
Abstract
The nuclear lamina (NL) is thought to aid in the spatial organization of interphase chromosomes by providing an anchoring platform for hundreds of large genomic regions named lamina associated domains (LADs). Recently, a new live-cell imaging approach demonstrated directly that LAD-NL interactions are dynamic and in part stochastic. Here we discuss implications of these new findings and introduce Lamin A and BAF as potential modulators of stochastic LAD positioning.
Introduction
The NL is a thin filamentous meshwork that lines the inner nuclear membrane (INM) and forms a structural scaffold that is thought to support the spatial organization of the genome. The NL associates with very large genomic regions that range in size from ~10 kb to ~10 Mb and together cover approximately about 40% of the genome. The mammalian genome typically harbors about 1100–1400 of such lamina-associated domains (LADs).Citation1-Citation4
In mammalian somatic cells, the NL consists of two B-type lamins—B1 and B2—and two splice variants of A-type lamin named Lamin A and C. In most cell types, the B-type lamins are almost exclusively located at the NL, while A-type lamins additionally reside in the nucleoplasm.Citation5,Citation6 The role of this nucleoplasmic pool of Lamin A/C remains largely enigmatic.
Besides lamins, the NL contains many other proteins. One of these is the barrier-to-autointegration factor (BAF, encoded by the BANF1 gene), a ~10 kDa evolutionary conserved protein that associates with the NL through interaction with LEM-domain containing proteins of the INM. BAF binds non-specifically to DNACitation7,Citation8 and chromatin.Citation9,Citation10 One possibility is that BAF acts as a “bridging” factor that connects chromatin to the NL, but direct evidence for this has been lacking so far. Like Lamin A, BAF is partially nucleoplasmatic,Citation11 raising the question whether it binds to chromatin at the NL, in the nuclear interior, or both. In fact, it is not known whether BAF interacts with LADs at all.
Recently we reported that the interactions of LADs with the NL appear intrinsically stochastic, i.e., they are variable from cell to cell. Here, we discuss and build on these findings and present data in support of a role for Lamin A and BAF in controlling LAD positioning between the nucleoplasm and the NL in single cells.
Stochastic Positioning of LADs
Our laboratory developed the m6A-Tracer technique to specifically mark and trace chromatin that contacts the NL in single cells.Citation12 The method utilizes a GFP-tagged protein module that recognizes m6A on chromatin after deposition of this adenine modification by Dam-Lamin. Thus, any DNA that contacts the NL can be visualized and followed in live cells. Using Dam-Lamin B1 to label DNA in contact with the NL, this approach revealed that during interphase LADs are somewhat mobile but confined to a narrow zone in the immediate vicinity of the NL. However, after mitosis these same LADs are seemingly randomly redistributed throughout the nucleus of the daughter cells, with only a subset returning to the NL. From these and other data we inferred that many LADs do not contact the NL in every single cell, but rather in a stochastic mannerCitation12; after every mitosis, a different subset of LADs is located at the NL. This model is concordant with various microscopy studiesCitation13 and with recent single-cell HiC experiments,Citation14 which revealed that chromosomes adopt remarkably diverse configurations from cell to cell.
Interestingly, we noticed that some of the nucleoplasmic LADs accumulate around nucleoli.Citation12 This is in agreement with the partial overlap between LADs and nucleoli-associated-domains (NADs) as identified by genome-wide mapping.Citation15,Citation16 It thus appears that some LADs can be located either at the NL or at a nucleolus, and that this choice may be random in a population of cells.
Stochastic Chromatin Modification State of LADs
According to genome-wide studies, LADs tend to be enriched in the heterochromatic histone modification H3K9me2.Citation3,Citation17 However, by microscopy and combined ChIP-DamID experiments we observed that H3K9me2 is primarily present on those LADs that are actually located at the NL, while the same LADs carry less of this histone mark when they are located in the nuclear interior.Citation12 By inference, this means that the H3K9me2 status of LADs is also stochastic, and directly linked to their nuclear position.
