
Abstract
Current anti-cancer therapies have a great deal of undesirable side effects; therefore, there is a need to develop efficient and cancer cell-specific new drugs without strong dose-limiting side effects. In my opinion, mechanisms of nuclear assembly and organization represent a novel platform for drug targets, which might fulfill these criteria. The nuclear stiffness and organization of some cancer types are often compromised, making them more vulnerable for further targeting the mechanisms of nuclear integrity than their normal counterparts. Here I will discuss the nuclear organization of normal cells and cancer cells, the molecular mechanisms that govern nuclear assembly with emphasis on those that, in my view, might be considered as targets for future anti-cancer therapies.
Pitfalls of Current Anti-Mitotic Drugs
Currently the most efficient anti-cancer chemotherapy agents are the cytotoxic drugs that target cancer cells, in a most vulnerable state, during mitosis.Citation1,Citation2 These include the microtubule-binding taxanes (e.g., paclitaxel) that stabilize and vinca alkaloids (e.g., vinblastine) that destabilize microtubule polymers.Citation3 Both promote abnormal spindle assembly, chromosome misalignment, consequent activation of the spindle assembly checkpoint (SAC), and induce prolonged mitotic arrest and mitotic cell death.Citation4 However, cancer cells often adapt to these drugs and exit mitosis in a process called mitotic slippage.Citation5 Moreover, these drugs target not only mitotic cells but also affect the microtubule cytoskeleton functions of non-proliferating cells. They disrupt the interphase microtubule bundles of quiescent neuronal cells, along which molecular and vesicular transport occurs. Neurotoxicity is therefore one of the most common dose-limiting side effects of microtubule-targeting drugs.Citation6 Hence, there is a need to develop new anti-mitotic drugs, which by targeting specific mitotic proteins and mechanisms would achieve a similar level of anti-cancer efficacy without the unwanted side effects on non-proliferating cells. Such new targets included mitotic kinases (e.g., PLK1, Aurora kinases) and mitotic motor proteins (e.g., Eg5, CENPE) required for spindle assembly, chromosome alignment, and segregation. Their inhibition also induces SAC-mediated mitotic arrest and leads to mitotic cell death.Citation1 Although these new anti-mitotic drugs are highly specific in vitro and had some efficacy in xenograft models, they were so far less convincing in clinical trials than the microtubule-targeting drugs.Citation7,Citation8 The reasons for these disappointing clinical results might be the low mitotic index (i.e., low number of cells undergoing division) of certain human tumorsCitation1,Citation7 and strong neutropenia, the main dose-limiting toxicity of the new anti-mitotic drugs.Citation7,Citation9 Neutropenia refers to an abnormally low number of neutrophil granulocytes, the most abundant type of white blood cells. The reason why this occurs as a side effect is that during their development neutrophils frequently divide. Anti-mitotic drugs are not cancer cell-specific; they target every dividing cell, including neutrophils, and thus neutropenia is a consequence of their activity. Future anti-cancer drugs should therefore possess a larger therapeutic window and exert a higher-level cancer cell-specificity. One strategy to achieve this goal might be to develop new drugs that are synthetic lethal with mechanisms of cancer cell-specific hallmarks. In my opinion such new drug targets are the molecular mechanisms of nuclear assembly and organization because some of these mechanisms might get compromised during tumorigenesis, making the nuclei of these cancer cells more vulnerable than their normal counterparts. Below I will discuss the nuclear organization of normal cells and cancer cells, the molecular mechanisms that govern nuclear assembly, and I will hypothesize about the usefulness of targeting some of these mechanisms in future anti-cancer therapies.
Nuclear Organization of Normal Cells
The nucleus contains the genome of the eukaryotic cell, whose precise organization is essential for normal cell function. The structure that defines the nucleus is the nuclear envelope (NE), a sub-domain of the endoplasmic reticulum (ER) ().Citation10,Citation11 The NE is composed of outer nuclear membranes (ONM) and inner nuclear membranes (INM) with different protein compositions, which are fused at the sites of nuclear pore complex (NPC) insertion. Underlying the INM is the nuclear lamina, which is mainly composed of intermediate lamin filaments. In vertebrates, lamin proteins are grouped into A-type lamins (lamin A, Δ10, and C) encoded by the LMNA gene and B-type lamins (lamin B1 and B2) encoded by LMNB1 and LMNB2 genes.Citation12,Citation13 While B-type lamins are expressed in every somatic cell, A-type lamins are absent from highly pluripotent and rapidly dividing cells and are present only in differentiated cells. Lamins are alternatively spliced, and the ratio between the expressions of individual lamin isoforms is characteristic of each cell type.Citation14,Citation15,Citation16 Lamins mainly form homodimers, which assemble into head-to-tail homopolymers. These polymers then laterally assemble into filaments, which form a lattice of remarkably regular arrangement underneath the INM.Citation17,Citation18 During maturation most lamins are C-terminally farnesylated. While this short lipid chain is retained on B-type lamins and anchors the proteins in the membranes, the farnesylated C-terminal part of lamin A is enzymatically cleaved after its incorporation into lamin polymers. The stabilization and organization of lamin filaments underneath the INM also requires lamin-associated membrane proteins, such as lamin B receptor (LBR), diverse LAP2/emerin/MAN1 (LEM) domain-containing proteins, and others.Citation12,Citation13
Figure 1. Nuclear organization. (A) The nuclear shape of most normal cells is oval. Nuclear envelope (yellow) is penetrated by the nuclear pore complexes (red). Underneath the nuclear envelope is the nuclear lamina (green), which provides the stiffness to the NE and serves as a tethering surface for chromosomes (blue lines). (B) Several cancer types are characterized by abnormal expression of lamins (particularly A-type), altered chromatin organization, and multi-lobulated nuclear shape. Other changes involve enlargement or fragmentation of the nucleolus (brown, large circles) and promyelocytic leukemia (PML) bodies (orange, small circles). Note that all these changes might not occur simultaneously in every cancer type. (C) Possible nuclear appearance of cancer cell nuclei after the treatment with drugs that interfere with mechanisms of nuclear assembly and organization.
