
Abstract
The discovery by Hans Spemann of the “organizer” tissue and its ability to induce the formation of the amphibian embryo’s neural tube inspired leading embryologists to attempt to elucidate embryonic induction’s underlying mechanisms. Since then several studies have described several developmental model system to better understand the role of specific signaling molecules, the interplay of different signals and tissue interactions in regulating tissue induction and patterning events. Different groups of workers set out to subject embryonic amphibian tissues and inductive adult organs to various extraction methods in the hope that the active agents could be isolated and chemically identified. In addition, a large number of well characterized chemical compounds were tested.
Keywords: :
The Nature of the Organizer
Embryogenesis is composed of two major morphogenetic processes: the biochemical patterning of the embryo and the mechanical morphogenetic movements that geometrically shape the embryo. The embryonic organization, i.e., the allocation or differentiation processes to various areas of the embryo, is laid down in its general feature by the polarity of the egg cell or by an interplay of factors derived from the egg polarity, sometime after fertilization. However, realization of the plan of organization is dependent upon multitude of epigenetic events occurring in the later stages.
The action of the “organizer” is the most important of these events. A series of well-designed surgical experiments demonstrated that the function of the organizer is to initiate directly or indirectly many of the differentiation processes in other areas of the embryo (induction), and at the sometime, to initiate in its own area a number of differentiation processes leading to the formation of the essential part of the axial system of the embryo. The effect of the organizer is to induce this area to switch from one type of differentiation to another, but not to acquire the ability to differentiate.
Embryonic induction consists of an interaction between inducing and responding tissues that brings about alterations in the developmental pathway of the responding tissue. Inductive stimuli operate only at certain stages, as a rule, during early development, and they are normally ineffective unless there is an intimate contact between inducing and reacting tissues. Once stimulated, the cells proceed along their new course of differentiation independently of a continued application of inducing stimulus.
Spemann’s and Mangold’s Experiment
As a young biologist Hans Spemann began work in 1894 at Würzburg as a doctoral student and teacher, and was the favorite pupil of Theodor Boveri. It was there, just after taking his doctorate, that he married Clara Binder. After 14 y at Würzburg, Spemann became professor at Rostock (1908–1914). He spent the years of World War I as director of the Kaiser Wilhelm Institute of Biology in Berlin-Dahlem; and in 1919 he succeeded Weismann as professor at Freiburg im Breisgau. He remained at Freiburg for the rest of his life, retiring in 1938.
In 1924, Spemann and his assistant Hilde Pröscholdt Mangold () published the results of their work in “W. Roux Archive für Entwicklungsmekanik der Organismen,” at the time the most prestigious journal in the field of experimental embryology ().Citation1 They showed that a partial second embryo could be induced by grafting one small region of a newt embryo onto a new site on another embryo and a supernumerary neural plate developed at the site of the graft. Only a narrow central part of this was of the donor origin. This small region they called the “organizer,” since it seemed to be responsible for the organization of a complete embryonic body.
Figure 1. A portrait of Hans Spemann and his assistant Hilde Pröscholdt Mangold.

Figure 2. The cover of the paper published in 1924 by Spemann and Mangold.

The grafted tissue was taken from the dorsal lip of the blastopore (the slit-like invagination that forms where gastrulation begins on the dorsal surface of the amphibian embryo) from a color less (unpigmented) Triton Cristatus and implanted it onto a deeply pigmented Triton Taeniatus. In the secondary embryo that resulted, most of the ectoderm and some of the mesoderm was pigmented, i.e., tissue of the host embryo (). The dorsal lip of the blastopore acts as the primary organizer of the embryo after it is carried inside the gastrula to form a layer of chordamesoderm (notochord) beneath the ectoderm. The contact thus established between these two layers results in an induction by the chordamesoderm of nervous tissue in the overlying ectoderm. Spemann and Mangold concluded that: “The piece of the upper blastoporal lip exerts an organizing effect on its surrounding in such a way that when transplanted into an indifferent location of another embryo, it leads to the formation of a secondary embryo anlage.”
Figure 3. (A and B) The Spemann-Mangold organizer experiment. A graft, taken from near the blastopore of a pigmented amphibian embryo, is transplanted to a different site in an un-pigmented embryo; the transplant initiates a secondary invagination and gastrulation, which eventually results in an entire secondary larva. The graft derived tissues are perfectly integrated anatomically with host cells as shown in the transparent view of the secondary tail region (bottom right).
