
Abstract
Chromatin loops are pervasive and permit the tight compaction of DNA within the confined space of the nucleus. Looping enables distal genes and DNA elements to engage in chromosomal contact, to form multigene complexes. Advances in biochemical and imaging techniques reveal that loop-mediated contact is strongly correlated with transcription of interacting DNA. However, these approaches only provide a snapshot of events and therefore are unable to reveal the dynamics of multigene complex assembly. This highlights the necessity to develop single cell-based assays that provide single molecule resolution, and are able to functionally interrogate the role of chromosomal contact on gene regulation. To this end, high-resolution single cell imaging regimes, combined with genome editing approaches, are proving to be pivotal to advancing our understanding of loop-mediated dynamics.
Introduction
In each mammalian cell, chromosomal DNA is compacted by extensive looping and reduced in size ~500 000 fold to be housed inside each diploid nucleus. Due to this tight compaction, regions of looped chromatin are permitted to interact or “kiss.”Citation1 Chromosomal contact is strongly correlated with the transcriptional activity of interacting DNA elements.Citation2-Citation7 Therefore, the physical contact between looped chromatin appears to be fundamental to gene regulation. However, whether the “kissing” of chromatin is a direct cause, or consequence, of transcription is a long-standing question in molecular biology and the subject of intense scrutiny.
By identifying sites where chromatin interacts, the population-based chromosome conformation capture (3C) and derivative (4C, 5C, Hi-C, etc.) technologies () allow us to “peer” into the highly interconnected environment of the nucleus and to characterize the global interactome (see ref. Citation8 for review). 3C-based techniques involve crosslinking of adjacent chromatin, followed by shearing of DNA, by digestion or sonication.Citation9,Citation10 Proximal genomic regions are then ligated to each other, so interacting DNA elements can be recovered by sequencing or microarray analysis.Citation9,Citation11 In-depth analyses of chromatin interaction data sets reveal a large heterogeneity that infers significant cell-to-cell variability.Citation3,Citation6 Thus, to decrypt the granularity of these events, the physical juxtaposition of interacting loci is verified at a single-cell level by fluorescent in situ hybridization (FISH). This microscopy-based assay, which can target RNA or DNA (), reveals proximal FISH foci in a subset of the population.Citation3,Citation4,Citation6 Collectively, 3C and FISH data suggest that a special pool of cells in the population possesses the correct chromatin organization to permit specific chromosomal interactions.
Table 1. Techniques for unraveling chromatin interactions
These observations raise several important questions. (1) Does every cell possess a unique organization of its chromosomes, or alternatively, can each cell across a population adopt a similar spatial organization of their genome? (2) If it is indeed only a small percentage of cells that possess the appropriate chromatin arrangement to permit chromosomal contact, how do these cells establish optimal chromatin organization? (3) What are the general principles governing loop-mediated transcriptional regulation? In order to accurately resolve these fundamental questions we need to move from indirect approaches conducted on millions of cells, to single cell functional assays conducted systematically across large populations of cells. This requires the ability to develop assays capable of discretely interfering with chromatin architecture while simultaneously monitoring the position of multiple DNA elements, as well as their associated transcription. Moreover, the ability to perform these assays in live cells will be essential to uncover the complexities and dynamic nature of such events. Here we summarize some of the latest advances in biochemical and microscopic assays that are profoundly altering how we understand gene regulation in the 3D space of the nucleus.
Higher Order Genome Organization Mediates Reproducible Chromatin Interactions
Chromosomal contact between genes and their enhancer elements are the best studied of all genomic interactions. Genome-wide chromatin interactome studies reveal that enhancer-promoter interactions are pervasive and may occur in cis even over large genomic distances, or in trans across different chromosomes.Citation12,Citation13 One of the most well characterized examples is the enhancer-promoter model between the locus control region (LCR), and the promoter of the β-globin gene.Citation2 By physically contacting the promoter, the LCR has been shown to induce the expression of the β-globin gene.Citation14 Other similar enhancer-promoter interactions have also been described, such as looping between distal regulators and the α-globin locus,Citation15 the CFTR locus,Citation16 the c-MYC locus,Citation17 and TNFα-responsive genes.Citation18 Interestingly, recent studies demonstrate that enhancer-promoter contacts are not dynamic, but rather are formed prior to gene activation.Citation19-Citation21 Although it is unclear whether this is a general feature of chromatin organization these studies indicate that, once formed, specific chromosomal interactions may be highly stable. Yet, at a fundamental level, it is still poorly understood how these distal elements are able to locate their “interacting” partners in the highly crowded environment of the nucleus.
In situ hybridization assays that “paint” entire chromosomes reveal that chromosomal DNA is not haphazardly intertwined in the interphase nucleus, but rather divided into distinct territories ().Citation21 Moreover, the juxtaposition of neighboring chromosome territories is non-random, with smaller chromosomes preferentially occupying the nuclear center, while, larger chromosomes are more commonly located near the nuclear periphery.Citation22,Citation23 Territories are not insulated from each other, and regions of chromatin have been observed to “roam” into neighboring territories to engage in chromosomal contact.Citation24 Although, these inter-chromosomal contacts are rare, “intermingling” between pairs of loci is highly correlated with translocation frequency.Citation24 This reveals that the 3D positioning of chromosome territories in the nucleus is a key contributor to genomic structure and function.
