010; 466:1129 - 33; http://dx.doi.org/10.1038/nature09303; PMID: 20639862
- Ficz G, Branco MR, Seisenberger S, Santos F, Krueger F, Hore TA, et al. Dynamic regulation of 5-hydroxymethylcytosine in mouse ES cells and during differentiation. Nature 2011; 473:398 - 402; http://dx.doi.org/10.1038/nature10008; PMID: 21460836
- Tahiliani M, Koh KP, Shen Y, Pastor WA, Bandukwala H, Brudno Y, et al. Conversion of 5-methylcytosine to 5-hydroxymethylcytosine in mammalian DNA by MLL partner TET1. Science 2009; 324:930 - 5; http://dx.doi.org/10.1126/science.1170116; PMID: 19372391
- Ruzov A, Tsenkina Y, Serio A, Dudnakova T, Fletcher J, Bai Y, et al. Lineage-specific distribution of high levels of genomic 5-hydroxymethylcytosine in mammalian development. Cell Res 2011; 21:1332 - 42; http://dx.doi.org/10.1038/cr.2011.113; PMID: 21747414
- Ito S, Shen L, Dai Q, Wu SC, Collins LB, Swenberg JA, et al. Tet proteins can convert 5-methylcytosine to 5-formylcytosine and 5-carboxylcytosine. Science 2011; 333:1300 - 3; http://dx.doi.org/10.1126/science.1210597; PMID: 21778364
- He YF, Li BZ, Li Z, Liu P, Wang Y, Tang Q, et al. Tet-mediated formation of 5-carboxylcytosine and its excision by TDG in mammalian DNA. Science 2011; 333:1303 - 7; http://dx.doi.org/10.1126/science.1210944; PMID: 21817016
- Inoue A, Shen L, Dai Q, He C, Zhang Y. Generation and replication-dependent dilution of 5fC and 5caC during mouse preimplantation development. Cell Res 2011; 21:1670 - 6; http://dx.doi.org/10.1038/cr.2011.189; PMID: 22124233
- Almeida RD, Sottile V, Loose M, De Sousa PA, Johnson AD, Ruzov A. Semi-quantitative immunohistochemical detection of 5-hydroxymethyl-cytosine reveals conservation of its tissue distribution between amphibians and mammals. Epigenetics 2012; 7:137 - 40; http://dx.doi.org/10.4161/epi.7.2.18949; PMID: 22395462
- Gu TP, Guo F, Yang H, Wu HP, Xu GF, Liu W, et al. The role of Tet3 DNA dioxygenase in epigenetic reprogramming by oocytes. Nature 2011; 477:606 - 10; http://dx.doi.org/10.1038/nature10443; PMID: 21892189
- Kellinger MW, Song CX, Chong J, Lu XY, He C, Wang D. 5-formylcytosine and 5-carboxylcytosine reduce the rate and substrate specificity of RNA polymerase II transcription. Nat Struct Mol Biol 2012; 19:831 - 3; http://dx.doi.org/10.1038/nsmb.2346; PMID: 22820989
- Raiber EA, Beraldi D, Ficz G, Burgess HE, Branco MR, Murat P, et al. Genome-wide distribution of 5-formylcytosine in embryonic stem cells is associated with transcription and depends on thymine DNA glycosylase. Genome Biol 2012; 13:R69; http://dx.doi.org/10.1186/gb-2012-13-8-r69; PMID: 22902005