Abstract
Cancers utilize multiple mechanisms to overcome immune responses. Emerging evidence suggest that immunotherapy of cancer should focus on inducing and re-programming cells of the innate and adaptive immune systems rather than focusing solely on T cells. Recently, we have shown that such a multifaceted approach can improve immunotherapy of breast cancer.
Major barriers/challenges to the advancement of cancer immunotherapy include: (1) immunological tolerance due to the fact that cancer cells originate from normal tissue to which cells of the adaptive immune system were tolerized; (2) tumor escape as a result of epigenetic changes in the tumor cells induced by immune responses, e.g., antigen loss, MHC class I loss or “missing” unknown, yet critical, target antigens; (3) tumor-induced immune suppression mediated by an increased population of myeloid-derived suppressor cells (MDSC) in cancer patients.Citation1-Citation4 In fact, cancer cells utilize multiple strategies to survive in such an immunologically hostile environment, however many strategies used in cancer immunotherapy have been narrowly focused on a specific type of immune cell, particularly CD8+ T cells. Innate immune cells such as NK cells or NKT cells are usually considered as a secondary source of immunotherapy when tumor cells escape from adaptive immune responses by losing their target antigen or MHC class I molecule. In addition, patients who participate in immunotherapy clinical trials have received chemotherapy and radiation therapy. Such conventional therapies affect their immune system. Therefore, an effective cancer immunotherapy is expected to overcome the above mentioned barriers, and to be designed based on an understanding of the role of conventional therapies in boosting or compromising the immune responses.
Despite recent advances in adoptive cellular therapy (ACT) of melanoma, no success has been achieved in ACT of breast cancer. This is in part due to low immunogenicity of breast cancer expressing self antigens compared with melanoma that expresses a variety of highly immunogenic antigens as suitable targets for immunotherapy,Citation5 as well as increased MDSC in breast cancer patients.Citation3,Citation4 Using autologous T cells that are weakly and inefficiently reactive against the self antigens expressed by breast tumors would not generate objective responses. Therefore, re-programming of tumor-reactive immune cells toward the most effective phenotypes may be the only way to cure breast cancer and/or prevent recurrences immunologically. In addition, most ACT protocols have focused on T cells and ignored a critical role of the innate immune cells including NKT cells and NK cells in anti-tumor protection. Given the critical cross talk between cells of the innate and adaptive immune systems, a combined approach utilizing NKT cells, NK cells and T cells could result in highly effective anti-tumor immune responses.Citation6,Citation7 Our recent findings suggest that such a multifaceted strategy can overcome MDSC mediated immune suppression as well as tumor escape in the FVBN202 mouse model of spontaneous mammary carcinoma.Citation8 Others have demonstrated that NKT cells play a key role in overcoming MDSC by converting them into antigen-presenting cells (APC), thereby rescuing T cells from suppression and improving their effector function.Citation9,Citation10 It has been reported that T central memory (TCM) phenotypes are more effective than T effector (TE) phenotypes in generating long-lasting protection against tumor cells.Citation11,Citation12 The presence of NKT memory cells has also been suggested to be protective against tumor cells.Citation13 Therefore, the most effective ACT strategy would be to re-program cells of both innate and adaptive immune systems and differentiate the T cells toward memory phenotypes, while at the same time overcoming tumor-induced immune suppression.
We have recently developed an antigen-free protocol by means of pharmacological agents, bryostatin 1 (B) and ionomycin (I), and combined common gamma chain (γ-c) cytokines for re-programming tumor-reactive cells of the innate (NKT cells and NK cells) and adaptive (CD4+ and CD8+ T cells) immune systems which displayed resistance to MDSC. Broystatin 1 is a naturally occurring antineoplastic drug which is also a potent modulator of protein kinase C (PKC). Short-term effects of bryostatin 1 include activation of classical and novel PKCs, whereas prolonged stimulation leads to lowered PKC activation. It was reported long ago and repeatedly that bryostatin + ionomycin (B/I) selectively stimulate tumor-sensitized T cells in vitro; when lymphocytes from sarcoma-bearing mice were activated with B/I and expanded in IL-2, tumor-specific T cell frequency increased by orders of magnitude compared with the starting population.Citation14 In fact, B/I selectively activated CD62Llow (sensitized) T cells from mice bearing 4T1 mammary carcinomas.Citation15 Emerging interests in bryostatin 1 as an immune modulator have resulted in a better understanding of its role in the modulation of antigen presentation. For instance, it was reported that bryostatin 1 acts as TLR-4 ligand and activates dendritic cells (DCs),Citation16 and upregulates expression of IFNγ receptor in monocytesCitation17 as well as induction of IFNγ and T-bet transcripts.Citation18 These data suggest that B/I could be a potent modulator of the immune cells.