Figure 1. Hierarchical chromatin organization may facilitate variegated gene expression. (A) Chromatin is spatially segregated into territories, which are further organized into topological associating domains (TADs). Cis or intra-chromosomal interactions occur within a TAD, while, trans or inter-chromosomal contacts are facilitated by the proximal localization of neighboring TADs. (B) A subset or “jackpot cells” in the population may possess an optimal chromosomal arrangement to permit specific intra- and/or inter-chromosomal contacts. As a consequence of loop-mediated transcriptional regulation, these cells display elevated levels of transcription.Citation3
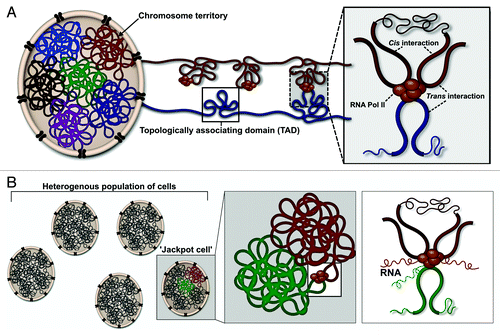
At a population level, 3C-based approaches are able to interrogate chromatin organization to a resolution of ~5 Kb.Citation18 Analysis of Hi-C data sets have uncovered that chromatin is roughly divided into two types of multi-Mb compartments, which can span the same, or different chromosomes.Citation25 Gene-rich and transcriptionally active loci preferentially associate in compartment A and are spatially segregated from gene-poor and transcriptionally silent loci located in compartment B. These compartments are further organized into smaller chromosomal domains, referred to as topologically associating domains (TADs), which range in size from several hundred Kb to ~3 Mb ().Citation26 By constraining the DNA topology, TADs may enhance the probability of loop-mediated interactions occurring. Hence, chromosomal contact, which may occur in cis over many Mb or in trans, would most likely result from the proximal localization of specific TADs on the same or a neighboring chromosome. Thus, this hierarchical mode of chromatin organization may facilitate specific chromosomal interactions, such as those that occur between distal regulatory elements and their target genes.
Do All Cells Possess the Same Genome Connectivity?
Within a TAD, or at the interface between TADs, chromosomal looping may also bring co-regulated genes into close proximity to permit chromosomal contact. These interactions are proposed to occur at discrete foci of active, hyper-phosphorylated RNA Pol II that have been referred to as multigene complexesCitation6 or transcription factories.Citation27 Numerous studies have demonstrated that loop-mediated contact between coregulated genes coincides with alterations in the transcription of interacting genes, including those encoding globins,Citation2 interleukins,Citation28 Hox genes,Citation29 and estrogen-responsive genesCitation10 as well as TNFα- and TGFβ-responsive genes.Citation30 Over 95% of genes engaged in promoter-mediated chromosomal contact in multigene complexes are associated with transcriptional activity.Citation6,Citation7 Therefore, chromosomal contact between co-regulated genes in multigene complexes appears to be a critical component of transcriptional regulation.
The subdivision of the genome into subchromosomal domains, or TADs, represents a highly conserved feature of chromosomal organization.Citation26,Citation31 This suggests that each cell across a population would possess a very similar arrangement of its chromosomes. Paradoxically, traditional in situ studies reveal a large degree of heterogeneity across a population of genotypically identical cells. For example, co-localization of FISH foci between the LCR and the β-globin gene is only observed in 5–10% of cells across a population.Citation3 In these “jackpot cells,” LCR-mediated chromosomal interactions induce β-globin expression ~100-fold (). This variability in β-globin gene transcript levels, or spatially variegated gene expression, may account for the stochastic effects of transcription observed across a population of identical cells.Citation3 In a comparable manner to the LCR/β-globin chromosomal contact, interacting genes in multigene complexes are only detected in a fraction of the population.Citation4-Citation6 Clearly, interrogating chromosomal interactions both at the level of a single cell and single molecule, is imperative to the interpretation of global interactome studies.
Recently, Nagano et al. adapted the Hi-C protocol to allow the simultaneous detection of thousands of contacts in the same nucleus.Citation32 Interestingly, analyses of single cell Hi-C data reveals that intra-chromosomal contacts are highly consistent between cells. This provides strong evidence that, within a chromosome territory, the 3D compaction of chromatin is highly ordered. Conversely, inter-chromosomal contacts were highly variable between individual cells.Citation32 Although the low sample number needs to be considered, this suggests that there may be considerable variation in the 3D organization of chromosomal territories across a population of cells. Clearly, these single cell interactome maps are critical to unravel the degree of variability in chromatin folding between individual cells. However, as these assays are performed in fixed cells, they are still unable to directly address how, or when, loop-mediated contact influences transcription of interacting genes.
What are the Rules Governing Chromosomal Contact?
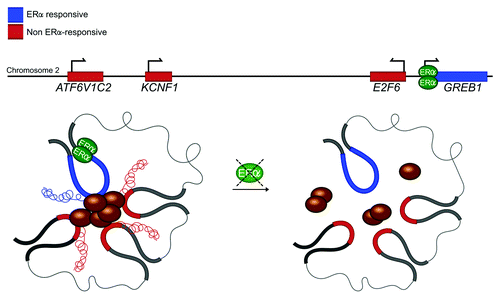
Through physical contact, enhancers are proposed to promote transcription by increasing the concentration of transcriptional regulators at the promoter of their “interacting” partner.Citation8,Citation14 There are two opposing views of how enhancer-promoter regulation occurs: recent evidence shows that these interactions are preformed,Citation18 whereas alternative evidence favors the view that these interactions are dynamically established.Citation14,Citation33 For example, through the looping out of ~50 Kb of intervening chromatin fiber, the LCR was shown to bring regulators to the promoter of the β-globin gene, to initiate its transcription.Citation14 Similarly, upon induction by viral infection, NFκB is delivered to the promoter of inducible genes via inter-chromosomal contact.Citation33 This raises an interesting question; does loop-mediated contact enhance the transcription of interacting genes in a multigene complex? Chromatin interaction analysis with paired end tags (ChIA-PET) revealed that the GREB1 gene engages in multiple intrachromosomal interactions to form a multigene complex.Citation6 Of the 4 interacting genes investigated, only GREB1 transcription is activated by the estrogen receptor-α (ERα).Citation6 Interestingly, the global knockout of ERα disrupted not only GREB1 transcription but the transcription of other 3 non-ERα responsive genes (). Thus, in an analogous manner to enhancer-promoter interactions, this study suggests that the GREB1 gene loop may be a prerequisite for the transcriptional activation of other interacting members of the multigene complex.