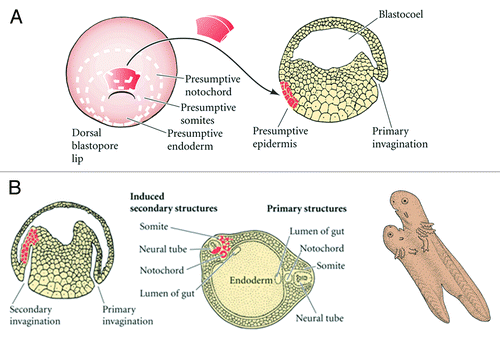
The participation of a dorsal blastopore lip in the program of development of a new embryo may be regarded in three significant aspects: (1) Formation of axial structures and establishment of the plane of bilateral symmetry; (2) Redetermination of fate for the party of host cells and their inclusion in the structures induced by the transplant; (3) Induction of the nervous system.
Spemann referred to the dorsal lip cells and their derivatives (notochord, prechordal mesoderm) as the organizer because they induced the host's ventral tissues to change their fates to form a neural tube and dorsal mesodermal tissue (such as somites), and they organized host and donor tissues into a secondary embryo with clear anterior-posterior and dorsal-ventral axes. He proposed that during normal development, these cells organize the dorsal ectoderm into a neural tube and transform the flanking mesoderm into the anterior-posterior body axis.
Because the fate of the transplanted cells could therefore be traced during development, Spemann and Mangold were able to demonstrate that the graft became notochord, yet induced neighboring cells to change fates. These neighboring cells adopted differentiation pathways that were more dorsal, and produced tissues such as the central nervous system, somites and kidneys.
In 1933, Otto Mangold published a work in which he demonstrated that the invaginating mesoderm, the archenteron roof, not only led to the neural determination of the ectoderm but also caused the regional specialization of the induced neural plate.Citation2 He dissected the archenteron roof of a young neurula into four consecutive segments and implanted into the blastocoel of an early gastrula. The anterior most mesoderm induced cement glands and eyes, posterior-most mesoderm induced tails, and the intermediate mesoderm induced the appropriate intermediate structures. These results indicate clearly how the different territories of the inductor induced different secondary structures in a cranio-caudal fashion.
In other words, the experiments performed by Mangold demonstrated that the involuted dorsal mesoderm is responsible for both the induction and patterning of the nervous system. In fact, transplantation of blastopore lips into the blastocoels cavity of a host embryo was shown to result in ectopic neural structures in the tadpole. Interestingly, the age of the blastopore lip determined the antero-posterior character of the induced ectopic neural structures. Thus, transplants of early lips, which contain the cells of head mesoderm exclusively, induced anterior neural tissues such as brain and eyes, while transplants of late lips, which in addition to head mesoderm contain cells of chordal mesoderm, induced mostly posterior neural tissue such as spinal cord.
In later experiments, devoted to the study of induction in other regions of the embryo, Spemann again found that what is induced usually is an organ, with its own characteristic shape. But some of these experiments, in which fragments from frogs’ eggs were transplanted to newts’ eggs, or viceversa, led to what should probably be considered Spemann’s second major contribution: the discovery that the character of the induced organ depends much more on its own intrinsic (presumably genetic) constitution than on that of the inducer.
In 1935, Spemann received the Nobel prize for Physiology or Medicine, the only one ever to be awarded to an embryologist. Unfortunately, Hilde Mangold had died earlier at the age of only 26, in a kitchen stove accident while warming milk for her recently born baby. In 1938, Spemann published is famous book “Embryonic Development and Induction.”
Subsequent lineage-tracing experiments demonstrated that while the head mesoderm and notochord of the second axis were derived from the transplanted lip, most of the neural tissue was comprised of induced host cells.Citation3 Moreover, further experiments showed that the organizer can also induce neural structures when recombined in vitro with competent ectoderm and confirmed that the dorsal lip is a potent source of signals for neural induction.Citation4
The Inducing Stimulus is Not Specific
Many studies have suggested that the inducing stimulus is not specific. In fact, amphibian ectoderm capable of response can be induced to form neural tissue by an array of inductors, including: (1) Living organizers from other vertebrate embryos; (2) Extracts from whole embryos or parts of embryos; (3) Tissues from embryonic or adult bodies of many kinds of animals; (4) Certain chemicals compounds.
Spemann himself as inclined to the conclusion that the inducing stimulus is nonspecific.Citation5 Johannnes Holtfreter (),Citation6 a student of Spemann’s, working independently in the laboratory of Otto Mangold in Berlin, showed that the ability to induce neural and mesodermal structures is present throughout the animal kingdom. He showed that non-inducing tissues such as ectoderm and endoderm of the gastrula, when killed, acquire the ability to induce the formation of neural plate from indifferent ectoderm. Denaturation of the proteins presumably liberates the inducing agent from an inactive complex.