Additional experiments indicated that H3K9me2 in part drives NL contacts, because inactivation of G9a, the principal enzyme responsible for H3K9 dimethylation, caused a reduction in LAD–NL contact frequencies.Citation12 In line with these findings, H3K9me2 and H3K9me3 were also shown to be involved in targeting the β-globin locus to the NLCitation18 and a similar mechanism was observed in C. elegans.Citation19
Thus, LADs that are stochastically positioned in the nuclear interior do not detectably interact with Lamin B1, and they tend to have reduced H3K9me2 levels. Below we consider the possibility that the nucleoplasmic pools of Lamin A and BAF substitute for these associations, and even compete for the association of LADs with the NL.
Lamin B1, Lamin B2, Lamin A, and BAF Associate with the Same Genomic Regions
Previously we showed that, despite their partially different nuclear localization,Citation20 Lamin B1 and Lamin A interact with the same genomic loci in populations of mouse and human cells.Citation21 An example of the nearly identical DamID patterns is shown for human HT1080 cells in and . For further comparison, we now also mapped the interaction pattern of Lamin B2. Lamin B2 again yielded a highly similar interaction profile ( and ).
Figure 1. Lamin B1, Lamin B2, Lamin A, and BAF bind to the same genomic regions. (A) Interaction profiles of chromosome 2 in HT1080 cells for Lamin B1, Lamin B2, Lamin A, and BAF. Each profile represents the average of two independent experiments. Data for Lamin B1 and Lamin A are from references Citation12 and Citation21. Samples were smoothed with a running median window over 11 probes. The black boxes at the bottom of the graph depict LADs as defined in reference Citation2. (B) Scatterplots of Lamin B1 in relation to: Lamin B2 (top left), Lamin A (top right), and BAF (bottom left). Samples were smoothed with a running median window of 11 probes. Bottom right: genome wide Pearson correlation matrix for all smoothed (as above) samples.
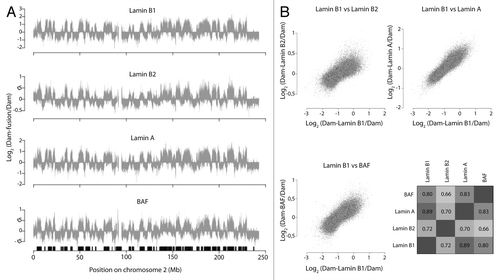
We then performed DamID of BAF, also in HT1080 cells. Remarkably, this protein again showed a genome-wide interaction profile very similar to that of the three lamins ( and ). Thus, BAF also interacts preferentially with LADs.
Lamin A Preferentially Contacts Chromatin around Nucleoli
Due to the stochastic positioning of LADs, an interaction profile of a population of cells does not distinguish between peripheral vs. interior interactions of BAF and Lamin A in single cells. The m6A-Tracer approach indicated that Lamin B1 in interphase exclusively contacts chromatin in the vicinity of the NL, with virtually no signals in the nuclear interior, whereas Lamin A contacts chromatin both at the NL and in the nuclear interior.Citation12
To study the nucleoplasmic m6A-Tracer signals obtained with Dam-Lamin A in more detail, we repeated these experiments with a cell line that stably expresses m6A-Tracer, which makes it easier to score distribution patterns. In this cell line, 20 h after transfection with Dam-Lamin A, m6A-Tracer signals are apparent both at the NL and throughout the nucleoplasm. Interestingly, in 35 ± 7% of the cells a clear accumulation of m6A-Tracer signal appeared around nucleoli (). Interestingly, peri-nucleolar enrichment is not detected with antibody staining of Lamin A/C,Citation12 indicative that the enrichment of the m6A-Tracer around nucleoli likely reflects a accumulation of m6A over time either due to re-current transient and/or more specific chromatin interactions of Lamin A with peri-nucleolar chromatin. In parallel experiments, transfection with Dam-LaminB1 and Dam-LaminB2 did not yield any detectable peri-nucleolar m6A-Tracer staining (), indicating that the cells had not progressed through mitosis since the time of transfection, which would lead to reshuffling of LADs.Citation12
Figure 2. Lamin A interacts with peri-nucleolar chromatin. (A-C) Clonal HT1080 cell line expressing m6A-Tracer construct (in green) transfected with: Dam-Lamin A (left), Lamin B1 (top middle), or Dam-Lamin B2 (top right). The cells were harvested 20 h post transfection. Nucleoli are labeled with an antibody against nucleophosmin (red, bottom panel in A) and the NL is labeled with an antibody against Lamin B1 (blue). The extensive cytoplasmic labeling originates from transfected plasmid molecules that carry m6A and are therefore bound by m6A-Tracer. (D) Percentage of cells that display peri-nucleolar m6ATracer staining. Error bars indicate standard deviation (two independent experiments).