Figure 2. Chromosomal contact in multigene complexes may promote transcription of interacting genes. GREB1, an ERα-inducible gene, is cotranscribed with non-ERα inducible genes in the same multigene complex. siRNA knockdown of ERα, does not only abrogate GREB1 transcription, but also the other interacting genes. In addition, chromosomal interactions between GREB1 and non-ERα inducible genes are abrogated.Citation6
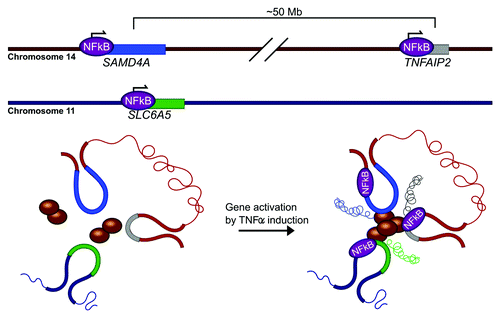
TNFα, a major proinflammatory cytokine, is another stimulus that induces the coordinated assembly of coregulated genes in NFκB-dependent multigene complexes.Citation30 SAMD4A, a ~221 Kb gene on chromosome 14, is rapidly switched on by TNFα in primary human umbilical vein endothelial cells (HUVEC).Citation34 4C analysis reveals that prior to stimulation with TNFα, SAMD4A seldom interacts with other genes.Citation30 After activation by TNFα, SAMD4A interacts with multiple coregulated genes to form a multigene complex.Citation30 TNFAIP2, a gene located on the same chromosome but ~50 Mb downstream, and SLC6A5 on chromosome 11, are two well-characterized interacting partners of SAMD4A ().Citation30 siRNA approaches cannot be utilized to interrogate loop-mediated dynamics in the SAMD4A/TNFAIP2/SLC6A5 multigene complex, as all three genes are activated by the same transcription factor ().Citation4 This exposes the necessity to develop functional assays able to discretely alter gene loop topology, without the global ablation of transcriptional regulators.
Figure 3. TNFα induces the formation of an NFκB-regulated multigene complex. In response to TNFα stimulation, NFκB translocates into nucleus to activate hundreds of responsive genes. The coordinated transcription of these coregulated genes appears to occur in multigene complexes. One such multigene complex that has been extensively characterized by 3C and FISH-based approaches includes SAMD4A (blue), TNFAIP2 (gray), and SLC6A5 (green).Citation4 Prior to activation, DNA FISH reveals that, in a fraction of the population, these three genes are in close proximity, but not physical contact.Citation46 After activation by TNFα, the promoters of the three genes associate via RNA Pol II to form a multigene complex.
Site-specific DNA binding tools, including zinc fingers (ZF), transcription activator-like effectors (TALE) and the Type II CRISPR system (clustered regularly interspaced short palindromic repeats) are emerging as powerful ways to discretely interrogate gene regulation ().Citation35-Citation38 Fusion of the FokI endonuclease to a ZF or TALE DNA binding region, or the CRISPR-associated protein Cas9, enables these tools to induce site-specific double strand breaks (DSB).Citation37-Citation39 Two features of FokI have made it an ideal enzyme to be used as a component of a site-specific nuclease. First, the well-characterized modular nature of the enzymeCitation40-Citation43 has allowed just its catalytic domain to be fused with customizable DNA binding proteins such as zinc fingers and TALEs. Second, since it acts as a dimer,Citation44 two nucleases binding in opposite orientations on adjacent sequences on the genomic target are necessary to generate a DSB, essentially enhancing its specificity.
Figure 4. Chromosomal contact permits transcription of coregulated genes. (A) Site-specific nucleases are powerful tools to discretely interrogate 3D gene regulation. Transcription activator-like effectors nucleases (TALENs) can be designed to target almost any DNA sequence, whereas zinc finger nuclease (ZFN) design is limited by the availability of triplet motifs, and CRISPR/Cas9 (RGEN) targeting requires the NGG or NAG protospacer adjacent motif (PAM). The disruption of gene loops can be visualized by immunofluorescent labeling of H2A.X pSer139. (B) TALEN-mediated disruption of sites of chromosomal contact reveals a hierarchical regulation between these three genes. (C) Repairing the disrupted SAMD4A gene loop restores transcription of interacting genes. Bar, 5μm.
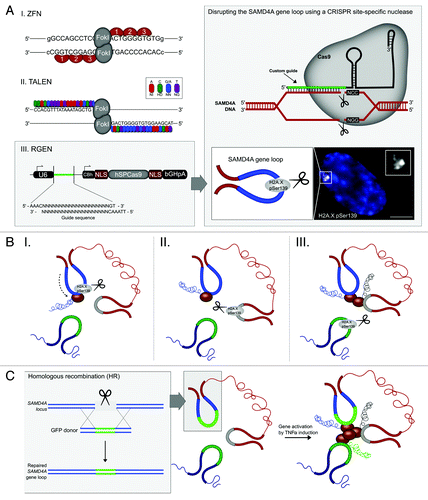
Typically, these site-specific nucleases are used in “cut and paste” experiments, whereby the cell’s own repair responses, nonhomologous end-joining (NHEJ) or homologous recombination (HR), are exploited to repair the DSB.Citation45 Recently, we developed a novel single cell microscopy-based assay, to address the role of loop-mediated contact on transcription of interacting genes in the SAMD4A/TNFAIP2/SLC6A5 multigene complex.Citation46 In the assay, TALE nucleases (TALENs) were used to induce a DSB at the approximate site within each of these three gene loops that was established by 3C-technologies to engage in contact.Citation4,Citation46 We have also tested the ability of other site-specific nucleases to disrupt the SAMD4A gene loop (). The DSB, or site of disruption, was detected by immunofluorescent staining of H2A.X pSer139, a factor of the DSB repair process.Citation47 In parallel to the detection of the DSB, transcriptional activity of interacting genes in the multigene complex was determined using intronic RNA FISH. As introns are typically excised and degraded co-transcriptionally, intronic FISH foci represent the transcriptional start site (TSS).Citation34 Consistent with previous studies,Citation4 co-localization between the FISH foci of these three genes was only observed at a low percentage of all alleles across the population (~5%).Citation46 This would suggest that this small fraction of the population represents a “special pool” of primed cells that permit loop-mediated contact.
