Abstract
The vertebrate eye consists of multiple tissues with distinct embryonic origins. To ensure formation of the eye as a functional organ, development of ocular tissues must be precisely coordinated. Besides intrinsic regulators, several extracellular pathways have been shown to participate in controlling critical steps during eye development. Many components of Wnt/Frizzled signaling pathways are expressed in developing ocular tissues, and substantial progress has been made in the past few years in understanding their function during vertebrate eye development. Here, I summarize recent work using functional experiments to elucidate the roles of Wnt/Frizzled pathways during development of ocular tissues in different vertebrates.
Introduction
The eye originates from the neuroepithelium of the ventral forebrain, which differentiates into distinct ocular tissues, a process involving derivatives from mesoderm, neural crest and ectoderm. Critical steps during eye development are the formation of the eye field, specification of the neural retina and retina pigment epithelium (RPE), proliferation, neurogenesis, lamination and development of retinal connectivity, differentiation of the lens, ciliary body and iris, as well as vasculogenesis. Developmental disruptions of any of these steps can result in severe eye defects and ultimately result in blindness. Thus, the development of ocular tissues must be precisely coordinated, controlled by intrinsic and extracellular factors (e.g., hedgehog, FGF, TGFβ).Citation1 Wnt/Frizzled pathways play important roles in tissue specification and polarity, cellular proliferation and differentiation, axonal outgrowth as well as cellular maintenance, especially in the central nervous system (CNS). A detailed update of Wnt signaling pathways is provided elsewhere in this issue and on the Wnt website (www.stanford.edu/~rnusse/wntwindow.html). Several ocular tissues express multiple components of Wnt/Frizzled pathways during development.Citation2–Citation14 Substantial progress has been made in the past few years in understanding the role of Wnt signaling during vertebrate eye development. Gain and loss of function-studies reveal that Wnt/Frizzled pathways are involved in coordinating critical processes during ocular tissue development, which will be summarized here. I will refer to the β-catenin-independent Wnt pathways that activate JNK or Rho-Rac (PCP pathway), or intracellular calcium release, as non-canonical Wnt signaling pathways.
Specification of the Anterior Neural Plate
The forebrain originates from the anterior neuroectoderm during gastrulation and is further specified into regional subdomains including the presumptive eye. Several studies, for example in the gastrulating chick embryo and in cell culture experiments, suggest that Wnt/β-catenin signaling is active in an increasing anterior to posterior gradient along the brain to generate posterior neural tissue.Citation15–Citation19 Consistent with this model, Wnt/β-catenin signaling is not detectable in the most rostral parts of the forebrain, the telencephalon and rostral diencephalon.Citation11 To maintain anterior neural tissue, expression of secreted antagonists (SFRP1, Tlc, Dkk-1) or intracellular inhibitors (ICAT, TCF-3/headless, Axin1/masterblind, Shisa) of the Wnt/β-catenin pathway are required in the anterior neural plate ().Citation17,Citation20–Citation24 Loss of function of these inhibitors or ectopic activation of Wnt/β-catenin signaling in zebrafish and frog usually results in posteriorization and often truncation of the forebrain, and consequently decreasing or ablating expression of eye-specific genes. One possible explanation is that different levels of Wnt/β-catenin activation can specify different brain regions, with the lowest level required for telencephalic fate and highest levels for hindbrain specification.Citation18 Together, these studies suggest that distinct mechanisms precisely coordinate Wnt/β-catenin pathway activation at an appropriate level in the anterior neural plate to permit specification of forebrain subdomains. However, in mammals this model needs to be explored in further detail. Homozygous mutations of Shisa, SFRPs and other Wnt antagonists do not exhibit obvious forebrain defects, which could result from redundancy between different Wnt antagonists.Citation25–Citation27
Formation of the Eye Field
Ocular development starts with the formation of a single field of eye precursors in the anterior neural plate during late gastrulation (). This field expresses eye field transcription factors (EFTFs; e.g., Rx, Pax6, Six3, Optx2) that establish a genetic network to control eye field specification, which requires concomitant inhibition of BMP, Nodal and Wnt/β-catenin signaling in the anterior neural plate.Citation28–Citation31 EFTFs can directly prevent activation of Wnt/β-catenin signaling. The homeobox transcription factor Six3 is initially expressed in the anterior neural plate and becomes later restricted to the eye field.Citation32,Citation33 Six3 prevents activation of the Wnt/β-catenin pathway in the rostral forebrain of mouse by binding to and repressing the 3′Wnt1 enhancer.Citation34 Consistent with this, loss of Six3 function results in upregulation of Wnt1 expression and truncation of rostral forebrain structures including the eye field. Conversely, overexpression of mouse Six3 can rescue the headless phenotype in zebrafish, which is caused by mutation of the Wnt/β-catenin repressor TCF-3.Citation24,Citation34 However, for formation of the eye field, low levels of Wnt/β-catenin signaling could be required. One report shows that pygopus-2β, confirmed by overexpression experiments to be a nuclear co-activator for Wnt/β-catenin activation, is necessary for eye development.Citation35 Pygopus-2β morpholino injection leads to midbrain and forebrain defects, including in the eye, as well as downregulation of expression of the EFTFs Rx and Pax6. While a Wnt/β-catenin-independent role of pygopus-2β is possible, another explanation could be that pygopus-2β maintains Wnt/β-catenin signaling at a low but significant level in the presumptive eye field.
Several lines of evidence indicate that non-canonical Wnt signaling is essential for formation and/or maintenance of the eye field, acting to mediate morphogenetic movements. Lee et al. (2006) showed in frog that crosstalk of intracellular ephrinB1 and PCP/JNK signaling is required for retinal progenitors to execute movement into the eye field.Citation36 Activation of non-canonical Wnt signaling by Wnt11 contributes to morphogenetic movements as shown in both in frog and zebrafish.Citation36,Citation37 In zebrafish, ectopic Wnt11 induces bigger eyes and mediates strong intercellular adhesion while loss of function of Wnt11 (silberblick mutation) results in severe defects such as partial fusion of the eye field, possibly caused by defects in morphogenetic movements.Citation22,Citation37 In fish, Fzd5 is a good candidate for a receptor to mediate the effects of Wnt11, and it is proposed that non-canonical Wnt signaling antagonizes Wnt/β-catenin signaling directly, as shown previously in Wnt5/pipetail mutant zebrafish embryos ().Citation37,Citation38 In addition, it is suggested that secreted frizzled related protein 1 (SFRP1), which is expressed in the eye field in Medaka, may enhance Wnt11 signaling by further refining and separating the forebrain into telencephalic and eye precursor domains.Citation20 In Xenopus, another non-canonical Wnt ligand, Wnt4, is expressed adjacent to the eye field, and loss-of-function experiments result in a dramatic and specific loss of the eye field markers Rx and Pax6.Citation39 A potential candidate for mediating Wnt4 effects is Fzd3; low doses of Wnt4 and Fzd3 expression constructs can synergistically rescue the Wnt4 loss-of-function phenotype and Fzd3 is sufficient and required for eye formation in Xenopus.Citation39,Citation40 Furthermore, the metastasis-associated kinase (MAK) has recently been identified as a novel potential downstream effector of Fzd3 for non-canonical Wnt/JNK activation during eye field formation.Citation41 Finally, gain of function experiments show that activation of non-canonical Wnt signaling can directly upregulate expression of EFTFs. For example, overexpression of Fzd3 or Wnt11 induces ectopic expression of the EFTFs Rx and Pax6 in frog and zebrafish.Citation37,Citation40 Thus, non-canonical Wnt signaling may act through distinct mechanisms to promote formation of the eye field.