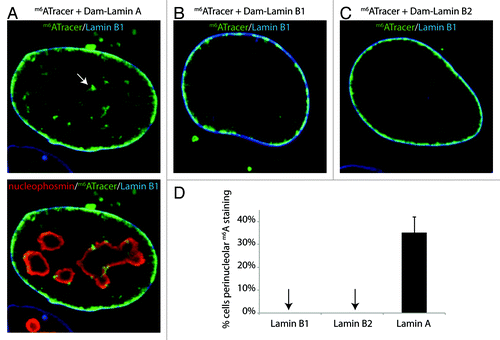
The peri-nucleolar m6A-Tracer enrichments indicate that Lamin A preferentially interacts with certain genomic regions that are positioned adjacent to nucleoli. Combined with the previous findings that (1) Lamin A and Lamins B1/B2 can interact with the same LADs, (2) some LADs are stochastically positioned near nucleoli, and (3) LADs partially overlap with nucleolus-associated-domains (NADs)Citation15,Citation16 ,these results lead us to propose that nucleolus-associated LADs interact with Lamin A. We note that LADs at the NL also interact with Lamin A, as demonstrated by the clear labeling of the nuclear rim by m6A-Tracer in the presence of Dam-Lamin A.
Lamin A and BAF Compete for Genome-NL Interactions
Because the m6A-Tracer data suggest that Lamin A can interact with internally positioned LADs, we reasoned that Lamin A could sequester these LADs in the nuclear interior and prevent them from interacting with the NL. To test this “tug of war” model, we reduced the levels of Lamin A by siRNA-mediated knockdowns, and then determined the contact frequencies of LADs with the NL (using Dam-Lamin B1) as previously described.Citation12 Strikingly, reducing the levels of Lamin A results on average in a ~1.4-fold increase in contact frequencies between several LADs and Lamin B1 ( and ). This is in agreement with a model in which Lamin A and B1 compete for the same LADs.
Figure 3. Lamin A and BAF compete with Lamin B1 for LAD binding. (A) m6A accumulation by Dam-LaminB1 at three LADs (Cyp2C19, LAD1, and LAD2) and one inter-LAD region (UBE2B) after siRNA knockdowns (KD) as indicated. Error bars indicate standard deviations (n = 4). *P < 0.05, **P < 0.01 according to a paired t test. (B) Estimated knockdown efficiencies, expressed as residual mRNA levels compared with control. Note the significant effect of Lamin A KD on Lamin B1 interactions with two LADs (A), despite the rather mild knockdown of Lamin A (mRNA level reduced to 67% of control).
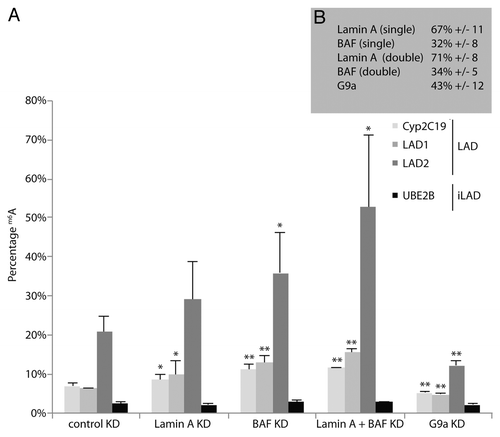
We also determined the effect of BAF on LAD-Lamin B1 interactions. Again, knockdown of BAF substantially increased the contact frequency of several LADs with Lamin B1 (by ~1.8-fold on average). Albeit modestly, a double knockdown of both Lamin A and BAF results in an even further enhancement of the contact-frequencies (~2.2-fold compared with the control knockdown). In contrast, reducing the levels of H3K9me2 by knocking down G9a results into a reduction of genome-NL interactions (), which is consistent with what we reported previously.Citation12 Hence, Lamin A/BAF and G9a are opposing forces in the regulation of genome-NL contact frequencies.