Using the TALEN single cell assay, we observed that perturbing the SAMD4A gene loop did not alter transcription 5' of the DSB ( section I). However, consistent with other studies,Citation48,Citation49 we observed silencing of SAMD4A transcription 3' of the DSB ( section I). Intriguingly, despite occupying distal genomic locations, the transcription of TNFAIP2 and SLC6A5 were also significantly reduced ( section I). Further, the effect on co-transcription was hierarchical, with the disruption of TNFAIP2 altering SLC6A5 expression, but having no influence on SAMD4A ( section II), whereas perturbing SLC6A5 did not influence SAMD4A or TNFAIP2 transcription ( section III). This suggests a hierarchical assembly between these three genes, whereby TNFAIP2 “collects” RNA Pol II from SAMD4A, which are then able to recruit SLC6A5 to the multigene complex (). Is this ordered assembly of these co-regulated genes reliant on chromosomal contact? In our study, the TALEN-induced DSB is visualized by immunofluorescent labeling of Ser139 phosphorylated H2A.X.Citation46 However, a large number of other proteins involved in the repair process are also recruited to the DSB site that may have several implications for loop-mediated contact. One possibility could be that chromosomal contacts between SAMD4A, TNFAIP2 and/or SLC6A5 may still occur, but are “bridged” via the repair complex ( section I). This may occlude the ability of SAMD4A to “deliver” Pol II to other interacting genes. DSB induction has been shown to enhance the mobility of damaged chromatin (see ref. Citation50 for review). Therefore, another explanation could be that, due to increased movement, the probability that the disrupted gene loop will be able to interact with other loci is significantly reduced ( section II). Alternatively, it is unknown how the DSB, and associated repair factors, influence gene topology. Therefore, we speculate that the DSB may prevent loop-mediated contact, by destabilizing, or “collapsing,” the disrupted loop ( section III). Nonetheless, in all of these scenarios, gene loop topology and chromosomal contact are strongly implicated in the regulation of multigene complex assembly. Therefore, our study provides the first direct evidence that chromatin contacts may be a cause, and not simply a consequence of transcription.
Figure 5. Hypothetical model for hierarchical assembly of a NFκB-dependent multigene complex. (A) The single-cell TALEN/RNA FISH assay reveals a hierarchical recruitment to co-regulated genes in a multigene complex. This suggests that SAMD4A (blue) collects TNFAIP2 (gray) which in turn recruit SLC6A5 (green) (B) Mechanisms depicting how the DSB may abrogate transcription of interacting genes. (I) The repair machinery recruited to SAMD4A may interpose between interacting genes, occluding the ability of Pol II to access TNFAIP2 and SLC6A5. (II) The DSB may enhance the mobility of SAMD4A, greatly reducing its capacity to engage in loop-mediated contact with TNFAIP2 and SLC6A5. (III) The recruitment of the repair machinery to the DSB may “collapse” the SAMD4A gene loop, and abrogate any associated chromosomal contact.
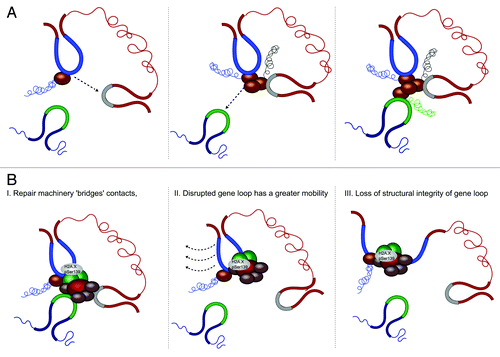
These observations beg a new question, what are the protein and/or sequence specific factors driving the hierarchy? Replacing the region of contact with a GFP sequence was sufficient to restore the transcription of interacting genes ().Citation46 This suggests that, while the sequence with the first intron that is engaging in loop-mediated contact is not determining the hierarchy, the topological framework of the multigene complex is a requirement for 3D transcriptional regulation. A recent study revealed that the co-localization of these NFκB-regulated genes may be driven by promoter-specific sequence elements.Citation51 Therefore, discrete repair experiments within the endogenous promoters of these genes may unveil whether promoter-specific sequence elements are involved.
What are the Dynamics of Loop-Mediated Chromosomal Contact?
Chromosomal interactions detected by 3C and FISH-based approaches represent a snapshot of chromatin organization across a heterogeneous population of cells. Therefore, live microscopy-based assays are indispensable to chromatin interactome studies. Historically, co-localized RNA or DNA FISH foci are used to validate chromosomal interactions that are detected by 3C-based approaches.Citation2,Citation4,Citation5 Overlapping FISH foci between SAMD4A, TNFAIP2, and SLC6A5 are only observed at 5% of all alleles.Citation46 However, when we disrupted the SAMD4A gene loop we observed a significant decrease in TNFAIP2 and SLC6A5 expression (at more than 5% of alleles). As the half-life of the SAMD4A intron is ~5 min,Citation34 it is possible that the intronic FISH foci may not always represent the exact site of transcription. Therefore, we speculate that percentage co-localization as measured by intronic RNA FISH may not always be an accurate measure of “jackpot” or “primed” cells. Live-cell imaging reveals that the movement of individual loci is limited to a 0.5 μm area, which corresponds to the approximate size of a ~1 Mb TAD.Citation52-Citation54 Therefore, each locus constrained within a TAD may be able to interact with every loci located in the same TAD, or even proximal TADs, within the time frame of each cell division.Citation8 Thus, live cell assays are critical to deciphering the principles governing loop-mediated gene regulation.