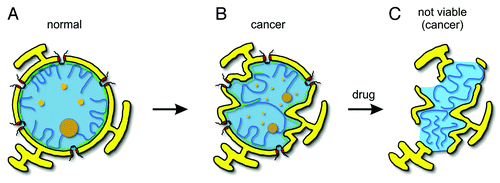
Nuclear lamina confers the shape, elasticity and the stiffness to the NE. The stoichiometry of A-type and B-type lamins correlates well with the mechanical stress the cells experience within a tissue.Citation16 In soft tissues such as liver or brain, A-type lamins are relatively low expressed, while in stiff tissues such as heart or muscle A-type lamins increase (up to 30-fold) to withstand the mechanical stress and to limit the potential disruption of the chromatin. B-type lamins are constitutively expressed and correlate much less with nuclear stiffness.Citation16,Citation19
By tethering the chromatin to the NE, nuclear lamina also contributes to the non-random chromatin organization within the nucleus (). Developmentally coregulated genes often form clusters on chromosomes, which are associated and corepressed at the nuclear periphery in cells where they are not expressed. Human chromosomes associate with the nuclear lamina via roughly 1000 sharply defined domains. These interactions are known to change progressively during differentiation.Citation20 Genes and chromosomal domains that become internalized during certain differentiation steps are either immediately activated or are unlocked and prepared for activation during further differentiation steps. This suggests that the nuclear lamina often provides a repressive environment for chromosomal domains at the nuclear periphery whose three-dimensional organization is specific for certain cell types and differentiation states. Mutation or downregulation of lamins or lamin-associated proteins results in disorganized lamin filamentsCitation21 and deformed, multi-lobulated, and fragile nuclei,Citation22,Citation23 which are often observed in human diseases such as cancer.Citation24-Citation26
Nuclear Organization is Disrupted in Cancer Cells
Cancer is initiated by genetic processes such as genome instability, genome rearrangements, or specific gene mutations, amplifications, or deletions, followed by epigenetic modifications, which ultimately lead to altered gene expression and result in deregulated cell proliferation. In cancer cells, nuclear size and shape are frequently altered.Citation24,Citation25 Cancer-related morphological changes include NE invaginations, multi-lobulation, malleable and passively distorted nuclei, and altered appearance of heterochromatin, nucleoli, and nuclear bodiesCitation27-Citation29 (). Although all these features do not occur simultaneously, they are often used individually in the clinical diagnosis of cancer, in the assessment of the degree of the malignancy, and for prognostic and predictive indications of the disease state.Citation27-Citation30
Lamins, the most important architectural elements of the nucleus, are often aberrantly expressed or localized in cancer cells, and it is likely that this contributes to the multi-lobulated nuclear shape often observed in different cancer typesCitation31 (). Many poorly differentiated cancer types exhibit downregulation of A-type lamins and concomitant irregularities in their nuclear shape. For example, in small cell lung cancer (SCLC) cells A-type lamins are either not or only weakly expressed, while in non-SCLC cells they are normally expressed but frequently mislocalized in the cytoplasm.Citation32-Citation34 In colon cancers,Citation35 gastric cancers,Citation36,Citation37 breast cancers,Citation38,Citation39 and diffuse large B-cell lymphomas,Citation40 the expression of A-type lamins is also strongly reduced, and this feature correlates with increased disease recurrence and poor patient prognosis. Consequently, the nuclei of these cancer types are often fragile and lobulated, and in breast cancer cells, they were shown to contain massive NE membrane invaginationsCitation27,Citation28 (). The nuclei of prostate cancer cellsCitation41 and of some other cancer typesCitation42 are not only lobulated but also contain structures called nuclear blebs, protrusions from the nuclear surface enriched in lamin A/C but deficient in lamin B. Further examples with multi-lobulated nuclear shape are ovarian cancers,Citation43 papillary thyroid cancers,Citation27 leukemias,Citation44 and different B-cell lymphomas.Citation40,Citation45,Citation46 In conclusion, altered lamin expression or localization and disrupted stoichiometry between A- and B-type lamins can change the elastic properties of the NE,Citation16 which renders it unable to withstand cytoskeleton-Citation47,Citation48 and chromosome-basedCitation49,Citation50 forces and leads to misshapen nuclei. Consistently, downregulation of lamin A/C in non-cancer primary breast epithelial cells results in nuclear alterations similar to those observed in breast cancer cell.Citation38 Moreover, mutations in LMNA and other genes encoding for proteins of nuclear lamina results in heritable diseases called laminopathies,Citation51 which are also characterized by fragileCitation52 and multi-lobulated nuclear shape.Citation53 Therefore the nuclear lamina alterations might directly account for the cancer-related changes in the nuclear morphology. However, it is important to note that several other cancer types display normal nuclear shape and that there is no simple universal pattern of lamin expressions for all cancer types.Citation31,Citation45,Citation46,Citation54
It is unclear whether the abnormal nuclear organization observed in cancer cells is the cause or the consequence of transformation and tumor progression. Lamins might modulate gene expression not only indirectly through influencing global chromatin organization but also by directly interacting with transcription factors that affect cellular proliferation, differentiation, and apoptosis.Citation55,Citation56,Citation57 Therefore, the absence of lamins from tumors derived from tissues where they are normally present led to the hypothesis that lamins might be directly involved in tumorigenesis. However, this tempting hypothesis is contradicted by several lines of evidence. LMNA gene contains the largest diversity of mutations that lead to rare human diseases. There are more than 20 distinct laminopathies associated with approximately 400 different mutations in the human LMNA gene.Citation13,Citation51 Most of these mutations affect the assembly, dynamics, or function of lamin filaments and result in deformed, multi-lobulated, and fragile nuclei and abnormal heterochromatin structure, similar to the nuclear abnormalities observed in cancer cells.Citation21,Citation58,Citation59,Citation60 However, none of the mutations in lamin or in lamin-organizing proteins are known to be tumorigenic, and patients with laminopathies are not more susceptible to cancer development than healthy individuals with normal nuclear architecture. Moreover, loss of lamin A from human fibroblast cellsCitation61 or ovary surface epithelial cellsCitation43 results in reduced mitosis and retarded cell growth, which at least in part might be explained by the active role of lamin A in nuclear localization of the cell cycle regulator retinoblastoma protein (Rb).Citation61 Finally, although higher order chromatin organization is commonly altered in cancer cells, this does not necessarily lead to tumorigenic transcriptional changes. In breast cancer for example, several genes have been identified that specifically change their nuclear position only in cancer cells. However, the absence of transcriptional changes associated with the movement of these genes suggests that these changes are not responsible for tumorigenesis.Citation62 In conclusion, although precise nuclear organization is essential for normal cellular function, up to now altered nuclear architecture has not been shown to drive cancer development, and therefore, it is more likely to be a consequence of cell transformation and tumor progression. Accordingly, nuclear irregularities arise dynamically during interphase following oncogene induction.Citation48,Citation63 Since the nuclear architecture of many cancer cells is abnormal, this hallmark might improve cancer cell-selectivity in therapies using mechanisms of mitotic nuclear assembly and nuclear organization as targets ().
Mitotic Nuclear Dynamics
The nuclear structure is disassembled and reassembled during every cell division to allow cytoplasmic spindle microtubules to segregate the duplicated sister chromatids. These dynamics are under precise spatial and temporal control of mitotic kinases and phosphatases.Citation64,Citation65,Citation66
Mitotic nuclear disassembly is controlled by cyclin-dependent kinase 1 (CDK1)Citation67,Citation68,Citation69 and other mitotic kinases that function downstream of its activation, such as protein kinase C (PKC),Citation70,Citation71 Aurora A,Citation72 polo-like kinase 1 (PLK1),Citation73 NIMA-related kinases,Citation68 and vaccinia-related kinase 1 (VRK1).Citation74,Citation75,Citation76 They phosphorylate proteins of nuclear lamina and NPC to disrupt their interphase molecular interactions. In support of their critical role, inactivation or inhibition of these mitotic kinases either blocks or delays various steps of nuclear disassembly. For example, VRK1 kinase is activated during mitotic entry upon degradation of its interphase inhibitor macroH2A1.Citation77 It relocalizes to the nuclear peripheryCitation75 and phosphorylate barrier-to-autointegration factor (BAF or BANF1) to release it from DNA and LEM domain-containing INM proteinsCitation75,Citation76 (). Inhibition of VRK1 thus prevents a late but essential step of nuclear disassembly. As a consequence of massive protein phosphorylation events, during NE breakdown soluble proteins become dispersed into the cytosol while membrane proteins become mobile and, together with the NE membranes, absorbed into the oscillating ER network.Citation78
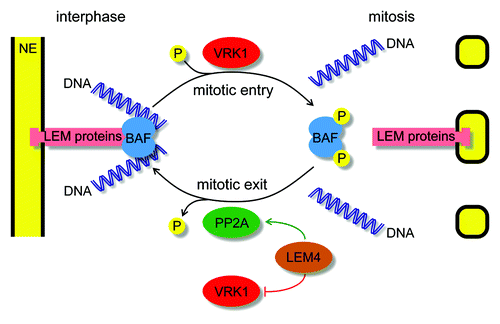
Figure 2. Mitotic regulation of BAF function. During interphase BAF binds as a dimer (light blue) to one LEM domain-containing integral nuclear envelope protein (pink) and to two DNA helices (dark blue). During mitotic entry VRK1 (red) phosphorylates BAF to disrupt its interactions and contributes to nuclear envelope breakdown. During mitotic exit LEM4 (brown) inhibits VRK1 (red) and promotes PP2A (green) to dephosphorylate BAF and to enable its function in post-mitotic nuclear reassembly.
CDK1 promotes mitotic progression until the alignment of mitotic chromosomes on the metaphase plate and the correct bipolar attachment of all the kinteochores with spindle poles is achieved. This turns off the SAC and promotes anaphase onset.Citation65,Citation66 Subsequently, members of the protein phosphatase 1 (PP1)Citation79 and 2A (PP2A)Citation74 family are activated. They counteract CDK1 and other mitotic kinases to allow the assembly of the interphase nucleus.Citation74,Citation79 Chromatin decondensation is an important step of post-mitotic nuclear assembly. Although its exact mechanism is still unclear, likely players are PP1, which acts by dephosphorylating histone H3,Citation79 AAA-ATPase p97, which extracts the polyubiquitylated Aurora B histone kinase from chromosomes,Citation80 and the small Ras-like GTPase Ran, which acts in a still poorly understood manner.