The common γ-c cytokines IL-2, IL-7 and IL-15 play a key role in homeostasis of the immune cells. An understanding of the distinct properties of these cytokines will lead to the development of an effective formulation or combination for the expansion of tumor-reactive immune cells ex vivo during a pharmacological re-programming. In addition to being a T cell growth factor, IL-2 also supports differentiation of CD8+ T cells toward effector and effector/memory phenotypes (TE and TEM) by downregulation of lymph node homing receptors CD62L and CCR7. This results in the trafficking of the IL-2-expanded T cells to the tumor site. Therefore, such cells may induce early anti-tumor responses but may not lead to a long-term memory response. IL-7 is crucial for the survival and homeostatic expansion of naive and memory CD8+ T cells; it is secreted by stromal cells, epithelial cells and fibroblasts but is not produced by lymphocytes. The IL-7 R is expressed by T cells, pre-B cells and DCs. The receptor comprises two polypeptides, an affinity binding receptor IL-7 Rα or CD127 and a signaling γ-chain receptor CD132. Because of an important role of IL-7 in all stages of T cell development and maintenance, it has been used in clinical trials in an attempt to increase the replenishment of T cells. Injection of IL-7 resulted in the expansion of both CD4+ and CD8+ T cells as well as a relative reduction of CD4+ Tregs.Citation19 IL-15 is produced by monocytes, DCs and epithelial cells. IL-15 R is expressed by T cells and NK cells, and consists of three polypeptide subunits: an IL-15 Rα chain, which determines its binding to IL-15, and shared IL-2 β (CD122) and γ (CD132) chains.
About 20% of the human CD8+ T cell pool in peripheral blood has low expression of CD127. Although CD127 is a specific receptor for IL-7, its expression on T cells also determines responsiveness of T cells to common gamma chain cytokines other than IL-7. For example, IL-15 decreases activation induced cell death (AICD) in CD8+CD127+ T cells but not in CD8+CD127− T cells while inducing comparable proliferation of the two subsets.Citation20 Such a differential effect was in part mediated by IL-15-induced expression of anti-apoptotic Bcl-2 as well as inhibition of the pro-apoptotic Bim in CD8+CD127+ T cells but not in CD8+CD127− T cells. Although IL-15 can induce expression of Bcl-2 in naive and memory T cells, its best defined role is supporting memory T cells. It has been shown that naïve T cells and TCM cells are CD8+CD127+ while TE and TEM cells are mostly CD8+CD127−.Citation21 Culture of antigen-experienced T cells with IL-15 ex vivo restores their ability to respond to the antigen.Citation21
Due to the variable expression of the common γ-c receptors at different stages of immune cell homeostasis, a sequential use of the cytokines should be considered during the expansion and re-programming of tumor-sensitized immune cells.
The role of NKT cells in tumor immunity has not been studied extensively. NKT cells are classified into three types which include: (1) type I or classical or invariant NKT cells (iNKT cells); (2) type II or non-classical or non-invariant NKT cells. Type I and II NKT cells recognize glycolipid antigens associated with MHC class I-like molecules CD1d; (3) NKT-like cells or CD1d-independent NKT cells.Citation22 Type I NKT cells express an invariant Vα24Jα18 chain paired with a Vβ11 in humans or an invariant Vα14Jα18 chain paired with a Vβ8.2, Vβ2, or Vβ7 in mice. In contrast, TcR Vβ regions used by type II NKT cells are highly diverse. Type I NKT cells produce IFNγ whereas type II NKT cells produce IL-13 that facilitates production of TGFβ by myeloid cells.Citation23 Recent studies have shown that iNKT cells conferred protection against lymphoma, whereas type II NKT cells facilitated immune suppression.Citation24 For instance, increased iNKT cells at the tumor site of patients with colorectal cancer were found to be associated with a favorable prognosis.Citation25 iNKT cells have been shown to act as adjuvants for anti-tumor T cell vaccines.Citation26,Citation27 The precise mechanism responsible for such an adjuvant effect is not fully understood. A recent report suggested that iNKT cells were shown to increase expression of CD70 on DCs, following immunization with the glycolipid, α-galactosylceramide (α-GalCer), in mice, thereby supporting CD8+ T cell responses through engagement with the CD27 receptor.Citation7 Most iNKT cells in mice are double negative (CD4−CD8−) and express CD28 as well as CD154 upon activation.Citation6,Citation7 Ligation of these costimulatory molecules increases secretion of IFNγ by iNKT cells.