In fixed cells, light microscopy studies reveal that the co-localization of coregulated genes occurs at discrete foci of RNA Pol II.Citation2 However, these microscopy approaches are unable to accurately discern the dynamics of RNA Pol II/gene loop assembly. Moreover, the resolution of conventional light microscopy is at best, ~200 nm in the x- and y-axes and ~500 nm in the z-axis. Novel super-resolution microscopy approaches such as photo-activated localization microscopy (PALM), stochastic optical reconstruction microscopy (STORM) and selective plane illumination microscopy (SPIM) are able to breach this limit, to achieve sub-diffractive resolution.Citation55 These approaches are based on repetitive stochastic activation and deactivation of fluorescent proteins or probes, allowing the acquisition of super-resolution information and localization information down to 20 nm.Citation55 Recently, through the endogenous labeling of RNA Pol II, Cisse et al. used PALM to study RNA Pol II dynamics in live cells at subdiffractive resolution.Citation56 By replacing the endogenous Pol II catalytic subunit (RPB1) with the photo-switchable fluorescent protein, Dendra2, they were able to show that the clustering of RNA Pol II precedes transcription.Citation56 This study provides critical insight into the dynamics of multigene complex assembly. However, to what extent does the replacement of an endogenous protein subunit with GFP alter the normal functioning of RNA Pol II? Clearly, in order to resolve questions around the rules governing chromosomal contact and transcriptional regulation, requires the ability to perform functional assays at their endogenous locations, and as discretely as possible. To this end, high-resolution single cell imaging regimes combined with genome-editing approaches, including those involving transcriptional activation/repression and live imaging,Citation57-Citation59 will pave the way to a fuller understanding of 3D transcriptional regulation.
Disclosure of Potential Conflicts of Interest
No potential conflict of interest was disclosed.
Acknowledgments
We thank S Barichievy, J Scholefield, and O Bischof for comments on the manuscript. This research is supported by grant PG-V2KYP07 from the Council for Industrial and Scientific Research (CSIR, South Africa) and a grant from the Emerging Research Area Program of The Department of Science and Technology (DST, South Africa) and grant PTDC/SAU-GMG/115652/2009 from the Fundação para a Ciência e a Tecnologia (FCT, Portugal) all to M.M.
References
- Kioussis D. Gene regulation: kissing chromosomes. Nature 2005; 435:579 - 80; http://dx.doi.org/10.1038/435579a; PMID: 15931207
- Osborne CS, Chakalova L, Brown KE, Carter D, Horton A, Debrand E, Goyenechea B, Mitchell JA, Lopes S, Reik W, et al. Active genes dynamically colocalize to shared sites of ongoing transcription. Nat Genet 2004; 36:1065 - 71; http://dx.doi.org/10.1038/ng1423; PMID: 15361872
- Noordermeer D, de Wit E, Klous P, van de Werken H, Simonis M, Lopez-Jones M, Eussen B, de Klein A, Singer RH, de Laat W. Variegated gene expression caused by cell-specific long-range DNA interactions. Nat Cell Biol 2011; 13:944 - 51; http://dx.doi.org/10.1038/ncb2278; PMID: 21706023
- Papantonis A, Larkin JD, Wada Y, Ohta Y, Ihara S, Kodama T, Cook PR. Active RNA polymerases: mobile or immobile molecular machines?. PLoS Biol 2010; 8:e1000419; http://dx.doi.org/10.1371/journal.pbio.1000419; PMID: 20644712
- Schoenfelder S, Sexton T, Chakalova L, Cope NF, Horton A, Andrews S, Kurukuti S, Mitchell JA, Umlauf D, Dimitrova DS, et al. Preferential associations between co-regulated genes reveal a transcriptional interactome in erythroid cells. Nat Genet 2010; 42:53 - 61; http://dx.doi.org/10.1038/ng.496; PMID: 20010836
- Li G, Ruan X, Auerbach RK, Sandhu KS, Zheng M, Wang P, Poh HM, Goh Y, Lim J, Zhang J, et al. Extensive promoter-centered chromatin interactions provide a topological basis for transcription regulation. Cell 2012; 148:84 - 98; http://dx.doi.org/10.1016/j.cell.2011.12.014; PMID: 22265404
- Sandhu KS, Li G, Poh HM, Quek YL, Sia YY, Peh SQ, Mulawadi FH, Lim J, Sikic M, Menghi F, et al. Large-scale functional organization of long-range chromatin interaction networks. Cell Rep 2012; 2:1207 - 19; http://dx.doi.org/10.1016/j.celrep.2012.09.022; PMID: 23103170
- Gibcus JH, Dekker J. The hierarchy of the 3D genome. Mol Cell 2013; 49:773 - 82; http://dx.doi.org/10.1016/j.molcel.2013.02.011; PMID: 23473598
- Dekker J, Rippe K, Dekker M, Kleckner N. Capturing chromosome conformation. Science 2002; 295:1306 - 11; http://dx.doi.org/10.1126/science.1067799; PMID: 11847345