The first step of NE reformation is the attachment of ER membranesCitation81,Citation82,Citation83 to the chromatin surface. Although direct lipid-chromatin interactions might play a role, this interaction is mainly mediated by trans-membrane and membrane-associated proteins. A large population of INM proteins possesses a highly basic nucleoplasmic domain that can directly bind to DNA.Citation84 The INM proteins function redundantly, thus their individual inactivation has only a minor effect on NE assembly and even simultaneous inactivation of several only delays but does not prevent the recruitment of membranes to the chromatin surface.Citation75,Citation85,Citation86 Other INM proteins interact with chromatin through specific adaptor proteins. These interactions are controlled spatially by the small GTPase Ran and temporally by protein dephosphorylation. GTP-loaded Ran is generated in the vicinity of chromosomesCitation87 and mediates the release of the inhibitory importin receptors from their target proteins, providing spatial control of NE reformation.Citation11 As a specific example, during mitotic exit RanGTP releases importin-β from the chromatin-binding domain of LBR,Citation88,Citation89 thereby allowing its binding to histones H3/H4Citation90 and HP1.Citation91
In the subsequent steps, chromatin-attached membranes start to spread from the “peripheral” margins of the separating chromatin to surround the entire chromatin mass and enclose it in a single nuclear compartment. There is constant membrane supply from the ER, and manipulation of ER structure was shown to influence NE assembly.Citation70,Citation82,Citation92,Citation93 The recruited membranes on the chromatin surface are organized by BAF, which as a dimer can bind to one LEM domain of an INM protein and to two DNA helices ().Citation64,Citation94,Citation95 Consistently, inactivation of BAF results in deformed, multi-lobulated nuclei, with NE membranes trapped between individual chromosomes.Citation15,Citation75,Citation96 The localization and function of BAF is regulated by phosphorylation, which is temporally controlled by LEM4 ().Citation64 During mitotic exit LEM4 binds to VRK1, BAF’s mitotic kinase, and inhibits its further activity on BAF.Citation74 Concomitantly, LEM4 also binds to and activates a complex of PP2A (i.e., PP2A-B55α) to dephosphorylate BAF and to allow its re-association with chromatin and INM protein.Citation74 This particular PP2A complex has been further implicated in other mitotic functions and is the only protein phosphatase essential for mitotic exit.Citation97 Recently PP4C, which is known to form complexes with different PP2A subunits,Citation98,Citation99 was also suggested to influence the phosphorylation state of BAF.Citation100 Nevertheless, during mitotic exit BAF rapidly and strongly accumulates on the central surface of the chromatin “core” regions, which surround the anaphase chromatin mass, on one side facing the spindle microtubules and on the other facing the midzone microtubules.Citation75,Citation101 The “core” region is a thick, electrondense structure mainly devoid of membranes. One of the features of the “core” region is that it rapidly shrinks with mitotic progression prior to the spread of the NE membranes from the peripheral regions toward the central region.Citation75,Citation101 The exact function of BAF during NE membrane organization is still unclear; however, it may synchronize different membrane and chromatin events during NE assembly to enable the incorporation of the entire chromatin mass into a single nuclear compartment.Citation75
Concomitantly with NE reformation, NPCs are also assembled post-mitotically via mechanisms that are different from those used during interphase NPC assembly.Citation102 NPC sub-complexes form pre-pores on the chromatin surface, which are then incorporated in the NE during membrane spreading on the chromatin surface. In the final step of NE assembly, remaining holes in the NE membranes may be fused by SNARE proteins.Citation103 However, since these holes can also be occupied by NPCs, the membrane fusion machinery is less important during NE assembly than was initially anticipated. Upon the formation of a closed NE, nucleocytoplasmic transport is reactivated. The lamin proteins are actively imported into the nucleiCitation104 where they are polymerized and organized underneath the INM to provide shape, elasticity, and stiffness to the nuclei.
Synthetic Lethality with Abnormal Nuclear Architecture of Cancer Cells
In genetic studies using different experimental model organisms, synthetic lethality is based on the fact that inactivation of one gene makes the cell vulnerable for the inactivation of the other gene, while neither of these two genes is essential on its own. In anti-cancer therapies similar synthetic lethality might be achieved between cancer cells and drugs, in case the cancer cell-specific molecular lesions sensitize the cancer cells for drugs inhibiting particular protein functions. The value of such an approach is that normal cells without such lesions will be unaffected. As discussed earlier, the nuclear architecture of different cancer cells is compromised and unable to withstand cytoskeleton-Citation47,Citation48 and chromosome-basedCitation49,Citation50 forces and often so malleable that they can be crushed during biopsies (e.g., SCLC cells). This suggests that future drugs hitting mechanisms of nuclear assembly and organization to further weakening the NE structure might result in a synergistic effect and specific killing of such cancer cells (). In support of this hypothesis, several synthetic lethality interactions have been described in the literature between different NE proteins.Citation86,Citation105 For example, B-type lamins were shown to be dispensable in mouse cells expressing A-type lamins but essential in those where lamin A was suppressed.Citation47,Citation106,Citation107
Drug targets that are potentially synthetic lethal with abnormal nuclear organization of some cancer types might be identified either via phenotypic and synthetic lethality screens or by literature mining. Based on the latter one, in my opinion, BAF represents one of the most promising targets for specifically killing cancer cells with altered nuclear appearance. I base this assumption on facts that BAF is often highly expressed in some cancer types (e.g., ovarian cancer, endometrial cancer, breast cancer, colorectal cancer, lung cancer, prostate cancer, glioma, melanoma, and lymphoma),Citation108,Citation109 and it is an important player of post-mitotic nuclear assemblyCitation75,Citation85,Citation96 whose downregulationCitation75,Citation85 or mutation in humansCitation96 delays NE formation and induces strong nuclear irregularities. Due to its small size and relatively flat protein surfaceCitation94,Citation95 without suitable binding pockets for small molecular weight inhibitory compounds, BAF is considered to be undraggable. To overcome this, new anti-mitotics might be designed against its mitotic regulators (). First of all, the dephosphorylation of BAF might be targeted because this is essential for its correct localization and function during mitotic exit.Citation64 Therefore, drugs interfering with either the inhibitory interaction between LEM4 and VRK1, the mitotic kinase of BAF, or with the activating interaction between LEM4 and PP2A, the mitotic phosphatase of BAF, might be developed ().Citation74,Citation75 Consequently, they would enhance the phosphorylation of BAF, hereby weakening its interactions with LEM domain containing INM proteins and DNA.Citation94,Citation95 A second strategy might rely on interfering with the nuclear structure during mitotic entry by inhibiting BAF’s kinases VRK1 and VRK2. Consistently, depletion of VRK1 from breast cancer cells results in retarded tumor growth and reduced incidence of metastasis in a murine orthotopic xenograft model.Citation110 Furthermore, the first inhibitor of BAF phosphorylation with an in vitro anti-cancer activity was recently isolated from a species of tree used in traditional medicine.Citation111 Another approach might aim to target the inhibitory interaction between LBR and importin-β because perturbation of this interaction also results in NE assembly failure, abnormal chromatin decondensation, and daughter cell death.Citation88 Finally, B-type lamins are also promising targets because they are highly expressed in several cancer types,Citation31,Citation108,Citation109 and they might become essential for survival of cancer cells with reduced expression of A-type lamins. Since lamins are also considered to be low-druggable, they might be targeted indirectly by inhibiting their regulators. Such examples include AKAP149, which via promoting PP1 mediates the assembly of B-type lamins into nuclear lamina,Citation112,Citation113 and farnesyltransferase enzymes that add a 15-carbon farnesyl group to the C-terminal of most of the lamins to keep the B-type lamins anchored to the membranes.Citation12
Currently, only a few drugs exist that directly target mechanisms of nuclear assembly and organization, but interestingly, they can induce cancer cell death. For example, the farnesyltransferase inhibitor R115777 inhibits the growth of B-cell lymphomaCitation114 and breast and ovarian cancer cellsCitation115 in vitro and reduces the tumor growth in xenograft model systems. Since lamins are not the only substrates of farnesyltransferases, it is possible that the main anti-tumor activity of this inhibitor is achieved via other substrates, such as the small GTPase Ras proteins. Betulinic acid inhibits the expression of lamin B1 in pancreatic cancer cells and induces dose-dependent anti-cancer activities in both in vitro cultures and xenograft model systems.Citation45 Covalent (NMS-859) and allosteric (NMS-873) inhibitors of p97, an AAA-ATPase known to extract the polyubiquitylated Aurora B from chromatin during post-mitotic nuclear reassembly,Citation80 have antiproliferative effect in vitro on few cancer types.Citation116 However, it is possible that the anti-cancer activity is linked to other roles of p97, such as the ER-associated protein degradation. Obtusilactone B is a new inhibitor of BAF phosphorylation by VRK1, and it also inhibits the proliferation of few cancer cells in vitro.Citation111 In line with this, drugs that directly or indirectly activate PP2A (e.g., ceramide, FTY7220, dithiolethione, etc.), the mitotic phosphatase of BAF, also have anti-cancer activities in different malignancies like prostate cancers, breast cancers, lung cancers, or leukemia.Citation117,Citation118 Finally, microtubules have an influence on the nuclear shape,Citation48,Citation63 and they are directly involved in NE breakdownCitation119,Citation120 and reformation.Citation101 Therefore, microtubule-binding drugs, at least in part, could interfere with the abnormal nuclear structure of some cancer types.