Figure 4. A portrait of Johannes Holtfreter.

Moreover, Holtfreter provoked the induction effect merely by mechanical damage with glass needles and observed that prolonged boiled of a tissue or heating at 135 °C reduced the activity of the inductor and a temperature of 172 °C completely abolished it, and pointed out that neutralization of competent ectoderm “can be achieved by the application of various agents which have not more in common than the faculty of increasing the permeability of the cell membrane, and of causing cytolisis if applied in excess.”Citation7 In the 1940s, Barth and Holtfreter discovered that ectodermal cells of Amblystoma maculatum could differentiate into neural tissue without the need for any organizer or inducing substance, simply by culturing explants in sub-optimal saline solutions.Citation7,Citation8
Toward the end of 1933 some important communications were published, from which it appeared that the inductive substance occurs not only in some embryonic cell groups but also in many organs of adult invertebrate and vertebrate animals, but is not observed in vegetable substances. Subsequent studies along this line led ChuangCitation9,Citation10 and ToivonenCitation11 to the discovery that various tissues of various animals bring forth inductions which are regionally specific. For example, mouse kidney is a strong inductor of brain and sense organs, but does not induce mesodermal structures, whereas liver is effective in eliciting muscle and notochord. ChuangCitation9 observed the induction of muscle, nothocord, pronephros, and mesodermal structures by fresh mouse kidney. Induction of notochord and pronephros failed if the kidney had boiled for 5 min and no mesodermal structures were obtained if it had been boiled for 15 min.
It is very likely that also in normal embryonic development the indifferent material must be activated by factors emanating from its immediate neighborhood. In the experiment this activation can be performed by various substances. During the ten following years the leading scientists of the time studied a tremendous number of different substances in order to ascertain the chemical nature of inducers.
The Chemical Nature of the Factor Responsible for Embryonic Induction
A chemical stimulus which is relatively stable in extreme temperatures and in fat solvents, rather than mere physical stimulation, seems implicated in neural induction.
Several years after the discovery of the organizer action by Spemann and Mangold, a vivid interest arose in the chemical nature of the factor responsible for embryonic induction. Different groups of workers set out to subject embryonic amphibian tissues and inductive adult organs to various extraction methods in the hope that the active agents could be isolated and chemically identified. In addition, a large number of well characterized chemical compounds were tested.
Two prominent groups of workers, the Cambridge group, including J. Needham, C.H. Waddington and others (Waddington, who was the prime experimental embryologist in Britain, joined Needham and together they led the biochemical analysis of the organizer phenomena), and the Freiburg group, including F.G. Fischer, E. Wehmeier, F.E. Lehmann, and others conducted extensive research in the field. The Cambridge group reported results suggesting that the active compound might be a sterol, while the German school obtained inductions with various acidic preparations, including nucleic acid, oleic acid, and linoleic acid. However, in other laboratories, neural inductions were obtained with non-sterol chemicals: fatty acids, nucleoproteins, nucleic acids, adenylic acid, ATP, were among the active substances.Citation12
Needham during the 1920s and 1930s studied embryonic metabolism and the clinical properties of the developing egg (), and in 1931 published a monograph entitled “Chemical Embryology” (Cambridge University Press) summarizing knowledge on the biochemistry of the developing embryo ().Citation13 Needham began exploring the physic-chemical basis of embryonic induction at a time when scientific industrial interests in sterol hormones and vitamins expanded significantly. Needham postulated that the cells of the dorsal lip possess the inducing steroid substance in a bound form, probably in a polysaccharide-protein-sterol complex.Citation14
Figure 5. A portrait of Jospeh Needham in his laboratory.

Figure 6. The cover of the monograph entitled “Chemical Embryology” published by Joseph Needham in 1930.

In the course of gastrulation, the characteristic metabolism of the organizer breaks down the complex, releasing the sterol which induces the overlying ectoderm to from a neural tube. Evidence presented by Needham includes the following points: (1) Pure sterols or sterol like-compounds acted as evocators (Waddington termed this activity the “evocator,” and the molecules specifying the type of neural tissue were referred to as the “individuators”) of a neural response in competent ectoderm; (2) The dosage of an active steroid required for induction was very low.
Needham’s research reflected the interests of biochemists in hormone and cancer research, offering novel theoretical models and experimental methods for engaging with the function of the hormones and carcinogens they isolated.