We do not know at which stage of the cell cycle Lamin A and BAF interfere with the positioning of LADs at the NL. This could happen right after mitosis, when the NL is reassembled onto chromosomes, or in early G1, when chromatin is still mobileCitation22 and LADs stochastically assume their positions in the nucleusCitation12 (). Some LADs could be localized to the nucleolar periphery by an interaction of BAF and Lamin A with nucleophosminCitation23 and nucleolin,Citation24 respectively.
Figure 4. Cartoon model of a possible role of Lamin A in modulating NL interactions. In wild-type cells, LADs may be anchored at the NL by interacting with Lamin A (orange) or B (blue), or be kept in the nuclear interior by interacting with Lamin A, at the surface of nucleoli (left panel). In the absence of Lamin A, anchoring in the nuclear interior is lost and LADs are more likely to interact with Lamin B at the NL (right panel, arrows indicate relocated LADs).
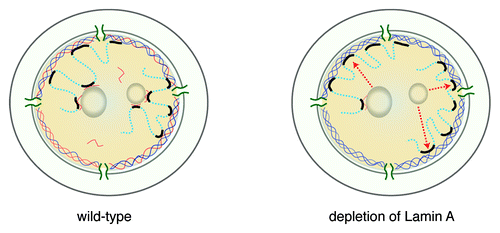
We note that our observation that Lamin A counteracts peripheral positioning of LADs seems contradictory to observations in mouse, where Lamin A and Lamin B Receptor (LBR) are thought to be redundantly involved in the anchoring of heterochromatin to the nuclear periphery.Citation25 Perhaps the modulatory role of Lamin A is different in the presence and absence of LBR, or is different between mouse and human cells.
Stochastic NL Interactions: Stochastic Gene Repression?
Most genes in LADs have very low expression levels, indicating that LADs constitute a repressive chromatin environment. In differentiating mouse ES cells, dissociation of genes from the NL often precedes the actual transcriptional activation at a next differentiation step.Citation3 Conversely, in Drosophila neuronal progenitor cells, tethering of the hunchback gene to the NL is necessary for its stable repression.Citation26 These observations support the notion that the NL is a repressive environment. Indeed, reporter genes integrated in LADs tend to be ~5-fold less active than the same reporter genes integrated in inter-LAD regions,Citation27 and tethering experiments have indicated that contact with NL itself can contribute to this transcriptional repression,Citation28-Citation30 although not in all instances.Citation31
Even though LADs exhibit detachment from the NL in a subset of cells, for the majority of genes in LADs no expression is detected by mRNA profiling of pools of cells.Citation12 Apparently, for these genes, the stochastic detachment from the NL does not lead to strong transcriptional activation. Possibly, association of Lamin A with these genes preserves their repressed status in the nucleoplasm. Indeed, binding of Lamin A to promoters is generally incompatible with transcription,Citation32,Citation33 and downregulation of Lamin A results in increased H3K4me3 levels at promoters, a mark that is thought to be permissive for transcription.Citation33 Other nucleoplasmic proteins such as BAF may further mark the detached LADs for repression.
It is also possible that transient NL contacts are sufficient to reinforce epigenetic silencing mechanisms for longer periods. For example, the histone deacetylase HDAC3 was found enriched at the NLCitation34-Citation36; perhaps an occasional visit to the NL is sufficient to remove histone acetylation from LADs and thereby preserve repression over multiple generations.
Nevertheless, some genes located in LADs are transcriptionally active according to mRNA profiling data of cell pools.Citation2,Citation12 We observed for several of these genes that their stochastic location at the NL is inversely linked to levels of H3K36me3, which is a marker of transcriptional activity.Citation12 Stochastic activity of genes has been extensively studied,Citation37,Citation38 but so far had not been linked to nuclear positioning to our knowledge. Thus, for some genes, the stochastic detachment from the NL coincides with transcriptional activation. This leads to intrinsic cell-to-cell variability in gene expression, a feature that may be exploited during cell-fate transitions.Citation39
Future Directions
Nuclear architecture is much more dynamic than anticipated. Whether all cells at different stages of development exhibit similarly stochastic nuclear organization of LADs should be further investigated. It is tempting to speculate that, when cells become gradually committed to a particular lineage, the stochastic positioning of LADs decreases concomitantly. It will be interesting to compare the dynamics of LAD positioning in various cell types.