One possible side effect of targeting the mechanisms of nuclear assembly and organization in future anti-cancer therapies might be the artificial induction of laminopathy-like symptoms, such as muscular dystrophies or lipodystrophies. This is suggested by the fact that laminopathies are caused by mutations in different lamina proteins such as lamin A, BAF, LBR, or emerin.Citation51,Citation96 However, laminopathies are developmental disorders, and therefore, it is possible that they might require more time to appear than the duration of the anti-cancer therapy itself. Second, similarly to other anti-mitotic therapies, reduced levels of platelets and blood cells, such as thrombocytopenia or neutropenia, might be also induced. Interestingly, however, neutrophils have multi-lobulated and malleable nuclear structures, probably required for the extrusion of their chromatin fibers to trap and kill bacteria at the sites of infection.Citation121 It is likely that reduced expression of BAFCitation108,Citation109 and its binding partners emerin, LAP2β, lamin A/C, and lamin B2Citation122,Citation123 might account for such nuclear appearance. Therefore, targeting proteins that are highly expressed in cancer cells but repressed in neutrophils could lead to at least the terminally differentiated neutrophils being resistant to these new drugs.
Concluding Remarks
Cancer is one of the leading causes of mortality worldwide. Anti-cancer drugs, in addition to surgery, have been proven to be beneficial for patients with particular cancer types; however, their effectiveness is often limited by dose-limiting toxicities. Therefore, there is a need to develop new drugs that can achieve efficient and cancer cell-specific effects without undesirable side effects. In my opinion, particular molecular mechanisms of post-mitotic nuclear assembly and nuclear organization represent attractive new targets for such next-generation anti-cancer therapies. This strategy relies on putative synthetic lethality between the altered nuclear structure of some cancer types and drugs targeting, directly or indirectly, BAF, B-type lamins, VRK1, VRK2, and other similar proteins that are important for proper assembly and organization of the nuclei and are clearly expressed or overexpressed in these cancer types.
Disclosure of Potential Conflicts of Interest
No potential conflict of interest was disclosed.
Acknowledgments
I would like to thank Iain W Mattaj, Karl Ziegelbauer, Volker Gekeler, and Dominik Mumberg for critically reading and discussing the manuscript.
References
- Chan KS, Koh CG, Li HY. Mitosis-targeted anti-cancer therapies: where they stand. Cell Death Dis 2012; 3:e411; http://dx.doi.org/10.1038/cddis.2012.148; PMID: 23076219
- Janssen A, Medema RH. Mitosis as an anti-cancer target. Oncogene 2011; 30:2799 - 809; http://dx.doi.org/10.1038/onc.2011.30; PMID: 21339734
- Jordan MA, Wilson L. Microtubules as a target for anticancer drugs. Nat Rev Cancer 2004; 4:253 - 65; http://dx.doi.org/10.1038/nrc1317; PMID: 15057285
- Gascoigne KE, Taylor SS. How do anti-mitotic drugs kill cancer cells?. J Cell Sci 2009; 122:2579 - 85; http://dx.doi.org/10.1242/jcs.039719; PMID: 19625502
- Huang HC, Mitchison TJ, Shi J. Stochastic competition between mechanistically independent slippage and death pathways determines cell fate during mitotic arrest. PLoS One 2010; 5:e15724; http://dx.doi.org/10.1371/journal.pone.0015724; PMID: 21203573
- Schmidt M, Bastians H. Mitotic drug targets and the development of novel anti-mitotic anticancer drugs. Drug Resist Updat 2007; 10:162 - 81; http://dx.doi.org/10.1016/j.drup.2007.06.003; PMID: 17669681
- Komlodi-Pasztor E, Sackett DL, Fojo AT. Inhibitors targeting mitosis: tales of how great drugs against a promising target were brought down by a flawed rationale. Clin Cancer Res 2012; 18:51 - 63; http://dx.doi.org/10.1158/1078-0432.CCR-11-0999; PMID: 22215906
- Mitchison TJ. The proliferation rate paradox in antimitotic chemotherapy. Mol Biol Cell 2012; 23:1 - 6; http://dx.doi.org/10.1091/mbc.E10-04-0335; PMID: 22210845
- Komlodi-Pasztor E, Sackett D, Wilkerson J, Fojo T. Mitosis is not a key target of microtubule agents in patient tumors. Nat Rev Clin Oncol 2011; 8:244 - 50; http://dx.doi.org/10.1038/nrclinonc.2010.228; PMID: 21283127
- Gorjánácz M, Jaedicke A, Mattaj IW. What can Caenorhabditis elegans tell us about the nuclear envelope?. FEBS Lett 2007; 581:2794 - 801; http://dx.doi.org/10.1016/j.febslet.2007.03.052; PMID: 17418822
- Hetzer MW, Walther TC, Mattaj IW. Pushing the envelope: structure, function, and dynamics of the nuclear periphery. Annu Rev Cell Dev Biol 2005; 21:347 - 80; http://dx.doi.org/10.1146/annurev.cellbio.21.090704.151152; PMID: 16212499
- Dechat T, Pfleghaar K, Sengupta K, Shimi T, Shumaker DK, Solimando L, Goldman RD. Nuclear lamins: major factors in the structural organization and function of the nucleus and chromatin. Genes Dev 2008; 22:832 - 53; http://dx.doi.org/10.1101/gad.1652708; PMID: 18381888
- Dittmer TA, Misteli T. The lamin protein family. Genome Biol 2011; 12:222; http://dx.doi.org/10.1186/gb-2011-12-5-222; PMID: 21639948
- de Las Heras JI, Meinke P, Batrakou DG, Srsen V, Zuleger N, Kerr AR, Schirmer EC. Tissue specificity in the nuclear envelope supports its functional complexity. Nucleus 2013; 4:4; http://dx.doi.org/10.4161/nucl.26872; PMID: 24213376
- Lin F, Worman HJ. Expression of nuclear lamins in human tissues and cancer cell lines and transcription from the promoters of the lamin A/C and B1 genes. Exp Cell Res 1997; 236:378 - 84; http://dx.doi.org/10.1006/excr.1997.3735; PMID: 9367621
- Swift J, Ivanovska IL, Buxboim A, Harada T, Dingal PC, Pinter J, Pajerowski JD, Spinler KR, Shin JW, Tewari M, et al. Nuclear lamin-A scales with tissue stiffness and enhances matrix-directed differentiation. Science 2013; 341:1240104; http://dx.doi.org/10.1126/science.1240104; PMID: 23990565
- Ben-Harush K, Wiesel N, Frenkiel-Krispin D, Moeller D, Soreq E, Aebi U, Herrmann H, Gruenbaum Y, Medalia O. The supramolecular organization of the C. elegans nuclear lamin filament. J Mol Biol 2009; 386:1392 - 402; http://dx.doi.org/10.1016/j.jmb.2008.12.024; PMID: 19109977
- Goldberg MW, Huttenlauch I, Hutchison CJ, Stick R. Filaments made from A- and B-type lamins differ in structure and organization. J Cell Sci 2008; 121:215 - 25; http://dx.doi.org/10.1242/jcs.022020; PMID: 18187453