Brachet’s Nucleoprotein Theory
J. Brachet obtained results strongly supporting his view that RNA is the basic factor in embryonic induction and differentiation. In sectioned amphibian embryos stained with pyronin or toluidine blue, Brachet found an abundance of basophilic cytoplasmic element whose stainability was lost after treatment with ribonuclease, suggesting that the basophilia was due mainly to the presence of RNA.
According to Brachet, the inducing substance may be a nucleoprotein released from the mesoderm in the form of granules which include other substances, possibly enzymes.Citation15 These granules are then engulfed by the ectoderm within which the neutralization is initiated. Another possibility is that the metabolism of the mesoderm splits the nucleoprotein into mononucleotides, which become the activating agents when transferred to the ectoderm. The evidence presented by Brachet for a relationship between production and ribonucleic acid include: (1) Grafts showed a decrease in cytoplasmic basophilia in those instances in which they act as an inductor but not in the absence of a response by the ectoderm overlying the graft. Cytoplasmic basophilia is indicative of the presence of ribonucleic acid; (2) Ectoderm which become induced to form a neural tube exhibited increased cytoplasmic basophilia, but not if neutralization fails; (3) Breakdown of the nucleoprotein by thermal or enzymatic methods abolished its inducing power.
Several objections have been made, however, to the nucleoprtient theory. Brachet himself could not excluded the possibility that the neutralizing even effect, even after RNA and nucleotides, was due to a “relay mechanism.”
Concluding Remarks
The classic work of Hans Spemann and his school demonstrated two fundamental properties of embryonic cells: (1) They are not necessarily predetermined and can be experimentally diverted from their normal developmental pathway, and (2) Cells within an embryonic organism require extrinsic messages in order to express their developmental capacities.
Further efforts in the 40s–60s led to the formation of the “double-gradient hypothesis” according to which there are two basic inducing structures—“neural” and “mesodermal.” It was supposed that both inducers may act jointly in various concentrations and form the complete spectrum of central nervous system.Citation16 In 1962, Waddington reinterpreted induction in terms of molecular biology; he linked embryonic induction to enzymatic induction.Citation17 Inducible enzymes had been called adaptive enzymes until the early 1950s, and their relationship to development had been proposed by Jacques Monod () as early as 1947.Citation18
Figure 7. A portrait of Jaçques Monod.

According to the view of embryonic induction that has emerged from classical experimental embryology, inductive interactions establish a spatially and temporally dynamic region of developmental potency, referred to as a morphogenetic field. The term 'developmental potency' can be defined as the range of differentiative capabilities that can be expressed in a given tissue under a variety of experimental conditions, as well as in vivo.Citation19
Although the organizer was shown to be present in various species, the nature of its signals remained obscure until experiments in the frog Xenopus laevis revealed the molecular events that might underlie the phenomenon of neural induction in vertebrates.Citation20-Citation22 Several molecules with direct neural activity (that is, molecules that induce the formation of neural tissue in the ectoderm without the concomitant formation of mesoderm) were described, including noggin,Citation23,Citation24 follistatin,Citation22 and chordin.Citation25,Citation26
The basic problem to be further elucidate is the following: how does a cell become adjusted to the synchronized development of an entire multicellular organism so as to express its genetic information at a strictly controlled time and place? To do this, a cell must be able to respond to (and to emit) almost two different signals: (1) At the organism and organ level, each cell must sense its position and respond to a control system (positional information, a matter developed by Lewis Wolpert []);Citation27 (2) At the tissue and cell level, each cell must exchange messages with both like and unlike adjacent cells. Such homo- and heterotypic cell interactions represent a central guiding mechanism for cytodifferentiation and morphogenesis.
Figure 8. A portrait of Lewis Wolpert.

The major criticism to induction and related concepts is that they—as Wolpert has sustained—“have completely obscured the problems of pattern formation; the failure of inductive theory to consider the problem of spatial organization;…and I regard the misure of concepts of induction as a major feature preventing progress in understanding pattern formation.”
Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
References
- Spemann H, Mangold H. Über Induktion von Embryonal anlagen durch Implantation arfrämder organisatoren. Arch. für Entwickl. der Organismen. 1924; 100:599 - 638
- Mangold O. Über die Induktionsfähikeit der verschidenen Bezirke der Neurula von urodelen. Naturwissenchaften 1933; 21:761 - 6; http://dx.doi.org/10.1007/BF01503740
- Gimlich RL, Gerhart JC. Early cellular interactions promote embryonic axis formation in Xenopus laevis. Dev Biol 1984; 104:117 - 30; http://dx.doi.org/10.1016/0012-1606(84)90042-3; PMID: 6203792
- Kintner CR, Dodd J. Hensen’s node induces neural tissue in Xenopus ectoderm. Implications for the action of the organizer in neural induction. Development 1991; 113:1495 - 505; PMID: 1811955
- Spemann H. Embryonic development and induction. 1938. New Haven, Connecticut: Yale University Press.