At present it is not clear whether NL contacts are equally stochastic for all LADs, or whether some LADs interact more robustly with the NL than others. This may be studied by fluorescence in situ hybridization of individual LADs, by tracking of selected LADs using new tagging methods,Citation40 or perhaps in the future by the construction of genome-wide maps of NL interactions in single cells.
So far, we have identified three proteins that modulate the NL contact frequency of LADs: G9a, Lamin A, and BAF. It is likely that other proteins are involved in this regulatory process. Identification of these proteins can provide us with tools to further investigate the links of dynamic LAD-NL interactions with stochastic gene expression, and perhaps with the single-cell dynamics of other nuclear processes such as DNA replication and DNA repair.
Experimental Procedures
DamID
DamID of BAF and Lamin B2 was performed as described in reference Citation41 except that Dam-Lamin B2 was introduced by transfection of the pLgw-EcoDam-V5-Lamin B2 plasmid with lipofectamine (Clontech) 48 h prior to genomic DNA collection. DamID microarray data was normalized as described in reference Citation42.
Immunofluorescence labeling and microscopy
Immunofluorescence was performed as described in reference Citation12. The antibodies used in this study are from Abcam: Lamin B1 (ab16048) and NPM1 (ab10530). Images were acquired on a confocal laser scanning Leica TCS SP2.
qPCR quantification of m6A levels
Quantitative PCR measurements of m6A levels at selected GATC sequences were performed as described in reference Citation12; Dam-LaminB1 expression was induced three days post siRNA transfections for 20 h. RNAi efficiencies were determined by RT-qPCR of the respective mRNAs 72 h after siRNA transfection.
Cell line
The m6A-Tracer line was derived by transfection of HT1080 cells with TetO-puromycin-IRES-m6A-Tracer and subsequent clonal selection with 2 μg/ml puromycin (Sigma).
m6A-Tracer experiment
The m6A-Tracer line was grown in absence of doxycycline and transfected with pLgw-EcoDam-V5-Lamin B1, pLgw-EcoDam-V5-Lamin B2, or pLgw-EcoDam-V5-Lamin A. Twenty hours after transfections cells were fixed and prepared for Immunofluorescence. For , n = 40 for Lamin B1 and Lamin B2, and n = 40 and n = 33 for Lamin A. Cells were manually scored as having peri-nucleolar enrichment when a striking enrichment of the m6A-Tracer signal was apparent around nucleoli in respect to the overall signal throughout the nucleoplasm.
RNAi m6A-quantification assay and RT-qPCR
As performed in reference Citation12, Lamin A primer sequences for RT-qPCR: CCGAGTCTGAAGAGGTGGTC (forward), AACTCCTCAC GCACTTTGCT (reverse). BAF primer sequences for RT-qPCR: GAACCGTTAC GGGAACTGAA (forward), CCCAGGACTT CACCAATCC (reverse).
Data availability
Genome-wide DamID data are available from the Gene Expression Omnibus (http://www.ncbi.nlm.nih.gov/geo/), accession GSE55066.
Disclosure of Potential Conflicts of Interest
No potential conflict of interest was disclosed.
Acknowledgments
We thank the NKI Genomics Core Facility for microarray hybridizations, and members of our laboratory for helpful suggestions. This work was supported by an EMBO Long-term Fellowship and NWO-ALW VENI (J.K.), and by NWO-ALW VICI (B.v.S.).