It is not known whether the role of non-canonical Wnt signaling during eye field formation is conserved in other vertebrates; for example, no obvious early eye phenotypes have been reported when Wnt4, Wnt11 or Fzd3 function are disrupted in mouse.Citation42–Citation45 In frog, Fzd5 function has been correlated with Wnt/β-catenin signaling, but Fzd5 is not expressed in the eye field and is not required for Pax6 or Rx expression but instead regulates neurogenesis in the retina at a later stage (see below).Citation46 In mouse, Fzd5 is not required for eye field formation, since the optic vesicle forms normally in Fzd5 null mice.Citation47 This further suggests that the function of individual Wnts and Frizzleds can be highly context- and species-dependent.
Neural Retina
Upon neurulation, the eye primordia evaginate and form two optic vesicles. Interaction of the distal optic vesicle with the overlying surface ectoderm results in invagination leading to formation of the optic cup (). The inner layer of the optic cup develops into the neural retina, whereas the outer layer differentiates into the non-neural retinal pigment epithelium (RPE).
Dorsoventral specification.
To follow active Wnt/β-catenin signaling, constructs with multimerized TCF/LEF binding sites have been used to generate transgenic reporters. Analysis of TCF/LEF reporters in zebrafish, frog, chick and mouse show activation of Wnt/β-catenin signaling in distinct regions of the optic vesicle and optic cup.Citation9–Citation11,Citation46,Citation48–Citation50 In chick and mouse, TCF/LEF reporters are activated in the dorsal optic vesicle.Citation11,Citation48,Citation51 In mouse, TCF/LEF reporter expression disappears in this domain when the Wnt/β-catenin co-receptor LRP6 is ablated. Disruption of LRP6 function also results in loss of the dorsal marker Tbx5, suggesting that the Wnt/β-catenin pathway might control dorsoventral patterning in the optic vesicle.Citation11
Proliferation and neurogenesis.
In frog, TCF/LEF reporter activation occurs in retinal progenitor cells of the central embryonic retina and ciliary margin zone.Citation46 Blocking the Wnt/β-catenin pathway results in loss of reporter expression as well as loss of the neural competence transcription factor Sox2. This leads to reduced proliferation, loss of proneural gene expression and a bias toward non-neural fate in progenitors of the central retina. Here, Fzd5 was identified as a good candidate to activate the Wnt/β-catenin pathway.Citation46 Another positive cofactor of Wnt/β-catenin signaling, the Dsh-binding protein Frodo, is required for eye development in frog and may support Fzd5/Wnt/β-catenin signaling in regulating retinal competence.Citation52 Finally, interference with Wnt/β-catenin signaling by overexpression of the secreted inhibitors Frzb and SFRP2 induces severe retinal defects in frog embryos.Citation53 Together, these studies demonstrate that the Wnt/β-catenin pathway is required and sufficient to control proliferation and neurogenesis in the developing frog retina. However, in mouse Fzd5, it does not activate Wnt/β-catenin signaling or expression of the retinal competence factor Sox2, in contrast to frog.Citation46,Citation47,Citation54 These observations suggest that the function of Fzd5 and Wnt/β-catenin signaling in regulating neurogenesis in the vertebrate retina is species-dependent. For example in frog, eye development occurs very rapidly, so that additional mechanisms must be employed to allow coordinated neurogenesis in the developing retina.Citation55
While ectopic expression of β-catenin in the developing retina can activate TCF/LEF reporters and affect proliferation (e.g., zebrafish), loss of function studies suggest that the role of Wnt/β-catenin signaling is not conserved across vertebrates.Citation56 In chick and zebrafish, TCF/LEF reporter activation is generally not or barely detectable in the central retina.Citation48–Citation50 In mouse, TCF/LEF reporter activation in the central retina varies depending on the transgene and it is not clear whether this expression pattern depends on Wnt/β-catenin signaling.Citation9,Citation10,Citation51 Since TCF/LEF reporter activation can be inconsistent, it raises the question of how faithful these reporters are.Citation57 For example, reporter activation could occur due to position effects of transgene insertion sites. Furthermore, previous studies revealed β-catenin-independent activation of TCF/LEF transcription factors, indicating that TCF/LEF reporter expression may not always reflect Wnt/β-catenin signaling.Citation57–Citation60 Thus, individual TCF/LEF reporters in mouse may be activated independently of Wnt/β-catenin in the retina. Indeed, loss of function of β-catenin in vivo and in vitro does not affect proliferation or differentiation of retinal cell types, suggesting that the Wnt/β-catenin pathway is not required during mammalian neurogenesis.Citation61,Citation62 However, the Wnt/β-catenin pathway may in fact play a role in retinal regeneration in mammals, since it promotes stem cell properties of adult Muller glia in vivo and in vitro.Citation63,Citation64 Furthermore, activation of TCF/LEF reporters in the developing and mature rodent retina could reflect a novel, unexpected role of TCF/LEF transcription factors during retinal development that needs further exploration.
Regulation of axonal outgrowth.