- Lammerding J, Fong LG, Ji JY, Reue K, Stewart CL, Young SG, Lee RT. Lamins A and C but not lamin B1 regulate nuclear mechanics. J Biol Chem 2006; 281:25768 - 80; http://dx.doi.org/10.1074/jbc.M513511200; PMID: 16825190
- Peric-Hupkes D, Meuleman W, Pagie L, Bruggeman SW, Solovei I, Brugman W, Gräf S, Flicek P, Kerkhoven RM, van Lohuizen M, et al. Molecular maps of the reorganization of genome-nuclear lamina interactions during differentiation. Mol Cell 2010; 38:603 - 13; http://dx.doi.org/10.1016/j.molcel.2010.03.016; PMID: 20513434
- Wiesel N, Mattout A, Melcer S, Melamed-Book N, Herrmann H, Medalia O, Aebi U, Gruenbaum Y. Laminopathic mutations interfere with the assembly, localization, and dynamics of nuclear lamins. Proc Natl Acad Sci U S A 2008; 105:180 - 5; http://dx.doi.org/10.1073/pnas.0708974105; PMID: 18162544
- Melcer S, Gruenbaum Y, Krohne G. Invertebrate lamins. Exp Cell Res 2007; 313:2157 - 66; http://dx.doi.org/10.1016/j.yexcr.2007.03.004; PMID: 17451683
- Lammerding J, Hsiao J, Schulze PC, Kozlov S, Stewart CL, Lee RT. Abnormal nuclear shape and impaired mechanotransduction in emerin-deficient cells. J Cell Biol 2005; 170:781 - 91; http://dx.doi.org/10.1083/jcb.200502148; PMID: 16115958
- Chow KH, Factor RE, Ullman KS. The nuclear envelope environment and its cancer connections. Nat Rev Cancer 2012; 12:196 - 209; PMID: 22337151
- Las Heras JI, Batrakou DG, Schirmer EC. Cancer biology and the nuclear envelope: a convoluted relationship. Semin Cancer Biol 2013; 23:125 - 37; http://dx.doi.org/10.1016/j.semcancer.2012.01.008; PMID: 22311402
- Vargas JD, Hatch EM, Anderson DJ, Hetzer MW. Transient nuclear envelope rupturing during interphase in human cancer cells. Nucleus 2012; 3:88 - 100; http://dx.doi.org/10.4161/nucl.18954; PMID: 22567193
- Bussolati G. Proper detection of the nuclear shape: ways and significance. Rom J Morphol Embryol 2008; 49:435 - 9; PMID: 19050790
- Bussolati G, Marchiò C, Gaetano L, Lupo R, Sapino A. Pleomorphism of the nuclear envelope in breast cancer: a new approach to an old problem. J Cell Mol Med 2008; 12:209 - 18; http://dx.doi.org/10.1111/j.1582-4934.2007.00176.x; PMID: 18053086
- Fischer AH, Bardarov S Jr., Jiang Z. Molecular aspects of diagnostic nucleolar and nuclear envelope changes in prostate cancer. J Cell Biochem 2004; 91:170 - 84; http://dx.doi.org/10.1002/jcb.10735; PMID: 14689589
- Petersen I, Kotb WF, Friedrich KH, Schlüns K, Böcking A, Dietel M. Core classification of lung cancer: correlating nuclear size and mitoses with ploidy and clinicopathological parameters. Lung Cancer 2009; 65:312 - 8; http://dx.doi.org/10.1016/j.lungcan.2008.12.013; PMID: 19168259
- Foster CR, Przyborski SA, Wilson RG, Hutchison CJ. Lamins as cancer biomarkers. Biochem Soc Trans 2010; 38:297 - 300; http://dx.doi.org/10.1042/BST0380297; PMID: 20074078
- Broers JL, Raymond Y, Rot MK, Kuijpers H, Wagenaar SS, Ramaekers FC. Nuclear A-type lamins are differentially expressed in human lung cancer subtypes. Am J Pathol 1993; 143:211 - 20; PMID: 8391215
- Kaufmann SH, Mabry M, Jasti R, Shaper JH. Differential expression of nuclear envelope lamins A and C in human lung cancer cell lines. Cancer Res 1991; 51:581 - 6; PMID: 1985776
- Machiels BM, Broers JL, Raymond Y, de Ley L, Kuijpers HJ, Caberg NE, Ramaekers FC. Abnormal A-type lamin organization in a human lung carcinoma cell line. Eur J Cell Biol 1995; 67:328 - 35; PMID: 8521872
- Belt EJ, Fijneman RJ, van den Berg EG, Bril H, Delis-van Diemen PM, Tijssen M, van Essen HF, de Lange-de Klerk ES, Beliën JA, Stockmann HB, et al. Loss of lamin A/C expression in stage II and III colon cancer is associated with disease recurrence. Eur J Cancer 2011; 47:1837 - 45; http://dx.doi.org/10.1016/j.ejca.2011.04.025; PMID: 21621406
- Moss SF, Krivosheyev V, de Souza A, Chin K, Gaetz HP, Chaudhary N, Worman HJ, Holt PR. Decreased and aberrant nuclear lamin expression in gastrointestinal tract neoplasms. Gut 1999; 45:723 - 9; http://dx.doi.org/10.1136/gut.45.5.723; PMID: 10517909
- Wu Z, Wu L, Weng D, Xu D, Geng J, Zhao F. Reduced expression of lamin A/C correlates with poor histological differentiation and prognosis in primary gastric carcinoma. Journal of experimental & clinical cancer research. CR (East Lansing, Mich) 2009; 28:8
- Capo-chichi CD, Cai KQ, Smedberg J, Ganjei-Azar P, Godwin AK, Xu XX. Loss of A-type lamin expression compromises nuclear envelope integrity in breast cancer. Chin J Cancer 2011; 30:415 - 25; http://dx.doi.org/10.5732/cjc.010.10566; PMID: 21627864
- Wazir U, Ahmed MH, Bridger JM, Harvey A, Jiang WG, Sharma AK, Mokbel K. The clinicopathological significance of lamin A/C, lamin B1 and lamin B receptor mRNA expression in human breast cancer. Cell Mol Biol Lett 2013; 18:595 - 611; http://dx.doi.org/10.2478/s11658-013-0109-9; PMID: 24293108
- Agrelo R, Setien F, Espada J, Artiga MJ, Rodriguez M, Pérez-Rosado A, Sanchez-Aguilera A, Fraga MF, Piris MA, Esteller M. Inactivation of the lamin A/C gene by CpG island promoter hypermethylation in hematologic malignancies, and its association with poor survival in nodal diffuse large B-cell lymphoma. J Clin Oncol 2005; 23:3940 - 7; http://dx.doi.org/10.1200/JCO.2005.11.650; PMID: 15867203
- Helfand BT, Wang Y, Pfleghaar K, Shimi T, Taimen P, Shumaker DK. Chromosomal regions associated with prostate cancer risk localize to lamin B-deficient microdomains and exhibit reduced gene transcription. J Pathol 2012; 226:735 - 45; http://dx.doi.org/10.1002/path.3033; PMID: 22025297
- Funkhouser CM, Sknepnek R, Shimi T, Goldman AE, Goldman RD, Olvera de la Cruz M. Mechanical model of blebbing in nuclear lamin meshworks. Proc Natl Acad Sci U S A 2013; 110:3248 - 53; http://dx.doi.org/10.1073/pnas.1300215110; PMID: 23401537
- Capo-chichi CD, Cai KQ, Simpkins F, Ganjei-Azar P, Godwin AK, Xu XX. Nuclear envelope structural defects cause chromosomal numerical instability and aneuploidy in ovarian cancer. BMC Med 2011; 9:28; http://dx.doi.org/10.1186/1741-7015-9-28; PMID: 21439080
- Stadelmann B, Khandjian E, Hirt A, Lüthy A, Weil R, Wagner HP. Repression of nuclear lamin A and C gene expression in human acute lymphoblastic leukemia and non-Hodgkin’s lymphoma cells. Leuk Res 1990; 14:815 - 21; http://dx.doi.org/10.1016/0145-2126(90)90076-L; PMID: 2232854
- Li L, Du Y, Kong X, Li Z, Jia Z, Cui J, Gao J, Wang G, Xie K. Lamin B1 is a novel therapeutic target of betulinic acid in pancreatic cancer. Clin Cancer Res 2013; 19:4651 - 61; http://dx.doi.org/10.1158/1078-0432.CCR-12-3630; PMID: 23857605