References
- Pickersgill H, Kalverda B, de Wit E, Talhout W, Fornerod M, van Steensel B. Characterization of the Drosophila melanogaster genome at the nuclear lamina. Nat Genet 2006; 38:1005 - 14; http://dx.doi.org/10.1038/ng1852; PMID: 16878134
- Guelen L, Pagie L, Brasset E, Meuleman W, Faza MB, Talhout W, Eussen BH, de Klein A, Wessels L, de Laat W, et al. Domain organization of human chromosomes revealed by mapping of nuclear lamina interactions. Nature 2008; 453:948 - 51; http://dx.doi.org/10.1038/nature06947; PMID: 18463634
- Peric-Hupkes D, Meuleman W, Pagie L, Bruggeman SW, Solovei I, Brugman W, Gräf S, Flicek P, Kerkhoven RM, van Lohuizen M, et al. Molecular maps of the reorganization of genome-nuclear lamina interactions during differentiation. Mol Cell 2010; 38:603 - 13; http://dx.doi.org/10.1016/j.molcel.2010.03.016; PMID: 20513434
- Ikegami K, Egelhofer TA, Strome S, Lieb JD. Caenorhabditis elegans chromosome arms are anchored to the nuclear membrane via discontinuous association with LEM-2. Genome Biol 2010; 11:R120; http://dx.doi.org/10.1186/gb-2010-11-12-r120; PMID: 21176223
- Dechat T, Gajewski A, Korbei B, Gerlich D, Daigle N, Haraguchi T, Furukawa K, Ellenberg J, Foisner R. LAP2alpha and BAF transiently localize to telomeres and specific regions on chromatin during nuclear assembly. J Cell Sci 2004; 117:6117 - 28; http://dx.doi.org/10.1242/jcs.01529; PMID: 15546916
- Moir RD, Yoon M, Khuon S, Goldman RD. Nuclear lamins A and B1: different pathways of assembly during nuclear envelope formation in living cells. J Cell Biol 2000; 151:1155 - 68; http://dx.doi.org/10.1083/jcb.151.6.1155; PMID: 11121432
- Zheng R, Ghirlando R, Lee MS, Mizuuchi K, Krause M, Craigie R. Barrier-to-autointegration factor (BAF) bridges DNA in a discrete, higher-order nucleoprotein complex. Proc Natl Acad Sci U S A 2000; 97:8997 - 9002; http://dx.doi.org/10.1073/pnas.150240197; PMID: 10908652
- Bradley CM, Ronning DR, Ghirlando R, Craigie R, Dyda F. Structural basis for DNA bridging by barrier-to-autointegration factor. Nat Struct Mol Biol 2005; 12:935 - 6; http://dx.doi.org/10.1038/nsmb989; PMID: 16155580
- Montes de Oca R, Andreassen PR, Wilson KL. Barrier-to-Autointegration Factor influences specific histone modifications. Nucleus 2011; 2:580 - 90; http://dx.doi.org/10.4161/nucl.2.6.17960; PMID: 22127260
- Montes de Oca R, Lee KK, Wilson KL. Binding of barrier to autointegration factor (BAF) to histone H3 and selected linker histones including H1.1. J Biol Chem 2005; 280:42252 - 62; http://dx.doi.org/10.1074/jbc.M509917200; PMID: 16203725
- Shimi T, Koujin T, Segura-Totten M, Wilson KL, Haraguchi T, Hiraoka Y. Dynamic interaction between BAF and emerin revealed by FRAP, FLIP, and FRET analyses in living HeLa cells. J Struct Biol 2004; 147:31 - 41; http://dx.doi.org/10.1016/j.jsb.2003.11.013; PMID: 15109603
- Kind J, Pagie L, Ortabozkoyun H, Boyle S, de Vries SS, Janssen H, Amendola M, Nolen LD, Bickmore WA, van Steensel B. Single-cell dynamics of genome-nuclear lamina interactions. Cell 2013; 153:178 - 92; http://dx.doi.org/10.1016/j.cell.2013.02.028; PMID: 23523135
- Gibcus JH, Dekker J. The hierarchy of the 3D genome. Mol Cell 2013; 49:773 - 82; http://dx.doi.org/10.1016/j.molcel.2013.02.011; PMID: 23473598
- Nagano T, Lubling Y, Stevens TJ, Schoenfelder S, Yaffe E, Dean W, Laue ED, Tanay A, Fraser P. Single-cell Hi-C reveals cell-to-cell variability in chromosome structure. Nature 2013; 502:59 - 64; http://dx.doi.org/10.1038/nature12593; PMID: 24067610