It was reported that MDSC loaded with α-GalCer on their CD1d showed enhanced immunostimulatory function through interaction with activated iNKT cells. Activated iNKT cells in turn converted MDSC into antigen-presenting cells (APCs), and supported antigen-specific proliferation of IFNγ producing CD8+ T cells.Citation9 In humans, activated iNKT cells have also been shown to direct monocytes to differentiate into immature DCs through the engagement of CD1d on monocytes.Citation28 These reports underscore a critical role of the activated iNKT cells in interactions with MDSC and tumor-reactive T cells. Very recently we made a similar observation in the FVBN202 transgenic mouse model of breast carcinoma, where anti-tumor efficacy of HER-2/neu-specific T cells, in vitro and in vivo, was influenced by the presence or absence of activated NKT cells.Citation8
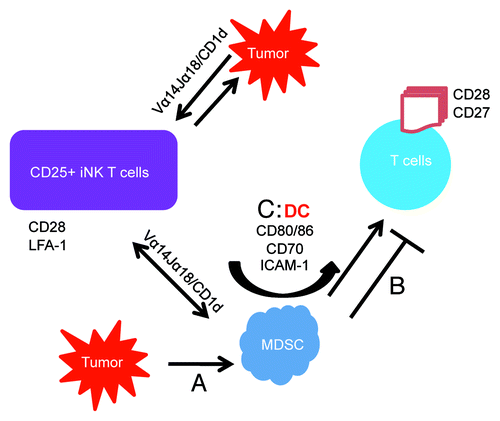
Based on these findings we propose a model to explain a cross talk between NKT cells and MDSC as well as with T cells during an effective anti-tumor immune response (). According to this model, double negative (CD4−CD8−) CD25+ invariant NKT (iNKT) cells interact with CD1d on MDSC, resulting in the conversion of MDSC into DC by increasing the expression of CD80/86, CD70 and ICAM-1. Engagement of CD80 and CD70 on newly converted DCs with CD28 and CD27 on T cells support T cell responses to the tumor cells and overcome MDSC suppression. CD25+ iNKT cells can also respond to tumor cells and MDSC in a CD1d-dependent manner. Extensive production of tumor-specific IFNγ by the expanded cells may also overcome tumor relapse that we found to be due to low levels of IFNγ production. This hypothetical model has been supported by our recent publicationCitation8 showing that tumor-specific IFNγ production was significantly increased by NKT cells when MDSC were present. In addition, the presence of NKT cells was required in order to overcome MDSC-mediated T cell suppression.
Figure 1. Mechanisms by which CD25+ iNKT cells interact with MDSC and rescue T cells from suppression. Tumor-derived soluble factors increase MDSC (A) which in turn suppress anti-tumor T cell responses (B). Activated CD25+ NKT cells interact with CD1d on tumor cells and MDSC and demonstrate enhanced anti-tumor responses (C). This will result in MDSCs increasing expression of CD80/86, CD70, ICAM-1 thus effectively converting to a DC phenotype, which then interacts with CD28 and CD27 on activated T cells, thereby enhancing T cell anti-tumor responses.
The source of tumor-reactive immune cells that are used for ex vivo re-programming is critical. We showed that radiation therapy compromised phenotypic distribution of T cells such that re-programming of these cells did not yield TCM phenotypes and failed to protect animals against the tumor cells.Citation8 Altogether, recent evidence demonstrates a shift from adaptive T cell responses to a multifaceted cellular immunity by means of immune cell expansion using γ-c cytokines utilizing cells harvested prior to radiation therapy for the treatment of human carcinomas.
| Abbreviations: | ||
| ACT | = | adoptive cellular therapy |
| MDSC | = | myeloid-derived suppressor cells |
| NKT cells | = | natural killer T cells |
| B/I | = | bryostatin 1/ionomycin |
| PKC | = | protein kinase C |
| ICAM | = | intracellular adhesion molecule |
Acknowledgments
This work was supported by the VCU Presidential Research Incentive Fund (M.H. Manjili). We gratefully acknowledge the support of VCU Massey Cancer Center and the Commonwealth Foundation for Cancer Research.