- Kong L, Schäfer G, Bu H, Zhang Y, Zhang Y, Klocker H. Lamin A/C protein is overexpressed in tissue-invading prostate cancer and promotes prostate cancer cell growth, migration and invasion through the PI3K/AKT/PTEN pathway. Carcinogenesis 2012; 33:751 - 9; http://dx.doi.org/10.1093/carcin/bgs022; PMID: 22301279
- Coffinier C, Jung HJ, Nobumori C, Chang S, Tu Y, Barnes RH 2nd, Yoshinaga Y, de Jong PJ, Vergnes L, Reue K, et al. Deficiencies in lamin B1 and lamin B2 cause neurodevelopmental defects and distinct nuclear shape abnormalities in neurons. Mol Biol Cell 2011; 22:4683 - 93; http://dx.doi.org/10.1091/mbc.E11-06-0504; PMID: 21976703
- Fukuda R, Hayashi A, Utsunomiya A, Nukada Y, Fukui R, Itoh K, Tezuka K, Ohashi K, Mizuno K, Sakamoto M, et al. Alteration of phosphatidylinositol 3-kinase cascade in the multilobulated nuclear formation of adult T cell leukemia/lymphoma (ATLL). Proc Natl Acad Sci U S A 2005; 102:15213 - 8; http://dx.doi.org/10.1073/pnas.0507184102; PMID: 16217039
- Chuang CH, Carpenter AE, Fuchsova B, Johnson T, de Lanerolle P, Belmont AS. Long-range directional movement of an interphase chromosome site. Curr Biol 2006; 16:825 - 31; http://dx.doi.org/10.1016/j.cub.2006.03.059; PMID: 16631592
- Weber SC, Spakowitz AJ, Theriot JA. Nonthermal ATP-dependent fluctuations contribute to the in vivo motion of chromosomal loci. Proc Natl Acad Sci U S A 2012; 109:7338 - 43; http://dx.doi.org/10.1073/pnas.1119505109; PMID: 22517744
- Maraldi NM, Capanni C, Cenni V, Fini M, Lattanzi G. Laminopathies and lamin-associated signaling pathways. J Cell Biochem 2011; 112:979 - 92; http://dx.doi.org/10.1002/jcb.22992; PMID: 21400569
- De Vos WH, Houben F, Kamps M, Malhas A, Verheyen F, Cox J, Manders EM, Verstraeten VL, van Steensel MA, Marcelis CL, et al. Repetitive disruptions of the nuclear envelope invoke temporary loss of cellular compartmentalization in laminopathies. Hum Mol Genet 2011; 20:4175 - 86; http://dx.doi.org/10.1093/hmg/ddr344; PMID: 21831885
- Capell BC, Collins FS. Human laminopathies: nuclei gone genetically awry. Nat Rev Genet 2006; 7:940 - 52; http://dx.doi.org/10.1038/nrg1906; PMID: 17139325
- Willis ND, Cox TR, Rahman-Casañs SF, Smits K, Przyborski SA, van den Brandt P, van Engeland M, Weijenberg M, Wilson RG, de Bruïne A, et al. Lamin A/C is a risk biomarker in colorectal cancer. PLoS One 2008; 3:e2988; http://dx.doi.org/10.1371/journal.pone.0002988; PMID: 18714339
- Puckelwartz MJ, Depreux FF, McNally EM. Gene expression, chromosome position and lamin A/C mutations. Nucleus 2011; 2:162 - 7; http://dx.doi.org/10.4161/nucl.2.3.16003; PMID: 21818408
- Schirmer EC, Foisner R. Proteins that associate with lamins: many faces, many functions. Exp Cell Res 2007; 313:2167 - 79; http://dx.doi.org/10.1016/j.yexcr.2007.03.012; PMID: 17451680
- Wilson KL, Foisner R. Lamin-binding Proteins. Cold Spring Harb Perspect Biol 2010; 2:a000554; http://dx.doi.org/10.1101/cshperspect.a000554; PMID: 20452940
- Bank EM, Ben-Harush K, Feinstein N, Medalia O, Gruenbaum Y. Structural and physiological phenotypes of disease-linked lamin mutations in C. elegans. J Struct Biol 2012; 177:106 - 12; http://dx.doi.org/10.1016/j.jsb.2011.10.009; PMID: 22079399
- Lammerding J, Lee RT. The nuclear membrane and mechanotransduction: impaired nuclear mechanics and mechanotransduction in lamin A/C deficient cells. Novartis Found Symp 2005; 264:264 - 73, discussion 273-8; http://dx.doi.org/10.1002/0470093765.ch18; PMID: 15773759
- Scaffidi P, Misteli T. Lamin A-dependent nuclear defects in human aging. Science 2006; 312:1059 - 63; http://dx.doi.org/10.1126/science.1127168; PMID: 16645051
- Pekovic V, Harborth J, Broers JL, Ramaekers FC, van Engelen B, Lammens M, von Zglinicki T, Foisner R, Hutchison C, Markiewicz E. Nucleoplasmic LAP2alpha-lamin A complexes are required to maintain a proliferative state in human fibroblasts. J Cell Biol 2007; 176:163 - 72; http://dx.doi.org/10.1083/jcb.200606139; PMID: 17227891
- Meaburn KJ, Gudla PR, Khan S, Lockett SJ, Misteli T. Disease-specific gene repositioning in breast cancer. J Cell Biol 2009; 187:801 - 12; http://dx.doi.org/10.1083/jcb.200909127; PMID: 19995938
- Fischer AH, Taysavang P, Jhiang SM. Nuclear envelope irregularity is induced by RET/PTC during interphase. Am J Pathol 2003; 163:1091 - 100; http://dx.doi.org/10.1016/S0002-9440(10)63468-2; PMID: 12937150
- Gorjánácz M. LEM-4 promotes rapid dephosphorylation of BAF during mitotic exit. Nucleus 2013; 4:14 - 7; http://dx.doi.org/10.4161/nucl.22961; PMID: 23211644
- Bollen M, Gerlich DW, Lesage B. Mitotic phosphatases: from entry guards to exit guides. Trends Cell Biol 2009; 19:531 - 41; http://dx.doi.org/10.1016/j.tcb.2009.06.005; PMID: 19734049
- Wurzenberger C, Gerlich DW. Phosphatases: providing safe passage through mitotic exit. Nat Rev Mol Cell Biol 2011; 12:469 - 82; http://dx.doi.org/10.1038/nrm3149; PMID: 21750572
- Gong D, Pomerening JR, Myers JW, Gustavsson C, Jones JT, Hahn AT, Meyer T, Ferrell JE Jr.. Cyclin A2 regulates nuclear-envelope breakdown and the nuclear accumulation of cyclin B1. Curr Biol 2007; 17:85 - 91; http://dx.doi.org/10.1016/j.cub.2006.11.066; PMID: 17208191
- Laurell E, Beck K, Krupina K, Theerthagiri G, Bodenmiller B, Horvath P, Aebersold R, Antonin W, Kutay U. Phosphorylation of Nup98 by multiple kinases is crucial for NPC disassembly during mitotic entry. Cell 2011; 144:539 - 50; http://dx.doi.org/10.1016/j.cell.2011.01.012; PMID: 21335236
- Peter M, Nakagawa J, Dorée M, Labbé JC, Nigg EA. In vitro disassembly of the nuclear lamina and M phase-specific phosphorylation of lamins by cdc2 kinase. Cell 1990; 61:591 - 602; http://dx.doi.org/10.1016/0092-8674(90)90471-P; PMID: 2188731
- Gorjánácz M, Mattaj IW. Lipin is required for efficient breakdown of the nuclear envelope in Caenorhabditis elegans. J Cell Sci 2009; 122:1963 - 9; http://dx.doi.org/10.1242/jcs.044750; PMID: 19494125
- Mall M, Walter T, Gorjánácz M, Davidson IF, Nga Ly-Hartig TB, Ellenberg J, Mattaj IW. Mitotic lamin disassembly is triggered by lipid-mediated signaling. J Cell Biol 2012; 198:981 - 90; http://dx.doi.org/10.1083/jcb.201205103; PMID: 22986494
- Portier N, Audhya A, Maddox PS, Green RA, Dammermann A, Desai A, Oegema K. A microtubule-independent role for centrosomes and aurora a in nuclear envelope breakdown. Dev Cell 2007; 12:515 - 29; http://dx.doi.org/10.1016/j.devcel.2007.01.019; PMID: 17419991