- van Koningsbruggen S, Gierlinski M, Schofield P, Martin D, Barton GJ, Ariyurek Y, den Dunnen JT, Lamond AI. High-resolution whole-genome sequencing reveals that specific chromatin domains from most human chromosomes associate with nucleoli. Mol Biol Cell 2010; 21:3735 - 48; http://dx.doi.org/10.1091/mbc.E10-06-0508; PMID: 20826608
- Németh A, Conesa A, Santoyo-Lopez J, Medina I, Montaner D, Péterfia B, Solovei I, Cremer T, Dopazo J, Längst G. Initial genomics of the human nucleolus. PLoS Genet 2010; 6:e1000889; http://dx.doi.org/10.1371/journal.pgen.1000889; PMID: 20361057
- Wen B, Wu H, Shinkai Y, Irizarry RA, Feinberg AP. Large histone H3 lysine 9 dimethylated chromatin blocks distinguish differentiated from embryonic stem cells. Nat Genet 2009; 41:246 - 50; http://dx.doi.org/10.1038/ng.297; PMID: 19151716
- Bian Q, Khanna N, Alvikas J, Belmont AS. β-Globin cis-elements determine differential nuclear targeting through epigenetic modifications. J Cell Biol 2013; 203:767 - 83; http://dx.doi.org/10.1083/jcb.201305027; PMID: 24297746
- Towbin BD, González-Aguilera C, Sack R, Gaidatzis D, Kalck V, Meister P, Askjaer P, Gasser SM. Step-wise methylation of histone H3K9 positions heterochromatin at the nuclear periphery. Cell 2012; 150:934 - 47; http://dx.doi.org/10.1016/j.cell.2012.06.051; PMID: 22939621
- Shimi T, Pfleghaar K, Kojima S, Pack CG, Solovei I, Goldman AE, Adam SA, Shumaker DK, Kinjo M, Cremer T, et al. The A- and B-type nuclear lamin networks: microdomains involved in chromatin organization and transcription. Genes Dev 2008; 22:3409 - 21; http://dx.doi.org/10.1101/gad.1735208; PMID: 19141474
- Meuleman W, Peric-Hupkes D, Kind J, Beaudry JB, Pagie L, Kellis M, Reinders M, Wessels L, van Steensel B. Constitutive nuclear lamina-genome interactions are highly conserved and associated with A/T-rich sequence. Genome Res 2013; 23:270 - 80; http://dx.doi.org/10.1101/gr.141028.112; PMID: 23124521
- Thomson I, Gilchrist S, Bickmore WA, Chubb JR. The radial positioning of chromatin is not inherited through mitosis but is established de novo in early G1. Curr Biol 2004; 14:166 - 72; http://dx.doi.org/10.1016/j.cub.2003.12.024; PMID: 14738741
- Montes de Oca R, Shoemaker CJ, Gucek M, Cole RN, Wilson KL. Barrier-to-autointegration factor proteome reveals chromatin-regulatory partners. PLoS One 2009; 4:e7050; http://dx.doi.org/10.1371/journal.pone.0007050; PMID: 19759913
- Roux KJ, Kim DI, Raida M, Burke B. A promiscuous biotin ligase fusion protein identifies proximal and interacting proteins in mammalian cells. J Cell Biol 2012; 196:801 - 10; http://dx.doi.org/10.1083/jcb.201112098; PMID: 22412018
- Solovei I, Wang AS, Thanisch K, Schmidt CS, Krebs S, Zwerger M, Cohen TV, Devys D, Foisner R, Peichl L, et al. LBR and lamin A/C sequentially tether peripheral heterochromatin and inversely regulate differentiation. Cell 2013; 152:584 - 98; http://dx.doi.org/10.1016/j.cell.2013.01.009; PMID: 23374351
- Kohwi M, Lupton JR, Lai SL, Miller MR, Doe CQ. Developmentally regulated subnuclear genome reorganization restricts neural progenitor competence in Drosophila. Cell 2013; 152:97 - 108; http://dx.doi.org/10.1016/j.cell.2012.11.049; PMID: 23332748
- Akhtar W, de Jong J, Pindyurin AV, Pagie L, Meuleman W, de Ridder J, Berns A, Wessels LF, van Lohuizen M, van Steensel B. Chromatin position effects assayed by thousands of reporters integrated in parallel. Cell 2013; 154:914 - 27; http://dx.doi.org/10.1016/j.cell.2013.07.018; PMID: 23953119