References
- Gabitass RF, Annels NE, Stocken DD, Pandha HA, Middleton GW. Elevated myeloid-derived suppressor cells in pancreatic, esophageal and gastric cancer are an independent prognostic factor and are associated with significant elevation of the Th2 cytokine interleukin-13. Cancer Immunol Immunother 2011; In press http://dx.doi.org/10.1007/s00262-011-1028-0; PMID: 21644036
- Poschke I, Mougiakakos D, Hansson J, Masucci GV, Kiessling R. Immature immunosuppressive CD14+HLA-DR-/low cells in melanoma patients are Stat3hi and overexpress CD80, CD83, and DC-sign. Cancer Res 2010; 70:4335 - 45; http://dx.doi.org/10.1158/0008-5472.CAN-09-3767; PMID: 20484028
- Diaz-Montero CM, Salem ML, Nishimura MI, Garrett-Mayer E, Cole DJ, Montero AJ. Increased circulating myeloid-derived suppressor cells correlate with clinical cancer stage, metastatic tumor burden, and doxorubicin-cyclophosphamide chemotherapy. Cancer Immunol Immunother 2009; 58:49 - 59; http://dx.doi.org/10.1007/s00262-008-0523-4; PMID: 18446337
- Mundy-Bosse BL, Thornton LM, Yang HC, Andersen BL, Carson WE. Psychological stress is associated with altered levels of myeloid-derived suppressor cells in breast cancer patients. Cell Immunol 2011; 270:80 - 7; http://dx.doi.org/10.1016/j.cellimm.2011.04.003; PMID: 21600570
- Davis ID, Chen W, Jackson H, Parente P, Shackleton M, Hopkins W, et al. Recombinant NY-ESO-1 protein with ISCOMATRIX adjuvant induces broad integrated antibody and CD4(+) and CD8(+) T cell responses in humans. Proc Natl Acad Sci USA 2004; 101:10697 702; http://dx.doi.org/10.1073/pnas.0403572101; PMID: 15252201
- Hayakawa Y, Takeda K, Yagita H, Van Kaer L, Saiki I, Okumura K. Differential regulation of Th1 and Th2 functions of NKT cells by CD28 and CD40 costimulatory pathways. J Immunol 2001; 166:6012 - 8; PMID: 11342617
- Taraban VY, Martin S, Attfield KE, Glennie MJ, Elliott T, Elewaut D, et al. Invariant NKT cells promote CD8+ cytotoxic T cell responses by inducing CD70 expression on dendritic cells. J Immunol 2008; 180:4615 - 20
- Kmieciak M, Basu D, Payne KK, Toor A, Yacoub A, Wang XY, et al. Activated NKT cells and NK cells render T cells resistant to myeloid-derived suppressor cells and result in an effective adoptive cellular therapy against breast cancer in the FVBN202 transgenic mouse. J Immunol 2011; 187:708 - 17; http://dx.doi.org/10.4049/jimmunol.1100502; PMID: 21670315
- Ko HJ, Lee JM, Kim YJ, Kim YS, Lee KA, Kang CY. Immunosuppressive myeloid-derived suppressor cells can be converted into immunogenic APCs with the help of activated NKT cells: an alternative cell-based antitumor vaccine. J Immunol 2009; 182:1818 - 28; http://dx.doi.org/10.4049/jimmunol.0802430; PMID: 19201833
- Lee JM, Seo JH, Kim YJ, Kim YS, Ko HJ, Kang CY. The restoration of myeloid-derived suppressor cells as functional antigen-presenting cells by NKT cell help and all-trans-retinoic acid treatment. Int J Cancer 2011; In press http://dx.doi.org/10.1002/ijc.26411; PMID: 21898392
- Klebanoff CA, Gattinoni L, Torabi-Parizi P, Kerstann K, Cardones AR, Finkelstein SE, et al. Central memory self/tumor-reactive CD8+ T cells confer superior antitumor immunity compared with effector memory T cells. Proc Natl Acad Sci USA 2005; 102:9571 - 6; http://dx.doi.org/10.1073/pnas.0503726102; PMID: 15980149
- Perret R, Ronchese F. Memory T cells in cancer immunotherapy: which CD8 T-cell population provides the best protection against tumours?. Tissue Antigens 2008; 72:187 - 94; http://dx.doi.org/10.1111/j.1399-0039.2008.01088.x; PMID: 18627571
- Berzofsky JA, Terabe M. NKT cells in tumor immunity: opposing subsets define a new immunoregulatory axis. J Immunol 2008; 180:3627 - 35; PMID: 18322166
- Fleming MD, Bear HD, Lipshy K, Kostuchenko PJ, Portocarero D, McFadden AW, et al. Adoptive transfer of bryostatin-activated tumor-sensitized lymphocytes prevents or destroys tumor metastases without expansion in vitro. J Immunother Emphasis Tumor Immunol 1995; 18:147 - 55; PMID: 8770770