Modulation of β-catenin and LEF function in mouse in vivo and in vitro revealed that β-catenin, but not LEF, is required for correct lamination and localization of retinal cells, which is consistent with the function of β-catenin as an adherens junctions protein.Citation61,Citation62 However, in mouse explant cultures, ectopic activation of β-catenin and LEF inhibits neurite outgrowth of retinal neurons.Citation62 It is suggested that Wnt/Frizzled signaling might be involved in the regulation of axonal guidance of the projection neurons in the retina (retinal ganglion cells; RGCs) as shown for other regions of the CNS.Citation44,Citation45,Citation61,Citation62,Citation65–Citation67 When RGC axons reach their first CNS target, a Wnt3 gradient participates in regulating medial-lateral topographic mapping by repulsion or attraction depending on the receptor, counteracting EphrinB family-regulated guidance.Citation68,Citation69 For example, high Wnt3 concentrations medially result in repulsion of ventral axons mediated by the atypical receptor tyrosine kinase Ryk, which is a novel high-affinity Wnt-receptor that activates the Wnt/β-catenin pathway.Citation68,Citation70
In contrast, initial RGC outgrowth may be regulated by non-canonical Wnt signaling. For example, Wnt5a and Wnt7b are expressed in the developing retina and promote RGC axon outgrowth of chick retinal neurons.Citation9,Citation66 Similarly, in vivo and in vitro assays in chick and frog demonstrate that SFRP1 interacts with Fzd2 to promote neurite outgrowth and modulate growth cone behavior, which is dependent on pertussis toxin-sensitive Gα protein activation.Citation66 Thus, it is possible that SFRP1 acts in parallel with Wnts to activate non-canonical signaling and to promote RGC axon outgrowth in the retina. In addition, SFRP1, some Wnts and Frizzled receptors can be significantly expressed in the developing inner nuclear and outer nuclear layers.Citation6,Citation9,Citation10,Citation13 Thus, it would be interesting to further explore whether non-canonical Wnt signaling acts in outgrowth of neuronal processes and synaptogenesis of other retinal neurons besides ganglion cells, as shown in other neuronal systems.Citation71–Citation73 Interestingly, retinal cells in culture are responsive to treatment with SFRP1 and Wnt4, although no required role of these Wnt components has been observed.Citation74,Citation75
Ciliary Body and Iris
The peripheral rim of the optic cup forms the ciliary margin, which differentiates into the ciliary body as well as iris ( and ). The ciliary body and the most peripherally located iris comprise the nonpigmented and pigmented ciliary epithelium that extends from the retina and RPE, respectively. It has been suggested that the ciliary body is specified at the optic vesicle stage in a transition zone between RPE and retina, which is regulated by a tightly coordinated gradient of FGF and TGFβ family factors.Citation76–Citation78 Furthermore, retinal stem cells have been identified in the ciliary body and iris.Citation79,Citation80
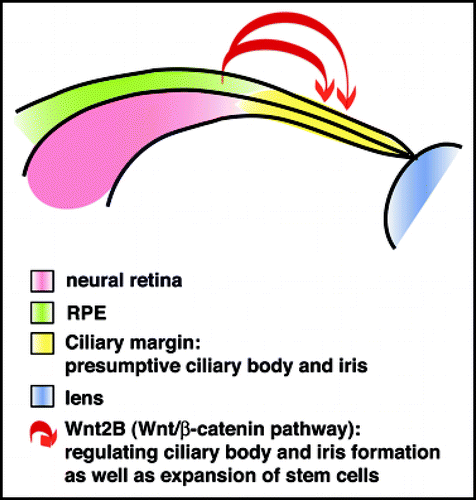
Across vertebrates, all TCF/LEF reporters show consistent activation in the developing ciliary margin.Citation9,Citation10,Citation46,Citation48–Citation50,Citation81 As mentioned above, reporter expression is already detectable at the optic vesicle stage in the dorsalmost region, suggesting that Wnt/β-catenin in addition to FGF and TGFβ factors could participate in specification of the ciliary body. Indeed, overexpression of constitutively active β-catenin results in ectopic ciliary body formation and ectopic TCF/LEF reporter activation in the developing retina while retinogenic genes such as Chx10, Pax6 and Notch1 are downregulated.Citation48,Citation81 Consistent with this, disruption of β-catenin in mouse or transfection with dominant-negative LEF1 in chick results in severe defects in ciliary body formation.Citation48,Citation81 Wnt2B was identified as one candidate ligand in this process in chick.Citation48 Thus, Wnt/β-catenin signaling is sufficient and required for formation of peripheral ocular tissues. Furthermore, these studies also demonstrate that the presumptive retina is capable of transdifferentiation into ciliary body and iris.
The adult ciliary margin contains stem cells that are quiescent in vivo but can be expanded by Wnt/β-catenin pathway activation.Citation82–Citation86 Wnt2B is a good candidate to maintain stem cells in the peripheral retina, since it is expressed in the ciliary margin in different vertebrates. In chick, ectopic expression of Wnt2B maintains proliferation and inhibits proneural gene expression in retinal progenitors, with Fzd4 suggested as a candidate receptor.Citation87,Citation88 Thus, Wnt/β-catenin signaling may function in the ciliary margin not only to establish ciliary body and iris formation but also to keep progenitor cells in an undifferentiated state to provide a constant pool of stem cells ().
The membrane-type frizzled related protein MFRP is expressed in the ciliary body and RPE in mammals, however, it is not clear whether it functions in one of the Wnt pathways. Mutations in Mfrp can cause angle-closure glaucoma and nanophthalmos in humans, suggesting a role in anterior segment development.Citation89 The mouse retinal degeneration mutant Rd6 is caused by a mutation of the Mfrp gene but exhibits no anterior segment defects, indicating a different functional role of Mfrp in mice and humans.Citation90,Citation91
Lens
Interactions between the distal optic vesicle and overlying lens ectoderm are required for thickening of the ectoderm to form the lens placode.Citation92 During optic cup formation, the lens placode invaginates and closes resulting in separation of the lens vesicle. Cells in the posterior lens vesicle differentiate into primary lens fibers, while the anterior lens develops into the anterior lens epithelium. The anterior lens epithelium proliferates throughout life and produces secondary lens fiber cells that are added to the posterior compartment just below the lens equator. This growth pattern establishes polarity of the lens. Subsequently, molecular and morphological changes of differentiating fiber cells ensure establishment of the specific optic properties of the lens.Citation93,Citation94
Conditional disruption of β-catenin in the lens ectoderm during optic cup morphogenesis results in severe defects in cell adhesion and morphogenesis of the lens.Citation51 For example, F-actin and ZO-1 complex formation becomes discontinuous upon loss of beta-catenin, while progression of cell fate in the abnormal lens structures occurs normally. Since TCF/LEF reporter expression is not observed in wildtype lens ectoderm or lens vesicle, the effect of loss of β-catenin is most likely due to cell adhesion defects during lens morphogenesis.Citation51,Citation95 In agreement with this, early stages of lens induction are not affected in LRP6 mutant eyes. Interestingly, large ectopic lens structures (lentoid bodies) were observed upon lens-specific β-catenin disruption in the adjacent nasal periocular ectoderm, which is responsive to Wnt/β-catenin signaling.Citation51,Citation95 These observations indicate that the Wnt/β-catenin pathway acts regionally within the periocular ectoderm to inhibit lens formation. Consistent with this, mis-expression of β-catenin in the presumptive lens ectoderm activates ectopic TCF/LEF reporter expression and prevents induction of lens fate.Citation51,Citation95,Citation96 Repression of the Wnt/β-catenin pathway is also required for the differentiation of another component of the anterior segment in mouse, the cornea.Citation97 Remarkably, genetic ablation of the Wnt antagonist Dkk-2 results in transformation of the corneal surface ectoderm into skin with characteristic appendages such as hair follicles and sebaceous glands.
In contrast to early lens induction, Wnt/β signaling appears to be necessary during later stages of lens development, when lens epithelium and lens fiber cells differentiate. Insertional mutation of the Wnt/β-catenin coreceptor LRP6 causes microphthalmia and coloboma with varying severity.Citation98 A more detailed analysis of LRP6 mutant eyes revealed that early lens development is delayed and that the anterior lens epithelium forms incompletely.Citation99 In agreement with this, Wnt3a-conditioned media stimulates proliferation, elongation and production of lens fiber proteins in lens epithelial cells following FGF treatment.Citation100 Together, these studies show that Wnt/β-catenin signaling needs to be first suppressed in the surface ectoderm and during lens morphogenesis, but is likely required later for proper development of the anterior lens epithelium and lens fibers. Interestingly, Wnt/β-catenin signaling has been recently suggested to promote lens regeneration in adult newt; for example, exogenous Dkk-1 prevents lens formation from the dorsal iris initiated by FGF.Citation101
Vasculature
Wnt/Frizzled signaling is highly important for vascular development and angiogenesis in the embryo, and the mammalian eye is a prominent example. Most mammals develop two vascular systems in the developing eye that arise near the optic nerve head. The extraretinal hyaloid vasculature forms a temporary capillary network, the hyaloid vessels and the pupillary membrane, which nourishes the lens, primary vitreous and retina initially and then regresses through macrophage-induced programmed cell death before eye opening.Citation102 The intraretinal vasculature develops several planar networks sequentially in the inner retinal layers that eventually reach the periphery. Angiogenic sprouting of the first innermost radial plexus provides deeper retinal layers with additional capillaries, which anastomose and undergo substantial remodeling to form horizontal plexuses.Citation103 Accumulating evidence shows that proper development of both these ocular vascular systems is tightly controlled by Wnt/Frizzled signaling.