- Reddy KL, Zullo JM, Bertolino E, Singh H. Transcriptional repression mediated by repositioning of genes to the nuclear lamina. Nature 2008; 452:243 - 7; http://dx.doi.org/10.1038/nature06727; PMID: 18272965
- Finlan LE, Sproul D, Thomson I, Boyle S, Kerr E, Perry P, Ylstra B, Chubb JR, Bickmore WA. Recruitment to the nuclear periphery can alter expression of genes in human cells. PLoS Genet 2008; 4:e1000039; http://dx.doi.org/10.1371/journal.pgen.1000039; PMID: 18369458
- Dialynas G, Speese S, Budnik V, Geyer PK, Wallrath LL. The role of Drosophila Lamin C in muscle function and gene expression. Development 2010; 137:3067 - 77; http://dx.doi.org/10.1242/dev.048231; PMID: 20702563
- Kumaran RI, Spector DL. A genetic locus targeted to the nuclear periphery in living cells maintains its transcriptional competence. J Cell Biol 2008; 180:51 - 65; http://dx.doi.org/10.1083/jcb.200706060; PMID: 18195101
- Lee DC, Welton KL, Smith ED, Kennedy BK. A-type nuclear lamins act as transcriptional repressors when targeted to promoters. Exp Cell Res 2009; 315:996 - 1007; http://dx.doi.org/10.1016/j.yexcr.2009.01.003; PMID: 19272320
- Lund E, Oldenburg AR, Delbarre E, Freberg CT, Duband-Goulet I, Eskeland R, Buendia B, Collas P. Lamin A/C-promoter interactions specify chromatin state-dependent transcription outcomes. Genome Res 2013; 23:1580 - 9; http://dx.doi.org/10.1101/gr.159400.113; PMID: 23861385
- Somech R, Shaklai S, Geller O, Amariglio N, Simon AJ, Rechavi G, Gal-Yam EN. The nuclear-envelope protein and transcriptional repressor LAP2beta interacts with HDAC3 at the nuclear periphery, and induces histone H4 deacetylation. J Cell Sci 2005; 118:4017 - 25; http://dx.doi.org/10.1242/jcs.02521; PMID: 16129885
- Demmerle J, Koch AJ, Holaska JM. The nuclear envelope protein emerin binds directly to histone deacetylase 3 (HDAC3) and activates HDAC3 activity. J Biol Chem 2012; 287:22080 - 8; http://dx.doi.org/10.1074/jbc.M111.325308; PMID: 22570481
- Zullo JM, Demarco IA, Piqué-Regi R, Gaffney DJ, Epstein CB, Spooner CJ, Luperchio TR, Bernstein BE, Pritchard JK, Reddy KL, et al. DNA sequence-dependent compartmentalization and silencing of chromatin at the nuclear lamina. Cell 2012; 149:1474 - 87; http://dx.doi.org/10.1016/j.cell.2012.04.035; PMID: 22726435
- Lionnet T, Singer RH. Transcription goes digital. EMBO Rep 2012; 13:313 - 21; http://dx.doi.org/10.1038/embor.2012.31; PMID: 22410830
- Raj A, van Oudenaarden A. Single-molecule approaches to stochastic gene expression. Annu Rev Biophys 2009; 38:255 - 70; http://dx.doi.org/10.1146/annurev.biophys.37.032807.125928; PMID: 19416069
- Balázsi G, van Oudenaarden A, Collins JJ. Cellular decision making and biological noise: from microbes to mammals. Cell 2011; 144:910 - 25; http://dx.doi.org/10.1016/j.cell.2011.01.030; PMID: 21414483
- Chen B, Gilbert LA, Cimini BA, Schnitzbauer J, Zhang W, Li GW, Park J, Blackburn EH, Weissman JS, Qi LS, et al. Dynamic imaging of genomic loci in living human cells by an optimized CRISPR/Cas system. Cell 2013; 155:1479 - 91; http://dx.doi.org/10.1016/j.cell.2013.12.001; PMID: 24360272
- Vogel MJ, Peric-Hupkes D, van Steensel B. Detection of in vivo protein-DNA interactions using DamID in mammalian cells. Nat Protoc 2007; 2:1467 - 78; http://dx.doi.org/10.1038/nprot.2007.148; PMID: 17545983
- Peric-Hupkes D, Meuleman W, Pagie L, Bruggeman SW, Solovei I, Brugman W, Gräf S, Flicek P, Kerkhoven RM, van Lohuizen M, et al. Molecular maps of the reorganization of genome-nuclear lamina interactions during differentiation. Mol Cell 2010; 38:603 - 13; http://dx.doi.org/10.1016/j.molcel.2010.03.016; PMID: 20513434