- Chin CS, Miller CH, Graham L, Parviz M, Zacur S, Patel B, et al. Bryostatin 1/ionomycin (B/I) ex vivo stimulation preferentially activates L-selectinlow tumor-sensitized lymphocytes. Int Immunol 2004; 16:1283 - 94; http://dx.doi.org/10.1093/intimm/dxh130; PMID: 15262898
- Ariza ME, Ramakrishnan R, Singh NP, Chauhan A, Nagarkatti PS, Nagarkatti M. Bryostatin-1, a naturally occurring antineoplastic agent, acts as a Toll-like receptor 4 (TLR-4) ligand and induces unique cytokines and chemokines in dendritic cells. J Biol Chem 2011; 286:24 - 34; http://dx.doi.org/10.1074/jbc.M110.135921; PMID: 21036898
- Garcia CS, Curiel RE, Mwatibo JM, Pestka S, Li H, Espinoza-Delgado I. The antineoplastic agent bryostatin-1 differentially regulates IFN-gamma receptor subunits in monocytic cells: transcriptional and posttranscriptional control of IFN-gamma R2. J Immunol 2006; 177:2707 - 16; PMID: 16888033
- Li H, Wojciechowski W, Dell'Agnola C, Lopez NE, Espinoza-Delgado I. IFN-gamma and T-bet expression in human dendritic cells from normal donors and cancer patients is controlled through mechanisms involving ERK-1/2-dependent and IL-12-independent pathways. J Immunol 2006; 177:3554 - 63; PMID: 16951314
- Rosenberg SA, Sportès C, Ahmadzadeh M, Fry TJ, Ngo LT, Schwarz SL, et al. IL-7 administration to humans leads to expansion of CD8+ and CD4+ cells but a relative decrease of CD4+ T-regulatory cells. J Immunother 2006; 29:313 - 9; http://dx.doi.org/10.1097/01.cji.0000210386.55951.c2; PMID: 16699374
- Crawley AM, Katz T, Parato K, Angel JB. IL-2 receptor gamma chain cytokines differentially regulate human CD8+CD127+ and CD8+CD127- T cell division and susceptibility to apoptosis. Int Immunol 2009; 21:29 - 42; http://dx.doi.org/10.1093/intimm/dxn120; PMID: 19011158
- Paiardini M, Cervasi B, Albrecht H, Muthukumar A, Dunham R, Gordon S, et al. Loss of CD127 expression defines an expansion of effector CD8+ T cells in HIV-infected individuals. J Immunol 2005; 174:2900 - 9; PMID: 15728501
- Berzofsky JA, Terabe M. A novel immunoregulatory axis of NKT cell subsets regulating tumor immunity. Cancer Immunol Immunother 2008; 57:1679 - 83; http://dx.doi.org/10.1007/s00262-008-0495-4; PMID: 18369622
- Godfrey DI, MacDonald HR, Kronenberg M, Smyth MJ, Van Kaer L. NKT cells: what's in a name?. Nat Rev Immunol 2004; 4:231 - 7; http://dx.doi.org/10.1038/nri1309; PMID: 15039760
- Renukaradhya GJ, Khan MA, Vieira M, Du W, Gervay-Hague J, Brutkiewicz RR. Type I NKT cells protect (and type II NKT cells suppress) the host's innate antitumor immune response to a B-cell lymphoma. Blood 2008; 111:5637 - 45; http://dx.doi.org/10.1182/blood-2007-05-092866; PMID: 18417738
- Tachibana T, Onodera H, Tsuruyama T, Mori A, Nagayama S, Hiai H, et al. Increased intratumor Valpha24-positive natural killer T cells: a prognostic factor for primary colorectal carcinomas. Clin Cancer Res 2005; 11:7322 - 7; http://dx.doi.org/10.1158/1078-0432.CCR-05-0877; PMID: 16243803
- Silk JD, Hermans IF, Gileadi U, Chong TW, Shepherd D, Salio M, et al. Utilizing the adjuvant properties of CD1d-dependent NK T cells in T cell-mediated immunotherapy. J Clin Invest 2004; 114:1800 - 11; PMID: 15599405
- Liu K, Idoyaga J, Charalambous A, Fujii S, Bonito A, Mordoh J, et al. Innate NKT lymphocytes confer superior adaptive immunity via tumor-capturing dendritic cells. J Exp Med 2005; 202:1507 - 16; http://dx.doi.org/10.1084/jem.20050956; PMID: 16330814
- Morales JK, Kmieciak M, Knutson KL, Bear HD, Manjili MH. GM-CSF is one of the main breast tumor-derived soluble factors involved in the differentiation of CD11b-Gr1- bone marrow progenitor cells into myeloid-derived suppressor cells. Breast Cancer Res Treat 2010; 123:39 - 49; http://dx.doi.org/10.1007/s10549-009-0622-8; PMID: 19898981