Human diseases of the intraretinal vasculature include different forms of familial exudative vitreoretinopathy (FEVR), a hereditary disorder showing hypovascularization of the peripheral retina with variable severity. In severe cases, compensatory retinal neovascularization can occur, which may result in retinal detachment and blindness. Human mutations of the Wnt pathway components Fzd4, LRP5 and the secreted cysteine-knot protein Norrin have been linked to FEVR.Citation104–Citation107 Mouse models with mutations in Fzd4, LRP5 and Norrin generally recapitulate FEVR. For example, the two intraretinal plexuses are completely missing in Fzd4 and Norrin knockout mice.Citation105,Citation107 Fzd4 is expressed in the adult retinal vasculature and can activate Wnt/β-catenin signaling and possibly non-canonical Wnt pathways as well.Citation105,Citation107–Citation111 Norrin is a novel non-Wnt, high affinity ligand for the Fzd4 receptor that is expressed in the retina and activates the Wnt/β-catenin pathway. Several mutations in the Norrin gene can cause other vascular retinopathies such as Norrie disease, retinopathy of prematurity and Coats disease. Ectopic Norrin appears to induce ocular growth and VEGF expression and can restore normal angiogenesis in the retina of mice with a disruption of the Norrin gene.Citation112 Since vasculogenesis is highly dependent on proper guidance and migration of endothelial precursors, a possible mechanism could be that loss of Fzd4, LRP5 or Norrin disturbs migration of endothelial cells into deeper retinal layers.Citation107 The resulting defect in intraretinal vascularization could lead to hypoxia and concomitant increased production of VEGF in the retina.
Incomplete vascularization of the intraretinal vasculature often leads to failed regression of the hyaloid vasculature, as in FEVR and Norrie disease. However, a phenotype with persistent hyaloid vasculature can also arise directly from mutations of Fzd4 and LRP5. Recent studies provide a more detailed insight into the mechanisms leading to persistent vasculature in the vitreous. Fzd4, LRP5 and LEF-1 are expressed in vascular endothelial cells of hyaloid vessels, which respond to macrophage-secreted Wnt7b by activation of Wnt/β-catenin signaling as shown by TCF/LEF reporter expression.Citation113 This mechanism in combination with angiopoietin signaling induces cell death of endothelial cells to ensure vascular regression.Citation113,Citation114 Consistent with this model, mutations in Fzd4, LRP5, Norrin, Wnt7b and LEF-1 result in persistent hyaloid vessels.Citation107,Citation112,Citation113,Citation115 These studies demonstrate that Wnt/β-catenin signaling directly regulates hyaloid regression by a distinct mechanism that is independent of Wnt/β-catenin-regulated angiogenesis in the retina.
Concluding Remarks and Perspectives
Recent work clearly demonstrates that Wnt pathways are required for the regulation of virtually every step of eye development, controlling distinct processes such as tissue specification, morphogenetic movements, proliferation, differentiation and apoptosis of developing ocular tissues. However, the activity of Wnt pathways during eye development shows a high degree of complexity. Some of the observed functions may be conserved across species (e.g., specification of the anterior neural plate, specification and differentiation of the ciliary margin), while others are species-dependent (e.g., retinal neurogenesis). More complexity is added by the observation that the actual Wnt pathway component participating in one specific developmental process can differ between species (e.g., Frizzled receptors regulating eye field formation in zebrafish and frog). On the other hand, individual Wnt pathway components can exert a different function or activate even different pathways depending on the cellular context or species (e.g., Frizzled-5). Another important factor is redundancy of pathway components such as Wnt ligands and receptors that may complicate functional studies, for example in mammals. However, although further studies are needed for comparison across species, it appears that non-canonical and Wnt/β-catenin signaling share specific roles during different processes of eye development. For example, non-canonical Wnt signaling may regulate eye field formation and axonal outgrowth, while Wnt/β-catenin signaling may be required for dorsoventral specification of the optic vesicle, differentiation of ciliary body and iris as well as regenerative capability of lens and retina.
Several open questions remain. So far, the role of Wnt/β-catenin signaling has been explored more comprehensively than non-canonical pathways. One reason may be that more specific tools are available to monitor and manipulate Wnt/β-catenin signaling. In addition, due to potential crosstalk between PCP and Wnt/Ca2+ signaling or between non-canonical Wnt and other pathways (e.g., PCP and ephrinB pathways), functional studies elucidating the role of non-canonical Wnt pathways can be very challenging. Furthermore, a potential interaction between Wnt/β-catenin and non-canonical Wnt signaling has not been extensively explored during eye development. It has been proposed that non-canonical Wnt signaling might directly antagonize Wnt/β-catenin signaling during eye field formation. One possibility is that non-canonical pathways upregulate inhibitors of Wnt/β-catenin signaling. Finally, Wnt pathways may be required for proper development of ocular and extraocular tissues showing expression or activation of Wnt pathways (e.g., RPE, optic stalk, extraocular mesenchyme) but have been not explored in detail.
Figures and Tables
Figure 1 Specification of the eye field in the anterior neural plate. Wnt antagonists are expressed in the neural plate and permit development of the forebrain including the eye field by suppressing Wnt/β-catenin signaling. Very close to the caudal border of the eye field, Wnt11 and Wn4 are expressed and act through Fzd5 and Fz3 (incl. MAK) to activate non-canonical Wnt pathways in the eye field. This mechanism permits morphogenetic movements of retinal precursors into the eye field and promotes expression of eye-field-specific transcription factors, e.g., Pax6 and Rx. For further explanation, see text.
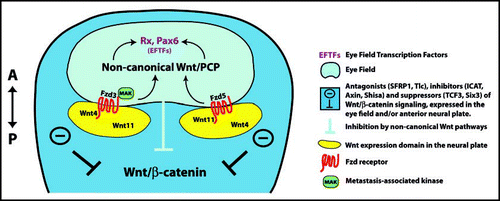
Figure 2 Morphogenesis of the optic cup. The neuroepithelium of the ventral forebrain evaginates and forms the optic vesicle, which is specified into presumptive retina, RPE and optic stalk (left). Upon interaction with the overlying lens ectoderm, the distal part of the optic vesicle (presumptive retina) invaginates, resulting in formation of the optic cup (right). The inner layer proliferates extensively and differentiates into the retina, whereas the outer layer develops into a single layer of cuboidal, pigmented cells, the RPE. The peripheral rim of the optic cup forms the ciliary margin, which differentiates into the ciliary body as well as the iris.