- Li H, Liu XS, Yang X, Song B, Wang Y, Liu X. Polo-like kinase 1 phosphorylation of p150Glued facilitates nuclear envelope breakdown during prophase. Proc Natl Acad Sci U S A 2010; 107:14633 - 8; http://dx.doi.org/10.1073/pnas.1006615107; PMID: 20679239
- Asencio C, Davidson IF, Santarella-Mellwig R, Ly-Hartig TB, Mall M, Wallenfang MR, Mattaj IW, Gorjánácz M. Coordination of kinase and phosphatase activities by Lem4 enables nuclear envelope reassembly during mitosis. Cell 2012; 150:122 - 35; http://dx.doi.org/10.1016/j.cell.2012.04.043; PMID: 22770216
- Gorjánácz M, Klerkx EP, Galy V, Santarella R, López-Iglesias C, Askjaer P, Mattaj IW. Caenorhabditis elegans BAF-1 and its kinase VRK-1 participate directly in post-mitotic nuclear envelope assembly. EMBO J 2007; 26:132 - 43; http://dx.doi.org/10.1038/sj.emboj.7601470; PMID: 17170708
- Nichols RJ, Wiebe MS, Traktman P. The vaccinia-related kinases phosphorylate the N’ terminus of BAF, regulating its interaction with DNA and its retention in the nucleus. Mol Biol Cell 2006; 17:2451 - 64; http://dx.doi.org/10.1091/mbc.E05-12-1179; PMID: 16495336
- Kim W, Chakraborty G, Kim S, Shin J, Park CH, Jeong MW, Bharatham N, Yoon HS, Kim KT. Macro histone H2A1.2 (macroH2A1) protein suppresses mitotic kinase VRK1 during interphase. J Biol Chem 2012; 287:5278 - 89; http://dx.doi.org/10.1074/jbc.M111.281709; PMID: 22194607
- Lu L, Ladinsky MS, Kirchhausen T. Cisternal organization of the endoplasmic reticulum during mitosis. Mol Biol Cell 2009; 20:3471 - 80; http://dx.doi.org/10.1091/mbc.E09-04-0327; PMID: 19494040
- Vagnarelli P, Ribeiro S, Sennels L, Sanchez-Pulido L, de Lima Alves F, Verheyen T, Kelly DA, Ponting CP, Rappsilber J, Earnshaw WC. Repo-Man coordinates chromosomal reorganization with nuclear envelope reassembly during mitotic exit. Dev Cell 2011; 21:328 - 42; http://dx.doi.org/10.1016/j.devcel.2011.06.020; PMID: 21820363
- Ramadan K, Bruderer R, Spiga FM, Popp O, Baur T, Gotta M, Meyer HH. Cdc48/p97 promotes reformation of the nucleus by extracting the kinase Aurora B from chromatin. Nature 2007; 450:1258 - 62; http://dx.doi.org/10.1038/nature06388; PMID: 18097415
- Anderson DJ, Hetzer MW. Nuclear envelope formation by chromatin-mediated reorganization of the endoplasmic reticulum. Nat Cell Biol 2007; 9:1160 - 6; http://dx.doi.org/10.1038/ncb1636; PMID: 17828249
- Anderson DJ, Hetzer MW. Reshaping of the endoplasmic reticulum limits the rate for nuclear envelope formation. J Cell Biol 2008; 182:911 - 24; http://dx.doi.org/10.1083/jcb.200805140; PMID: 18779370
- Lu L, Ladinsky MS, Kirchhausen T. Formation of the postmitotic nuclear envelope from extended ER cisternae precedes nuclear pore assembly. J Cell Biol 2011; 194:425 - 40; http://dx.doi.org/10.1083/jcb.201012063; PMID: 21825076
- Ulbert S, Platani M, Boue S, Mattaj IW. Direct membrane protein-DNA interactions required early in nuclear envelope assembly. J Cell Biol 2006; 173:469 - 76; http://dx.doi.org/10.1083/jcb.200512078; PMID: 16717124
- Anderson DJ, Vargas JD, Hsiao JP, Hetzer MW. Recruitment of functionally distinct membrane proteins to chromatin mediates nuclear envelope formation in vivo. J Cell Biol 2009; 186:183 - 91; http://dx.doi.org/10.1083/jcb.200901106; PMID: 19620630
- Liu J, Lee KK, Segura-Totten M, Neufeld E, Wilson KL, Gruenbaum Y. MAN1 and emerin have overlapping function(s) essential for chromosome segregation and cell division in Caenorhabditis elegans. Proc Natl Acad Sci U S A 2003; 100:4598 - 603; http://dx.doi.org/10.1073/pnas.0730821100; PMID: 12684533
- Kaláb P, Pralle A, Isacoff EY, Heald R, Weis K. Analysis of a RanGTP-regulated gradient in mitotic somatic cells. Nature 2006; 440:697 - 701; http://dx.doi.org/10.1038/nature04589; PMID: 16572176
- Lu X, Shi Y, Lu Q, Ma Y, Luo J, Wang Q, Ji J, Jiang Q, Zhang C. Requirement for lamin B receptor and its regulation by importin beta and phosphorylation in nuclear envelope assembly during mitotic exit. J Biol Chem 2010; 285:33281 - 93; http://dx.doi.org/10.1074/jbc.M110.102368; PMID: 20576617
- Ma Y, Cai S, Lv Q, Jiang Q, Zhang Q, Sodmergen, Zhai Z, Zhang C. Lamin B receptor plays a role in stimulating nuclear envelope production and targeting membrane vesicles to chromatin during nuclear envelope assembly through direct interaction with importin beta. J Cell Sci 2007; 120:520 - 30; http://dx.doi.org/10.1242/jcs.03355; PMID: 17251381
- Polioudaki H, Kourmouli N, Drosou V, Bakou A, Theodoropoulos PA, Singh PB, Giannakouros T, Georgatos SD. Histones H3/H4 form a tight complex with the inner nuclear membrane protein LBR and heterochromatin protein 1. EMBO Rep 2001; 2:920 - 5; http://dx.doi.org/10.1093/embo-reports/kve199; PMID: 11571267
- Ye Q, Worman HJ. Interaction between an integral protein of the nuclear envelope inner membrane and human chromodomain proteins homologous to Drosophila HP1. J Biol Chem 1996; 271:14653 - 6; http://dx.doi.org/10.1074/jbc.271.25.14653; PMID: 8663349
- Audhya A, Desai A, Oegema K. A role for Rab5 in structuring the endoplasmic reticulum. J Cell Biol 2007; 178:43 - 56; http://dx.doi.org/10.1083/jcb.200701139; PMID: 17591921
- Golden A, Liu J, Cohen-Fix O. Inactivation of the C. elegans lipin homolog leads to ER disorganization and to defects in the breakdown and reassembly of the nuclear envelope. J Cell Sci 2009; 122:1970 - 8; http://dx.doi.org/10.1242/jcs.044743; PMID: 19494126
- Bradley CM, Ronning DR, Ghirlando R, Craigie R, Dyda F. Structural basis for DNA bridging by barrier-to-autointegration factor. Nat Struct Mol Biol 2005; 12:935 - 6; http://dx.doi.org/10.1038/nsmb989; PMID: 16155580
- Cai M, Huang Y, Suh JY, Louis JM, Ghirlando R, Craigie R, Clore GM. Solution NMR structure of the barrier-to-autointegration factor-Emerin complex. J Biol Chem 2007; 282:14525 - 35; http://dx.doi.org/10.1074/jbc.M700576200; PMID: 17355960
- Puente XS, Quesada V, Osorio FG, Cabanillas R, Cadiñanos J, Fraile JM, Ordóñez GR, Puente DA, Gutiérrez-Fernández A, Fanjul-Fernández M, et al. Exome sequencing and functional analysis identifies BANF1 mutation as the cause of a hereditary progeroid syndrome. Am J Hum Genet 2011; 88:650 - 6; http://dx.doi.org/10.1016/j.ajhg.2011.04.010; PMID: 21549337
- Schmitz MH, Held M, Janssens V, Hutchins JR, Hudecz O, Ivanova E, Goris J, Trinkle-Mulcahy L, Lamond AI, Poser I, et al. Live-cell imaging RNAi screen identifies PP2A-B55alpha and importin-beta1 as key mitotic exit regulators in human cells. Nat Cell Biol 2010; 12:886 - 93; http://dx.doi.org/10.1038/ncb2092; PMID: 20711181