- Holtfreter J.. Über die Verbreitung induzierender Substanzen und ihre Leistungen im Triton-Keim. Roux Arch EntwMech Organ 1934; 132:307 383
- Holtfreter J. Neural differentiation of ectoderm through exposure to saline solution. J Exp Zool 1944; 95:307 43; http://dx.doi.org/10.1002/jez.1400950303
- Barth LG. Neural diffferentiation without organizer. J Exp Zool 1941; 87:371 - 83; http://dx.doi.org/10.1002/jez.1400870303
- Chuang HH. Induktionsleistungen von frischem und gekochten Organteilen (Niere, Leber) nach ihrer Verpflanzung in Explantate und Verschiedene Wirtsregionen von Tritonkeimen. Roux Arch EntwMech Organ 1939; 139:556 - 638; http://dx.doi.org/10.1007/BF00578755
- Chuang HH. Weitere Versuche über die Veränderung der Induktionsleisungen von gekochten. Roux Arch EntwMech Organ 1940; 140:25 38
- Toivonen S.. Über die Leistunsspezifität der abnormen Induktoren im Implantatversuch bei Triton. Ann Acad Sci fenn Ser 1940; A55:1 150
- Fischer FG, Wehmeier E, Lehmann H, Jühling L, Hultzsch K. Zur Kenntnis der Induktionsmittel in der Embryonal-Entwicklung. Berichte Chem Gesell 1935; 68:1196 - 9
- Needham J. Chemical Embryology. 3 vols. 1931; Cambridge, Cambridge University Press.
- Needham J. Biochemistry and Morphogenesis. 1942; Cambridge, Cambridge University Press.
- Brachet J. Nucleic acids in the cell and the embryo. Symp Soc Exp Biol 1947; 1:207 - 24; PMID: 20257027
- Saxen L, Toivonen S. Primary Embryonic Induction. 1962; Academic Press, London.
- Waddington CH. New Patterns in Genes and Development. 1962; Columbia University Press, NY.
- Monod J. The phenomenon of enzymatic adaptation and its bearing on problems of genetics and cellular differentiation. Growth Symp 1947; 11: 223-289.
- Slack JMW. From Egg to Embryo. Determinative Events in Early Development.1983; Cambridge, Cambridge University Press.
- Hemmati-Brivanlou A, Melton DA. A truncated activin receptor inhibits mesoderm induction and formation of axial structures in Xenopus embryos. Nature 1992; 359:609 - 14; http://dx.doi.org/10.1038/359609a0; PMID: 1328888
- Hemmati-Brivanlou A, Melton DA. Inhibition of activin receptor signaling promotes neuralization in Xenopus.. Cell 1994; 77:273 - 81; http://dx.doi.org/10.1016/0092-8674(94)90319-0; PMID: 8168134
- Hemmati-Brivanlou A, Kelly OG, Melton DA. Follistatin, an antagonist of activin, is expressed in the Spemann organizer and displays direct neuralizing activity. Cell 1994; 77:283 - 95; http://dx.doi.org/10.1016/0092-8674(94)90320-4; PMID: 8168135
- Smith WC, Harland RM. Expression cloning of noggin, a new dorsalizing factor localized to the Spemann organizer in Xenopus embryos. Cell 1992; 70:829 - 40; http://dx.doi.org/10.1016/0092-8674(92)90316-5; PMID: 1339313
- Lamb TM, Knecht AK, Smith WC, Stachel SE, Economides AN, Stahl N, Yancopolous GD, Harland RM. Neural induction by the secreted polypeptide noggin. Science 1993; 262:713 - 8; http://dx.doi.org/10.1126/science.8235591; PMID: 8235591
- Sasai Y, Lu B, Steinbeisser H, De Robertis EM. Regulation of neural induction by the Chd and Bmp-4 antagonistic patterning signals in Xenopus.. Nature 1995; 376:333 - 6; http://dx.doi.org/10.1038/376333a0; PMID: 7630399
- Piccolo S, Sasai Y, Lu B, De Robertis EM. Dorsoventral patterning in Xenopus: inhibition of ventral signals by direct binding of chordin to BMP-4. Cell 1996; 86:589 - 98; http://dx.doi.org/10.1016/S0092-8674(00)80132-4; PMID: 8752213
- Wolpert L. Positional information and pattern formation. Curr Top Dev Biol 1971; 6:183 - 224; http://dx.doi.org/10.1016/S0070-2153(08)60641-9; PMID: 4950136