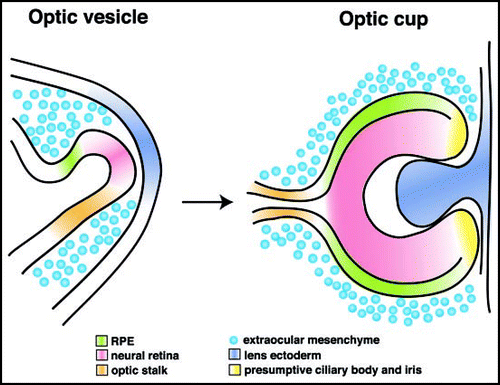
Figure 3 Wnt/β-catenin regulates differentiation of the ciliary margin and expansion of stem cells. Wnt/β-catenin signaling is activated in the ciliary margin and promotes formation of the ciliary body and iris. In addition, Wnt/β-catenin signaling increases proliferation of stem cells in the maturing or adult ciliary body and iris. Wnt2B, expressed in the dorsal RPE, is a good candidate, since it promotes ciliary body formation and maintenance of progenitors in the ciliary margin.
Acknowledgements
Many thanks to Chi-Bin Chien, Rich Dorsky, Ed Levine, Kathy Moore and Monica Vetter for critical reading and helpful suggestions on the manuscript. My apologies to those authors whose work is not cited due to space limitations.
Funded by: NIH, R01EY14954, NIH Core Grant, P30EY014800, unrestricted grant from Research to Prevent Blindness to the Department of Ophthalmology, University of Utah.
Note
This review is dedicated to the memory of my friend and colleague Dr. Ruben Adler.
References
- Adler R, Canto-Soler MV. Molecular mechanisms of optic vesicle development: complexities, ambiguities and controversies. Dev Biol 2007; 305:1 - 13
- Ang SJ, Stump RJ, Lovicu FJ, McAvoy JW. Spatial and temporal expression of Wnt and Dickkopf genes during murine lens development. Gene Expr Patterns 2004; 4:289 - 295
- Borello U, Buffa V, Sonnino C, Melchionna R, Vivarelli E, Cossu G. Differential expression of the Wnt putative receptors Frizzled during mouse somitogenesis. Mech Dev 1999; 89:173 - 177
- Filipe M, Goncalves L, Bento M, Silva AC, Belo JA. Comparative expression of mouse and chicken Shisa homologues during early development. Dev Dyn 2006; 235:2567 - 2573
- Fokina VM, Frolova EI. Expression patterns of Wnt genes during development of an anterior part of the chicken eye. Dev Dyn 2006; 235:496 - 505
- Fuhrmann S, Stark MR, Heller S. Expression of Frizzled genes in the developing chick eye. Gene Expr Patterns 2003; 3:659 - 662
- Jin EJ, Burrus LW, Erickson CA. The expression patterns of Wnts and their antagonists during avian eye development. Mech Dev 2002; 116:173 - 176
- Leimeister C, Bach A, Gessler M. Developmental expression patterns of mouse sFRP genes encoding members of the secreted frizzled related protein family. Mech Dev 1998; 75:29 - 42
- Liu H, Mohamed O, Dufort D, Wallace VA. Characterization of Wnt signaling components and activation of the Wnt canonical pathway in the murine retina. Dev Dyn 2003; 227:323 - 334
- Liu H, Thurig S, Mohamed O, Dufort D, Wallace VA. Mapping canonical Wnt signaling in the developing and adult retina. Invest Ophthalmol Vis Sci 2006; 47:5088 - 5097
- Maretto S, Cordenonsi M, Dupont S, Braghetta P, Broccoli V, Hassan AB, Volpin D, Bressan GM, Piccolo S. Mapping Wnt/beta-catenin signaling during mouse development and in colorectal tumors. Proc Natl Acad Sci USA 2003; 100:3299 - 3304
- Tissir F, Goffinet AM. Expression of planar cell polarity genes during development of the mouse CNS. Euro J Neurosci 2006; 23:597 - 607
- Van Raay TJ, Vetter ML. Wnt/frizzled signaling during vertebrate retinal development. Dev Neurosci 2004; 26:352 - 358
- Wang Y, Macke JP, Abella BS, Andreasson K, Worley P, Gilbert DJ, Copeland NG, Jenkins NA, Nathans J. A large family of putative transmembrane receptors homologous to the product of the Drosophila tissue polarity gene frizzled. J Biol Chem 1996; 271:4468 - 4476
- Lee SM, Tole S, Grove E, McMahon AP. A local Wnt-3a signal is required for development of the mammalian hippocampus. Development (Cambridge, England) 2000; 127:457 - 467
- Nordstrom U, Jessell TM, Edlund T. Progressive induction of caudal neural character by graded Wnt signaling. Nat Neurosci 2002; 5:525 - 532
- Satoh K, Kasai M, Ishidao T, Tago K, Ohwada S, Hasegawa Y, Senda T, Takada S, Nada S, Nakamura T, Akiyama T. Anteriorization of neural fate by inhibitor of beta-catenin and T cell factor (ICAT), a negative regulator of Wnt signaling. Proc Natl Acad Sci USA 2004; 101:8017 - 8021
- Wang J, Etheridge L, Wynshaw-Boris A. The Wnt-signaling pathwyas in mammalian patterning and morphogenesis. Adv Dev Biol 2007; 17:113 - 158
- Yamaguchi TP. Heads or tails: Wnts and anterior-posterior patterning. Curr Biol 2001; 11:713 - 724
- Esteve P, Lopez-Rios J, Bovolenta P. SFRP1 is required for the proper establishment of the eye field in the medaka fish. Mech Dev 2004; 121:687 - 701
- Glinka A, Wu W, Delius H, Monaghan AP, Blumenstock C, Niehrs C. Dickkopf-1 is a member of a new family of secreted proteins and functions in head induction. Nature 1998; 391:357 - 362
- Heisenberg CP, Tada M, Rauch GJ, Saude L, Concha ML, Geisler R, Stemple DL, Smith JC, Wilson SW. Silberblick/Wnt11 mediates convergent extension movements during zebrafish gastrulation. Nature 2000; 405:76 - 81
- Houart C, Caneparo L, Heisenberg C, Barth K, Take-Uchi M, Wilson S. Establishment of the telencephalon during gastrulation by local antagonism of Wnt signaling. Neuron 2002; 35:255 - 265
- Kim CH, Oda T, Itoh M, Jiang D, Artinger KB, Chandrasekharappa SC, Driever W, Chitnis AB. Repressor activity of Headless/Tcf3 is essential for vertebrate head formation. Nature 2000; 407:913 - 916
- Belo JA, Bachiller D, Agius E, Kemp C, Borges AC, Marques S, Piccolo S, De Robertis EM. Cerberus-like is a secreted BMP and nodal antagonist not essential for mouse development. Genesis 2000; 26:265 - 270
- Furushima K, Yamamoto A, Nagano T, Shibata M, Miyachi H, Abe T, Ohshima N, Kiyonari H, Aizawa S. Mouse homologues of Shisa antagonistic to Wnt and Fgf signalings. Dev Biol 2007; 306:480 - 492