- Fullwood MJ, Liu MH, Pan YF, Liu J, Xu H, Mohamed YB, Orlov YL, Velkov S, Ho A, Mei PH, et al. An oestrogen-receptor-alpha-bound human chromatin interactome. Nature 2009; 462:58 - 64; http://dx.doi.org/10.1038/nature08497; PMID: 19890323
- Simonis M, Klous P, Splinter E, Moshkin Y, Willemsen R, de Wit E, van Steensel B, de Laat W. Nuclear organization of active and inactive chromatin domains uncovered by chromosome conformation capture-on-chip (4C). Nat Genet 2006; 38:1348 - 54; http://dx.doi.org/10.1038/ng1896; PMID: 17033623
- Sanyal A, Lajoie BR, Jain G, Dekker J. The long-range interaction landscape of gene promoters. Nature 2012; 489:109 - 13; http://dx.doi.org/10.1038/nature11279; PMID: 22955621
- Zhang Y, Wong CH, Birnbaum RY, Li G, Favaro R, Ngan CY, Lim J, Tai E, Poh HM, Wong E, et al. Chromatin connectivity maps reveal dynamic promoter-enhancer long-range associations. Nature 2013; 504:306 - 10; http://dx.doi.org/10.1038/nature12716; PMID: 24213634
- Deng W, Lee J, Wang H, Miller J, Reik A, Gregory PD, Dean A, Blobel GA. Controlling long-range genomic interactions at a native locus by targeted tethering of a looping factor. Cell 2012; 149:1233 - 44; http://dx.doi.org/10.1016/j.cell.2012.03.051; PMID: 22682246
- Vernimmen D, De Gobbi M, Sloane-Stanley JA, Wood WG, Higgs DR. Long-range chromosomal interactions regulate the timing of the transition between poised and active gene expression. EMBO J 2007; 26:2041 - 51; http://dx.doi.org/10.1038/sj.emboj.7601654; PMID: 17380126
- Gheldof N, Smith EM, Tabuchi TM, Koch CM, Dunham I, Stamatoyannopoulos JA, Dekker J. Cell-type-specific long-range looping interactions identify distant regulatory elements of the CFTR gene. Nucleic Acids Res 2010; 38:4325 - 36; http://dx.doi.org/10.1093/nar/gkq175; PMID: 20360044
- Wright JB, Brown SJ, Cole MD. Upregulation of c-MYC in cis through a large chromatin loop linked to a cancer risk-associated single-nucleotide polymorphism in colorectal cancer cells. Mol Cell Biol 2010; 30:1411 - 20; http://dx.doi.org/10.1128/MCB.01384-09; PMID: 20065031
- Jin F, Li Y, Dixon JR, Selvaraj S, Ye Z, Lee AY, Yen CA, Schmitt AD, Espinoza CA, Ren B. A high-resolution map of the three-dimensional chromatin interactome in human cells. Nature 2013; 503:290 - 4; PMID: 24141950
- Melo CA, Drost J, Wijchers PJ, van de Werken H, de Wit E, Oude Vrielink JA, Elkon R, Melo SA, Léveillé N, Kalluri R, et al. eRNAs are required for p53-dependent enhancer activity and gene transcription. Mol Cell 2013; 49:524 - 35; http://dx.doi.org/10.1016/j.molcel.2012.11.021; PMID: 23273978
- Phillips-Cremins JE, Sauria ME, Sanyal A, Gerasimova TI, Lajoie BR, Bell JS, Ong CT, Hookway TA, Guo C, Sun Y, et al. Architectural protein subclasses shape 3D organization of genomes during lineage commitment. Cell 2013; 153:1281 - 95; http://dx.doi.org/10.1016/j.cell.2013.04.053; PMID: 23706625
- Cremer T, Cremer M. Chromosome territories. Cold Spring Harb Perspect Biol 2010; 2:a003889; http://dx.doi.org/10.1101/cshperspect.a003889; PMID: 20300217
- Croft JA, Bridger JM, Boyle S, Perry P, Teague P, Bickmore WA. Differences in the localization and morphology of chromosomes in the human nucleus. J Cell Biol 1999; 145:1119 - 31; http://dx.doi.org/10.1083/jcb.145.6.1119; PMID: 10366586
- Cremer T, Cremer M, Dietzel S, Müller S, Solovei I, Fakan S. Chromosome territories--a functional nuclear landscape. Curr Opin Cell Biol 2006; 18:307 - 16; http://dx.doi.org/10.1016/j.ceb.2006.04.007; PMID: 16687245
- Branco MR, Pombo A. Intermingling of chromosome territories in interphase suggests role in translocations and transcription-dependent associations. PLoS Biol 2006; 4:e138; http://dx.doi.org/10.1371/journal.pbio.0040138; PMID: 16623600
- Lieberman-Aiden E, van Berkum NL, Williams L, Imakaev M, Ragoczy T, Telling A, Amit I, Lajoie BR, Sabo PJ, Dorschner MO, et al. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science 2009; 326:289 - 93; http://dx.doi.org/10.1126/science.1181369; PMID: 19815776
- Dixon JR, Selvaraj S, Yue F, Kim A, Li Y, Shen Y, Hu M, Liu JS, Ren B. Topological domains in mammalian genomes identified by analysis of chromatin interactions. Nature 2012; 485:376 - 80; http://dx.doi.org/10.1038/nature11082; PMID: 22495300
- Cook PR. The organization of replication and transcription. Science 1999; 284:1790 - 5; http://dx.doi.org/10.1126/science.284.5421.1790; PMID: 10364545
- Spilianakis CG, Lalioti MD, Town T, Lee GR, Flavell RA. Interchromosomal associations between alternatively expressed loci. Nature 2005; 435:637 - 45; http://dx.doi.org/10.1038/nature03574; PMID: 15880101