- Chen GI, Tisayakorn S, Jorgensen C, D’Ambrosio LM, Goudreault M, Gingras AC. PP4R4/KIAA1622 forms a novel stable cytosolic complex with phosphoprotein phosphatase 4. J Biol Chem 2008; 283:29273 - 84; http://dx.doi.org/10.1074/jbc.M803443200; PMID: 18715871
- Glatter T, Wepf A, Aebersold R, Gstaiger M. An integrated workflow for charting the human interaction proteome: insights into the PP2A system. Mol Syst Biol 2009; 5:237; http://dx.doi.org/10.1038/msb.2008.75; PMID: 19156129
- Zhuang X, Semenova E, Maric D, Craigie R. Dephosphorylation of Barrier-to-autointegration Factor by Protein Phosphatase 4 and Its Role in Cell Mitosis. J Biol Chem 2014; 289:1119 - 27; http://dx.doi.org/10.1074/jbc.M113.492777; PMID: 24265311
- Haraguchi T, Kojidani T, Koujin T, Shimi T, Osakada H, Mori C, Yamamoto A, Hiraoka Y. Live cell imaging and electron microscopy reveal dynamic processes of BAF-directed nuclear envelope assembly. J Cell Sci 2008; 121:2540 - 54; http://dx.doi.org/10.1242/jcs.033597; PMID: 18628300
- Doucet CM, Talamas JA, Hetzer MW. Cell cycle-dependent differences in nuclear pore complex assembly in metazoa. Cell 2010; 141:1030 - 41; http://dx.doi.org/10.1016/j.cell.2010.04.036; PMID: 20550937
- Baur T, Ramadan K, Schlundt A, Kartenbeck J, Meyer HH. NSF- and SNARE-mediated membrane fusion is required for nuclear envelope formation and completion of nuclear pore complex assembly in Xenopus laevis egg extracts. J Cell Sci 2007; 120:2895 - 903; http://dx.doi.org/10.1242/jcs.010181; PMID: 17666429
- Loewinger L, McKeon F. Mutations in the nuclear lamin proteins resulting in their aberrant assembly in the cytoplasm. EMBO J 1988; 7:2301 - 9; PMID: 3056713
- Galy V, Mattaj IW, Askjaer P. Caenorhabditis elegans nucleoporins Nup93 and Nup205 determine the limit of nuclear pore complex size exclusion in vivo. Mol Biol Cell 2003; 14:5104 - 15; http://dx.doi.org/10.1091/mbc.E03-04-0237; PMID: 12937276
- Yang SH, Jung HJ, Coffinier C, Fong LG, Young SG. Are B-type lamins essential in all mammalian cells?. Nucleus 2011; 2:562 - 9; http://dx.doi.org/10.4161/nucl.2.6.18085; PMID: 22127257
- Yang SH, Chang SY, Yin L, Tu Y, Hu Y, Yoshinaga Y, de Jong PJ, Fong LG, Young SG. An absence of both lamin B1 and lamin B2 in keratinocytes has no effect on cell proliferation or the development of skin and hair. Hum Mol Genet 2011; 20:3537 - 44; http://dx.doi.org/10.1093/hmg/ddr266; PMID: 21659336
- Hruz T, Laule O, Szabo G, Wessendorp F, Bleuler S, Oertle L, Widmayer P, Gruissem W, Zimmermann P. Genevestigator v3: a reference expression database for the meta-analysis of transcriptomes. Adv Bioinformatics 2008; 2008:420747; http://dx.doi.org/10.1155/2008/420747; PMID: 19956698
- Uhlen M, Oksvold P, Fagerberg L, Lundberg E, Jonasson K, Forsberg M, Zwahlen M, Kampf C, Wester K, Hober S, et al. Towards a knowledge-based Human Protein Atlas. Nat Biotechnol 2010; 28:1248 - 50; http://dx.doi.org/10.1038/nbt1210-1248; PMID: 21139605
- Molitor TP, Traktman P. Molecular genetic analysis of VRK1 in mammary epithelial cells: depletion slows proliferation in vitro and tumor growth and metastasis in vivo. Oncogenesis 2013; 2:e48; http://dx.doi.org/10.1038/oncsis.2013.11; PMID: 23732708
- Kim W, Lyu HN, Kwon HS, Kim YS, Lee KH, Kim DY, Chakraborty G, Choi KY, Yoon HS, Kim KT. Obtusilactone B from Machilus Thunbergii targets barrier-to-autointegration factor to treat cancer. Mol Pharmacol 2013; 83:367 - 76; http://dx.doi.org/10.1124/mol.112.082578; PMID: 23150487
- Steen RL, Collas P. Mistargeting of B-type lamins at the end of mitosis: implications on cell survival and regulation of lamins A/C expression. J Cell Biol 2001; 153:621 - 6; http://dx.doi.org/10.1083/jcb.153.3.621; PMID: 11331311
- Steen RL, Martins SB, Taskén K, Collas P. Recruitment of protein phosphatase 1 to the nuclear envelope by A-kinase anchoring protein AKAP149 is a prerequisite for nuclear lamina assembly. J Cell Biol 2000; 150:1251 - 62; http://dx.doi.org/10.1083/jcb.150.6.1251; PMID: 10995432
- Rolland D, Camara-Clayette V, Barbarat A, Salles G, Coiffier B, Ribrag V, Thieblemont C. Farnesyltransferase inhibitor R115777 inhibits cell growth and induces apoptosis in mantle cell lymphoma. Cancer Chemother Pharmacol 2008; 61:855 - 63; http://dx.doi.org/10.1007/s00280-007-0543-3; PMID: 17639395
- Wärnberg F, White D, Anderson E, Knox F, Clarke RB, Morris J, Bundred NJ. Effect of a farnesyl transferase inhibitor (R115777) on ductal carcinoma in situ of the breast in a human xenograft model and on breast and ovarian cancer cell growth in vitro and in vivo. Breast Cancer Res 2006; 8:R21; http://dx.doi.org/10.1186/bcr1395; PMID: 16611371
- Magnaghi P, D’Alessio R, Valsasina B, Avanzi N, Rizzi S, Asa D, Gasparri F, Cozzi L, Cucchi U, Orrenius C, et al. Covalent and allosteric inhibitors of the ATPase VCP/p97 induce cancer cell death. Nat Chem Biol 2013; 9:548 - 56; http://dx.doi.org/10.1038/nchembio.1313; PMID: 23892893
- Chen W, Wang Z, Jiang C, Ding Y. PP2A-Mediated Anticancer Therapy. Gastroenterol Res Pract 2013; 2013:675429; http://dx.doi.org/10.1155/2013/675429; PMID: 24307892
- Kalev P, Sablina AA. Protein phosphatase 2A as a potential target for anticancer therapy. Anticancer Agents Med Chem 2011; 11:38 - 46; http://dx.doi.org/10.2174/187152011794941172; PMID: 21288198
- Beaudouin J, Gerlich D, Daigle N, Eils R, Ellenberg J. Nuclear envelope breakdown proceeds by microtubule-induced tearing of the lamina. Cell 2002; 108:83 - 96; http://dx.doi.org/10.1016/S0092-8674(01)00627-4; PMID: 11792323
- Mühlhäusser P, Kutay U. An in vitro nuclear disassembly system reveals a role for the RanGTPase system and microtubule-dependent steps in nuclear envelope breakdown. J Cell Biol 2007; 178:595 - 610; http://dx.doi.org/10.1083/jcb.200703002; PMID: 17698605
- Brinkmann V, Reichard U, Goosmann C, Fauler B, Uhlemann Y, Weiss DS, Weinrauch Y, Zychlinsky A. Neutrophil extracellular traps kill bacteria. Science 2004; 303:1532 - 5; http://dx.doi.org/10.1126/science.1092385; PMID: 15001782
- Olins AL, Zwerger M, Herrmann H, Zentgraf H, Simon AJ, Monestier M, Olins DE. The human granulocyte nucleus: Unusual nuclear envelope and heterochromatin composition. Eur J Cell Biol 2008; 87:279 - 90; http://dx.doi.org/10.1016/j.ejcb.2008.02.007; PMID: 18396345
- Hoffmann K, Sperling K, Olins AL, Olins DE. The granulocyte nucleus and lamin B receptor: avoiding the ovoid. Chromosoma 2007; 116:227 - 35; http://dx.doi.org/10.1007/s00412-007-0094-8; PMID: 17245605