- Satoh W, Gotoh T, Tsunematsu Y, Aizawa S, Shimono A. Sfrp1 and Sfrp2 regulate anteroposterior axis elongation and somite segmentation during mouse embryogenesis. Development (Cambridge, England) 2006; 133:989 - 999
- Yamamoto A, Nagano T, Takehara S, Hibi M, Aizawa S. Shisa promotes head formation through the inhibition of receptor protein maturation for the caudalizing factors, Wnt and FGF. Cell 2005; 120:223 - 235
- Zaghloul NA, Yan B, Moody SA. Step-wise specification of retinal stem cells during normal embryogenesis. Biol Cell 2005; 97:321 - 337
- Zuber ME, Gestri G, Viczian AS, Barsacchi G, Harris WA. Specification of the vertebrate eye by a network of eye field transcription factors. Development (Cambridge, England) 2003; 130:5155 - 5167
- Zuber ME, Harris WA. Formation of the eye field. Retinal Development 2006; 8 - 29
- Oliver G, Mailhos A, Wehr R, Copeland NG, Jenkins NA, Gruss P. Six3, a murine homologue of the sine oculis gene, demarcates the most anterior border of the developing neural plate and is expressed during eye development. Development (Cambridge, England) 1995; 121:4045 - 4055
- Zhou X, Hollemann T, Pieler T, Gruss P. Cloning and expression of xSix3, the Xenopus homologue of murine Six3. Mech Dev 2000; 91:327 - 330
- Lagutin OV, Zhu CC, Kobayashi D, Topczewski J, Shimamura K, Puelles L, Russell HR, McKinnon PJ, Solnica-Krezel L, Oliver G. Six3 repression of Wnt signaling in the anterior neuroectoderm is essential for vertebrate forebrain development. Genes Dev 2003; 17:368 - 379
- Lake BB, Kao KR. Pygopus is required for embryonic brain patterning in Xenopus. Dev Biol 2003; 261:132 - 148
- Lee HS, Bong YS, Moore KB, Soria K, Moody SA, Daar IO. Dishevelled mediates ephrinB1 signalling in the eye field through the planar cell polarity pathway. Nat Cell Biol 2006; 8:55 - 63
- Cavodeassi F, Carreira-Barbosa F, Young RM, Concha ML, Allende ML, Houart C, Tada M, Wilson SW. Early stages of zebrafish eye formation require the coordinated activity of Wnt11, Fz5, and the Wnt/beta-catenin pathway. Neuron 2005; 47:43 - 56
- Westfall TA, Brimeyer R, Twedt J, Gladon J, Olberding A, Furutani-Seiki M, Slusarski DC. Wnt-5/pipetail functions in vertebrate axis formation as a negative regulator of Wnt/beta-catenin activity. J Cell Biol 2003; 162:889 - 898
- Maurus D, Heligon C, Burger-Schwarzler A, Brandli AW, Kuhl M. Noncanonical Wnt-4 signaling and EAF2 are required for eye development in Xenopus laevis. EMBO J 2005; 24:1181 - 1191
- Rasmussen JT, Deardorff MA, Tan C, Rao MS, Klein PS, Vetter ML. Regulation of eye development by frizzled signaling in Xenopus. Proc Natl Acad Sci USA 2001; 98:3861 - 3866
- Kibardin A, Ossipova O, Sokol SY. Metastasis-associated kinase modulates Wnt signaling to regulate brain patterning and morphogenesis. Development (Cambridge, England) 2006; 133:2845 - 2854
- Majumdar A, Vainio S, Kispert A, McMahon J, McMahon AP. Wnt11 and Ret/Gdnf pathways cooperate in regulating ureteric branching during metanephric kidney development. Development (Cambridge, England) 2003; 130:3175 - 3185
- Stark K, Vainio S, Vassileva G, McMahon AP. Epithelial transformation of metanephric mesenchyme in the developing kidney regulated by Wnt-4. Nature 1994; 372:679 - 683
- Wang Y, Thekdi N, Smallwood PM, Macke JP, Nathans J. Frizzled-3 is required for the development of major fiber tracts in the rostral CNS. J Neurosci 2002; 22:8563 - 8573
- Wang Y, Zhang J, Mori S, Nathans J. Axonal growth and guidance defects in Frizzled3 knock-out mice: a comparison of diffusion tensor magnetic resonance imaging, neurofilament staining, and genetically directed cell labeling. J Neurosci 2006; 26:355 - 364
- Van Raay TJ, Moore KB, Iordanova I, Steele M, Jamrich M, Harris WA, Vetter ML. Frizzled 5 signaling governs the neural potential of progenitors in the developing Xenopus retina. Neuron 2005; 46:23 - 36
- Burns CJ, Zhang J, Brown EC, Van Bibber AM, Van Es J, Clevers H, Ishikawa T, Taketo MM, Vetter ML, Fuhrmann S. Investigation of Frizzled—5 during embryonic neural development in mouse. Dev Dyn 2008; 237:1614 - 1626
- Cho SH, Cepko CL. Wnt2b/beta-catenin-mediated canonical Wnt signaling determines the peripheral fates of the chick eye. Development (Cambridge, England) 2006; 133:3167 - 3177
- Dorsky RI, Sheldahl LC, Moon RT. A transgenic Lef1/beta-catenin-dependent reporter is expressed in spatially restricted domains throughout zebrafish development. Dev Biol 2002; 241:229 - 237
- Lee JE, Wu SF, Goering LM, Dorsky RI. Canonical Wnt signaling through Lef1 is required for hypothalamic neurogenesis. Development (Cambridge, England) 2006; 133:4451 - 4461
- Smith AN, Miller LA, Song N, Taketo MM, Lang RA. The duality of beta-catenin function: a requirement in lens morphogenesis and signaling suppression of lens fate in periocular ectoderm. Dev Biol 2005; 285:477 - 489
- Gloy J, Hikasa H, Sokol SY. Frodo interacts with Dishevelled to transduce Wnt signals. Nat Cell Biol 2002; 4:351 - 357
- Ladher RK, Church VL, Allen S, Robson L, Abdelfattah A, Brown NA, Hattersley G, Rosen V, Luyten FP, Dale L, Francis-West PH. Cloning and expression of the Wnt antagonists Sfrp-2 and Frzb during chick development. Dev Biol 2000; 218:183 - 198
- Taranova OV, Magness ST, Fagan BM, Wu Y, Surzenko N, Hutton SR, Pevny LH. SOX2 is a dose-dependent regulator of retinal neural progenitor competence. Genes Dev 2006; 20:1187 - 1202
- Moore KB, Schneider ML, Vetter ML. Posttranslational mechanisms control the timing of bHLH function and regulate retinal cell fate. Neuron 2002; 34:183 - 195
- Yamaguchi M, Tonou-Fujimori N, Komori A, Maeda R, Nojima Y, Li H, Okamoto H, Masai I. Histone deacetylase 1 regulates retinal neurogenesis in zebrafish by suppressing Wnt and Notch signaling pathways. Development (Cambridge, England) 2005; 132:3027 - 3043