- Noordermeer D, Leleu M, Splinter E, Rougemont J, De Laat W, Duboule D. The dynamic architecture of Hox gene clusters. Science 2011; 334:222 - 5; http://dx.doi.org/10.1126/science.1207194; PMID: 21998387
- Papantonis A, Kohro T, Baboo S, Larkin JD, Deng B, Short P, Tsutsumi S, Taylor S, Kanki Y, Kobayashi M, et al. TNFα signals through specialized factories where responsive coding and miRNA genes are transcribed. EMBO J 2012; 31:4404 - 14; http://dx.doi.org/10.1038/emboj.2012.288; PMID: 23103767
- Nora EP, Lajoie BR, Schulz EG, Giorgetti L, Okamoto I, Servant N, Piolot T, van Berkum NL, Meisig J, Sedat J, et al. Spatial partitioning of the regulatory landscape of the X-inactivation centre. Nature 2012; 485:381 - 5; http://dx.doi.org/10.1038/nature11049; PMID: 22495304
- Nagano T, Lubling Y, Stevens TJ, Schoenfelder S, Yaffe E, Dean W, Laue ED, Tanay A, Fraser P. Single-cell Hi-C reveals cell-to-cell variability in chromosome structure. Nature 2013; 502:59 - 64; http://dx.doi.org/10.1038/nature12593; PMID: 24067610
- Apostolou E, Thanos D. Virus Infection Induces NF-kappaB-dependent interchromosomal associations mediating monoallelic IFN-beta gene expression. Cell 2008; 134:85 - 96; http://dx.doi.org/10.1016/j.cell.2008.05.052; PMID: 18614013
- Wada Y, Ohta Y, Xu M, Tsutsumi S, Minami T, Inoue K, Komura D, Kitakami J, Oshida N, Papantonis A, et al. A wave of nascent transcription on activated human genes. Proc Natl Acad Sci U S A 2009; 106:18357 - 61; http://dx.doi.org/10.1073/pnas.0902573106; PMID: 19826084
- Kim YG, Cha J, Chandrasegaran S. Hybrid restriction enzymes: zinc finger fusions to Fok I cleavage domain. Proc Natl Acad Sci U S A 1996; 93:1156 - 60; http://dx.doi.org/10.1073/pnas.93.3.1156; PMID: 8577732
- Bibikova M, Beumer K, Trautman JK, Carroll D. Enhancing gene targeting with designed zinc finger nucleases. Science 2003; 300:764; http://dx.doi.org/10.1126/science.1079512; PMID: 12730594
- Li T, Huang S, Jiang WZ, Wright D, Spalding MH, Weeks DP, Yang B. TAL nucleases (TALNs): hybrid proteins composed of TAL effectors and FokI DNA-cleavage domain. Nucleic Acids Res 2011; 39:359 - 72; http://dx.doi.org/10.1093/nar/gkq704; PMID: 20699274
- Jinek M, Chylinski K, Fonfara I, Hauer M, Doudna JA, Charpentier E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012; 337:816 - 21; http://dx.doi.org/10.1126/science.1225829; PMID: 22745249
- Christian M, Cermak T, Doyle EL, Schmidt C, Zhang F, Hummel A, Bogdanove AJ, Voytas DF. Targeting DNA double-strand breaks with TAL effector nucleases. Genetics 2010; 186:757 - 61; http://dx.doi.org/10.1534/genetics.110.120717; PMID: 20660643
- Li L, Wu LP, Chandrasegaran S. Functional domains in Fok I restriction endonuclease. Proc Natl Acad Sci U S A 1992; 89:4275 - 9; http://dx.doi.org/10.1073/pnas.89.10.4275; PMID: 1584761
- Li L, Wu LP, Clarke R, Chandrasegaran S. C-terminal deletion mutants of the FokI restriction endonuclease. Gene 1993; 133:79 - 84; http://dx.doi.org/10.1016/0378-1119(93)90227-T; PMID: 8224897
- Li L, Chandrasegaran S. Alteration of the cleavage distance of Fok I restriction endonuclease by insertion mutagenesis. Proc Natl Acad Sci U S A 1993; 90:2764 - 8; http://dx.doi.org/10.1073/pnas.90.7.2764; PMID: 8464886
- Kim YG, Li L, Chandrasegaran S. Insertion and deletion mutants of FokI restriction endonuclease. J Biol Chem 1994; 269:31978 - 82; PMID: 7989374
- Smith J, Bibikova M, Whitby FG, Reddy AR, Chandrasegaran S, Carroll D. Requirements for double-strand cleavage by chimeric restriction enzymes with zinc finger DNA-recognition domains. Nucleic Acids Res 2000; 28:3361 - 9; http://dx.doi.org/10.1093/nar/28.17.3361; PMID: 10954606
- Hoeijmakers JHJ. Genome maintenance mechanisms for preventing cancer. Nature 2001; 411:366 - 74; http://dx.doi.org/10.1038/35077232; PMID: 11357144
- Fanucchi S, Shibayama Y, Burd S, Weinberg MS, Mhlanga MM. Chromosomal contact permits transcription between coregulated genes. Cell 2013; 155:606 - 20; http://dx.doi.org/10.1016/j.cell.2013.09.051; PMID: 24243018
- Rogakou EP, Boon C, Redon C, Bonner WM. Megabase chromatin domains involved in DNA double-strand breaks in vivo. J Cell Biol 1999; 146:905 - 16; http://dx.doi.org/10.1083/jcb.146.5.905; PMID: 10477747
- Kruhlak M, Crouch EE, Orlov M, Montaño C, Gorski SA, Nussenzweig A, Misteli T, Phair RD, Casellas R. The ATM repair pathway inhibits RNA polymerase I transcription in response to chromosome breaks. Nature 2007; 447:730 - 4; http://dx.doi.org/10.1038/nature05842; PMID: 17554310