- Barolo S. Transgenic Wnt/TCF pathway reporters: all you need is Lef?. Oncogene 2006; 25:7505 - 7511
- Hsu SC, Galceran J, Grosschedl R. Modulation of transcriptional regulation by LEF-1 in response to Wnt-1 signaling and association with beta-catenin. Mol Cell Biol 1998; 18:4807 - 4818
- Labbe E, Letamendia A, Attisano L. Association of Smads with lymphoid enhancer binding factor 1/T cell-specific factor mediates cooperative signaling by the transforming growth factor-beta and wnt pathways. Proc Natl Acad Sci USA 2000; 97:8358 - 8363
- Yi F, Merrill BJ. Stem cells and TCF proteins: a role for beta-catenin—independent functions. Stem Cell Rev 2007; 3:39 - 48
- Fu X, Sun H, Klein WH, Mu X. Beta-catenin is essential for lamination but not neurogenesis in mouse retinal development. Dev Biol 2006; 299:424 - 437
- Ouchi Y, Tabata Y, Arai K, Watanabe S. Negative regulation of retinal-neurite extension by beta-catenin signaling pathway. J Cell Sci 2005; 118:4473 - 4483
- Das AV, Mallya KB, Zhao X, Ahmad F, Bhattacharya S, Thoreson WB, Hegde GV, Ahmad I. Neural stem cell properties of Muller glia in the mammalian retina: regulation by Notch and Wnt signaling. Dev Biol 2006; 299:283 - 302
- Osakada F, Ooto S, Akagi T, Mandai M, Akaike A, Takahashi M. Wnt signaling promotes regeneration in the retina of adult mammals. J Neurosci 2007; 27:4210 - 4219
- Lyuksyutova AI, Lu CC, Milanesio N, King LA, Guo N, Wang Y, Nathans J, Tessier-Lavigne M, Zou Y. Anterior-posterior guidance of commissural axons by Wnt-frizzled signaling. Science 2003; 302:1984 - 1988
- Rodriguez J, Esteve P, Weinl C, Ruiz JM, Fermin Y, Trousse F, Dwivedy A, Holt C, Bovolenta P. SFRP1 regulates the growth of retinal ganglion cell axons through the Fz2 receptor. Nat Neurosci 2005; 8:1301 - 1309
- Rosso SB, Sussman D, Wynshaw-Boris A, Salinas PC. Wnt signaling through Dishevelled, Rac and JNK regulates dendritic development. Nat Neurosci 2005; 8:34 - 42
- Schmitt AM, Shi J, Wolf AM, Lu CC, King LA, Zou Y. Wnt-Ryk signalling mediates medial-lateral retinotectal topographic mapping. Nature 2006; 439:31 - 37
- Zou Y, Lyuksyutova AI. Morphogens as conserved axon guidance cues. Curr Opin Neurobiol 2007; 17:22 - 28
- Lu W, Yamamoto V, Ortega B, Baltimore D. Mammalian Ryk is a Wnt coreceptor required for stimulation of neurite outgrowth. Cell 2004; 119:97 - 108
- Hall AC, Lucas FR, Salinas PC. Axonal remodeling and synaptic differentiation in the cerebellum is regulated by WNT-7a signaling. Cell 2000; 100:525 - 535
- Klassen MP, Shen K. Wnt signaling positions neuromuscular connectivity by inhibiting synapse formation in C. elegans. Cell 2007; 130:704 - 716
- Salinas PC. Synaptogenesis: Wnt and TGF-beta take centre stage. Curr Biol 2003; 13:60 - 62
- Esteve P, Trousse F, Rodriguez J, Bovolenta P. SFRP1 modulates retina cell differentiation through a beta-catenin-independent mechanism. J Cell Sci 2003; 116:2471 - 2481
- Hunter DD, Zhang M, Ferguson JW, Koch M, Brunken WJ. The extracellular matrix component WIF-1 is expressed during, and can modulate, retinal development. Mol Cell Neurosci 2004; 27:477 - 488
- Dias da Silva MR, Tiffin N, Mima T, Mikawa T, Hyer J. FGF-mediated induction of ciliary body tissue in the chick eye. Dev Biol 2007; 304:272 - 285
- Hyer J. Looking at an oft-overlooked part of the eye: a new perspective on ciliary body development in chick. Dev Neurosci 2004; 26:456 - 465
- Zhao S, Chen Q, Hung FC, Overbeek PA. BMP signaling is required for development of the ciliary body. Development (Cambridge, England) 2002; 129:4435 - 4442
- Ahmad I, Das AV, James J, Bhattacharya S, Zhao X. Neural stem cells in the mammalian eye: types and regulation. Semin Cell Dev Biol 2004; 15:53 - 62
- Reh TA, Fischer AJ. Retinal stem cells. Methods in enzymology 2006; 419:52 - 73
- Liu H, Xu S, Wang Y, Mazerolle C, Thurig S, Coles BL, Ren JC, Taketo MM, van der Kooy D, Wallace VA. Ciliary margin transdifferentiation from neural retina is controlled by canonical Wnt signaling. Dev Biol 2007; 308:54 - 67
- Ahmad I, Tang L, Pham H. Identification of neural progenitors in the adult mammalian eye. Biochem Biophys Res Commun 2000; 270:517 - 521
- Asami M, Sun G, Yamaguchi M, Kosaka M. Multipotent cells from mammalian iris pigment epithelium. Dev Biol 2007; 304:433 - 446
- Inoue T, Kagawa T, Fukushima M, Shimizu T, Yoshinaga Y, Takada S, Tanihara H, Taga T. Activation of canonical Wnt pathway promotes proliferation of retinal stem cells derived from adult mouse ciliary margin. Stem Cells 2006; 24:95 - 104
- Sun G, Asami M, Ohta H, Kosaka J, Kosaka M. Retinal stem/progenitor properties of iris pigment epithelial cells. Dev Biol 2006; 289:243 - 252
- Tropepe V, Coles BL, Chiasson BJ, Horsford DJ, Elia AJ, McInnes RR, van der Kooy D. Retinal stem cells in the adult mammalian eye. Science 2000; 287:2032 - 2036
- Kubo F, Takeichi M, Nakagawa S. Wnt2b controls retinal cell differentiation at the ciliary marginal zone. Development (Cambridge, England) 2003; 130:587 - 598
- Kubo F, Takeichi M, Nakagawa S. Wnt2b inhibits differentiation of retinal progenitor cells in the absence of Notch activity by downregulating the expression of proneural genes. Development (Cambridge, England) 2005; 132:2759 - 2770
- Sundin OH, Leppert GS, Silva ED, Yang JM, Dharmaraj S, Maumenee IH, Santos LC, Parsa CF, Traboulsi EI, Broman KW, Dibernardo C, Sunness JS, Toy J, Weinberg EM. Extreme hyperopia is the result of null mutations in MFRP, which encodes a Frizzled-related protein. Proc Natl Acad Sci USA 2005; 102:9553 - 9558