- Shanbhag NM, Rafalska-Metcalf IU, Balane-Bolivar C, Janicki SM, Greenberg RA. ATM-dependent chromatin changes silence transcription in cis to DNA double-strand breaks. Cell 2010; 141:970 - 81; http://dx.doi.org/10.1016/j.cell.2010.04.038; PMID: 20550933
- Dion V, Gasser SM. Chromatin movement in the maintenance of genome stability. Cell 2013; 152:1355 - 64; http://dx.doi.org/10.1016/j.cell.2013.02.010; PMID: 23498942
- Larkin JD, Papantonis A, Cook PR. Promoter type influences transcriptional topography by targeting genes to distinct nucleoplasmic sites. J Cell Sci 2013; 126:2052 - 9; http://dx.doi.org/10.1242/jcs.123653; PMID: 23444365
- Abney JR, Cutler B, Fillbach ML, Axelrod D, Scalettar BA. Chromatin dynamics in interphase nuclei and its implications for nuclear structure. J Cell Biol 1997; 137:1459 - 68; http://dx.doi.org/10.1083/jcb.137.7.1459; PMID: 9199163
- Marshall WF, Straight A, Marko JF, Swedlow J, Dernburg A, Belmont A, Murray AW, Agard DA, Sedat JW. Interphase chromosomes undergo constrained diffusional motion in living cells. Curr Biol 1997; 7:930 - 9; http://dx.doi.org/10.1016/S0960-9822(06)00412-X; PMID: 9382846
- Heun P, Laroche T, Shimada K, Furrer P, Gasser SM. Chromosome dynamics in the yeast interphase nucleus. Science 2001; 294:2181 - 6; http://dx.doi.org/10.1126/science.1065366; PMID: 11739961
- Henriques R, Griffiths C, Hesper Rego E, Mhlanga MM. PALM and STORM: unlocking live-cell super-resolution. Biopolymers 2011; 95:322 - 31; http://dx.doi.org/10.1002/bip.21586; PMID: 21254001
- Cisse II, Izeddin I, Causse SZ, Boudarene L, Senecal A, Muresan L, Dugast-Darzacq C, Hajj B, Dahan M, Darzacq X. Real-time dynamics of RNA polymerase II clustering in live human cells. Science 2013; 341:664 - 7; http://dx.doi.org/10.1126/science.1239053; PMID: 23828889
- Chen B, Gilbert LA, Cimini BA, Schnitzbauer J, Zhang W, Li GW, Park J, Blackburn EH, Weissman JS, Qi LS, et al. Dynamic imaging of genomic loci in living human cells by an optimized CRISPR/Cas system. Cell 2013; 155:1479 - 91; http://dx.doi.org/10.1016/j.cell.2013.12.001; PMID: 24360272
- Miyanari Y, Ziegler-Birling C, Torres-Padilla ME. Live visualization of chromatin dynamics with fluorescent TALEs. Nat Struct Mol Biol 2013; 20:1321 - 4; http://dx.doi.org/10.1038/nsmb.2680; PMID: 24096363
- Gilbert LA, Larson MH, Morsut L, Liu Z, Brar GA, Torres SE, Stern-Ginossar N, Brandman O, Whitehead EH, Doudna JA, et al. CRISPR-mediated modular RNA-guided regulation of transcription in eukaryotes. Cell 2013; 154:442 - 51; http://dx.doi.org/10.1016/j.cell.2013.06.044; PMID: 23849981
- Dostie J, Richmond TA, Arnaout RA, Selzer RR, Lee WL, Honan TA, Rubio ED, Krumm A, Lamb J, Nusbaum C, et al. Chromosome Conformation Capture Carbon Copy (5C): a massively parallel solution for mapping interactions between genomic elements. Genome Res 2006; 16:1299 - 309; http://dx.doi.org/10.1101/gr.5571506; PMID: 16954542
- Horike S, Cai S, Miyano M, Cheng JF, Kohwi-Shigematsu T. Loss of silent-chromatin looping and impaired imprinting of DLX5 in Rett syndrome. Nat Genet 2005; 37:31 - 40; PMID: 15608638
- Langer-Safer PR, Levine M, Ward DC. Immunological method for mapping genes on Drosophila polytene chromosomes. Proc Natl Acad Sci U S A 1982; 79:4381 - 5; http://dx.doi.org/10.1073/pnas.79.14.4381; PMID: 6812046
- Bienko M, Crosetto N, Teytelman L, Klemm S, Itzkovitz S, van Oudenaarden A. A versatile genome-scale PCR-based pipeline for high-definition DNA FISH. Nat Methods 2013; 10:122 - 4; http://dx.doi.org/10.1038/nmeth.2306; PMID: 23263692
- Cremer T, Cremer M. Chromosome territories. Cold Spring Harb Perspect Biol 2010; 2:a003889; http://dx.doi.org/10.1101/cshperspect.a003889; PMID: 20300217
- Raj A, van den Bogaard P, Rifkin SA, van Oudenaarden A, Tyagi S. Imaging individual mRNA molecules using multiple singly labeled probes. Nat Methods 2008; 5:877 - 9; http://dx.doi.org/10.1038/nmeth.1253; PMID: 18806792
- Levesque MJ, Raj A. Single-chromosome transcriptional profiling reveals chromosomal gene expression regulation. Nat Methods 2013; 10:246 - 8; http://dx.doi.org/10.1038/nmeth.2372; PMID: 23416756
- Cong L, Ran FA, Cox D, Lin S, Barretto R, Habib N, Hsu PD, Wu X, Jiang W, Marraffini LA, et al. Multiplex genome engineering using CRISPR/Cas systems. Science 2013; 339:819 - 23; http://dx.doi.org/10.1126/science.1231143; PMID: 23287718
- Mali P, Yang L, Esvelt KM, Aach J, Guell M, DiCarlo JE, Norville JE, Church GM. RNA-guided human genome engineering via Cas9. Science 2013; 339:823 - 6; http://dx.doi.org/10.1126/science.1232033; PMID: 23287722