- Kameya S, Hawes NL, Chang B, Heckenlively JR, Naggert JK, Nishina PM. Mfrp, a gene encoding a frizzled related protein, is mutated in the mouse retinal degeneration 6. Hum Mol Genet 2002; 11:1879 - 1886
- Mandal MN, Vasireddy V, Jablonski MM, Wang X, Heckenlively JR, Hughes BA, Reddy GB, Ayyagari R. Spatial and temporal expression of MFRP and its interaction with CTRP5. Invest Ophthalmol Vis Sci 2006; 47:5514 - 5521
- Hyer J, Kuhlman J, Afif E, Mikawa T. Optic cup morphogenesis requires pre-lens ectoderm but not lens differentiation. Dev Biol 2003; 259:351 - 363
- Lang RA. Pathways regulating lens induction in the mouse. Int J Dev Biol 2004; 48:783 - 791
- Lovicu FJ, McAvoy JW. Growth factor regulation of lens development. Dev Biol 2005; 280:1 - 14
- Kreslova J, Machon O, Ruzickova J, Lachova J, Wawrousek EF, Kemler R, Krauss S, Piatigorsky J, Kozmik Z. Abnormal lens morphogenesis and ectopic lens formation in the absence of beta-catenin function. Genesis 2007; 45:157 - 168
- Miller LA, Smith AN, Taketo MM, Lang RA. Optic cup and facial patterning defects in ocular ectoderm beta-catenin gain-of-function mice. BMC Dev Biol 2006; 6:14
- Mukhopadhyay M, Gorivodsky M, Shtrom S, Grinberg A, Niehrs C, Morasso MI, Westphal H. Dkk2 plays an essential role in the corneal fate of the ocular surface epithelium. Development (Cambridge, England) 2006; 133:2149 - 2154
- Pinson KI, Brennan J, Monkley S, Avery BJ, Skarnes WC. An LDL-receptor-related protein mediates Wnt signalling in mice. Nature 2000; 407:535 - 538
- Stump RJ, Ang S, Chen Y, von Bahr T, Lovicu FJ, Pinson K, de Iongh RU, Yamaguchi TP, Sassoon DA, McAvoy JW. A role for Wnt/beta-catenin signaling in lens epithelial differentiation. Dev Biol 2003; 259:48 - 61
- Lyu J, Joo CK. Wnt signaling enhances FGF2-triggered lens fiber cell differentiation. Development (Cambridge, England) 2004; 131:1813 - 1824
- Hayashi T, Mizuno N, Takada R, Takada S, Kondoh H. Determinative role of Wnt signals in dorsal iris-derived lens regeneration in newt eye. Mech Dev 2006; 123:793 - 800
- Ito M, Yoshioka M. Regression of the hyaloid vessels and pupillary membrane of the mouse. Anat Embryol (Berl) 1999; 200:403 - 411
- Gariano RF, Gardner TW. Retinal angiogenesis in development and disease. Nature 2005; 438:960 - 966
- Gong Y, Slee RB, Fukai N, Rawadi G, Roman-Roman S, Reginato AM, Wang H, Cundy T, Glorieux FH, Lev D, Zacharin M, Oexle K, Marcelino J, Suwairi W, Heeger S, Sabatakos G, Apte S, Adkins WN, Allgrove J, Arslan-Kirchner M, Batch JA, Beighton P, Black GC, Boles RG, Boon LM, Borrone C, Brunner HG, Carle GF, Dallapiccola B, De Paepe A, Floege B, Halfhide ML, Hall B, Hennekam RC, Hirose T, Jans A, Juppner H, Kim CA, Keppler-Noreuil K, Kohlschuetter A, LaCombe D, Lambert M, Lemyre E, Letteboer T, Peltonen L, Ramesar RS, Romanengo M, Somer H, Steichen-Gersdorf E, Steinmann B, Sullivan B, Superti-Furga A, Swoboda W, van den Boogaard MJ, Van Hul W, Vikkula M, Votruba M, Zabel B, Garcia T, Baron R, Olsen BR, Warman ML. LDL receptor-related protein 5 (LRP5) affects bone accrual and eye development. Cell 2001; 107:513 - 523
- Robitaille J, MacDonald ML, Kaykas A, Sheldahl LC, Zeisler J, Dube MP, Zhang LH, Singaraja RR, Guernsey DL, Zheng B, Siebert LF, Hoskin-Mott A, Trese MT, Pimstone SN, Shastry BS, Moon RT, Hayden MR, Goldberg YP, Samuels ME. Mutant frizzled-4 disrupts retinal angiogenesis in familial exudative vitreoretinopathy. Nat Genet 2002; 32:326 - 330
- Toomes C, Bottomley HM, Jackson RM, Towns KV, Scott S, Mackey DA, Craig JE, Jiang L, Yang Z, Trembath R, Woodruff G, Gregory-Evans CY, Gregory-Evans K, Parker MJ, Black GC, Downey LM, Zhang K, Inglehearn CF. Mutations in LRP5 or FZD4 underlie the common familial exudative vitreoretinopathy locus on chromosome 11q. Am J Hum Genet 2004; 74:721 - 730
- Xu Q, Wang Y, Dabdoub A, Smallwood PM, Williams J, Woods C, Kelley MW, Jiang L, Tasman W, Zhang K, Nathans J. Vascular development in the retina and inner ear: control by Norrin and Frizzled-4, a high-affinity ligand-receptor pair. Cell 2004; 116:883 - 895
- Masckauchan TN, Kitajewski J. Wnt/Frizzled signaling in the vasculature: new angiogenic factors in sight. Physiology (Bethesda) 2006; 21:181 - 188
- Masckauchan TN, Kitajewski J. Wnt pathways in angiogenesis. Adv Dev Biol 2007; 17:224 - 238
- Mikels AJ, Nusse R. Purified Wnt5a protein activates or inhibits beta-catenin-TCF signaling depending on receptor context. PLoS Biol 2006; 4:115
- Umbhauer M, Djiane A, Goisset C, Penzo-Mendez A, Riou JF, Boucaut JC, Shi DL. The C-terminal cytoplasmic Lys-thr-X-X-X-Trp motif in frizzled receptors mediates Wnt/beta-catenin signalling. EMBO J 2000; 19:4944 - 4954
- Ohlmann A, Scholz M, Goldwich A, Chauhan BK, Hudl K, Ohlmann AV, Zrenner E, Berger W, Cvekl A, Seeliger MW, Tamm ER. Ectopic norrin induces growth of ocular capillaries and restores normal retinal angiogenesis in Norrie disease mutant mice. J Neurosci 2005; 25:1701 - 1710
- Lobov IB, Rao S, Carroll TJ, Vallance JE, Ito M, Ondr JK, Kurup S, Glass DA, Patel MS, Shu W, Morrisey EE, McMahon AP, Karsenty G, Lang RA. WNT7b mediates macrophage-induced programmed cell death in patterning of the vasculature. Nature 2005; 437:417 - 421
- Rao S, Lobov IB, Vallance JE, Tsujikawa K, Shiojima I, Akunuru S, Walsh K, Benjamin LE, Lang RA. Obligatory participation of macrophages in an angiopoietin 2-mediated cell death switch. Development (Cambridge, England) 2007; 134:4449 - 4458
- Kato M, Patel MS, Levasseur R, Lobov I, Chang BH, Glass DA 2nd, Hartmann C, Li L, Hwang TH, Brayton CF, Lang RA, Karsenty G, Chan L. Cbfa1-independent decrease in osteoblast proliferation, osteopenia and persistent embryonic eye vascularization in mice deficient in Lrp5, a Wnt coreceptor. J Cell Biol 2002; 157:303 - 314