Abstract
Despite their devastating impact, no effective therapeutic yet exists for prion diseases at the symptomatic stage in humans or animals. Progress is hampered by the difficulty in identifying compounds that affect PrPSc and the necessity of any potential therapeutic to gain access to the CNS. Synthetic polymers known as dendrimers are a particularly promising candidate in this area. Studies with cell culture models of prion disease and prion infected brain homogenate have demonstrated that numerous species of dendrimers eliminate PrPSc in a dose and time dependent fashion and specific glycodendrimers are capable of crossing the CNS. However, despite their potential a number of important questions remained unanswered such as what makes an effective dendrimer and how dendrimers eliminate prions intracellularly. In a number of recent studies we have tackled these questions and revealed for the first time that a specific dendrimer can inhibit the intracellular conversion of PrPC to PrPSc and that a high density of surface reactive groups is a necessity for dendrimers in vitro anti-prion activity. Understanding how a therapeutic works is a vital component in maximising its activity and these studies therefore represent a significant development in the race to find effective treatments for prion diseases.
Dendrimers
Dendrimers are repetitively branched synthetic molecules with a wide variety of biological applications. They generally consist of a core from which monomers branch out in a symmetrical, structured fashion, resulting in a spherical three dimensional morphology (). The number of branch points defines the generation of dendrimer while the surface functional groups are the primary factor in determining the ionic charge. Like many great discoveries, dendrimers anti-prion activity was serendipitously discovered when it was found that the poly(amidoamine) (PAMAM) dendrimer in a transfection reagent eliminated prions from ScN2a cells.Citation1 Since then numerous other dendrimer species have demonstrated anti-prion activity including phosphorous dendrimers, maltose based glycodendrimers (mPPI), Poly(propyleneimine) (PPI) and the dendrimer like hyperbranched polymer poly(ethyleneimine) (PEI).Citation1-Citation5 The ability of these dendrimers to eliminate PrPSc from infected cells is dose and time dependent and increases with the generation number of the dendrimer. In addition, mPPI, PPI, PAMAM and high MW PEI are all capable of eliminating protease resistant PrPSc from RML infected brain homogenate, making these synthetic polymers the only known therapeutics to be effective against prions in both an intracellular and in vitro setting.Citation1,Citation3,Citation4
Figure 1. Poly(amidoamine) dendrimer (PAMAM) structure and nomenclature
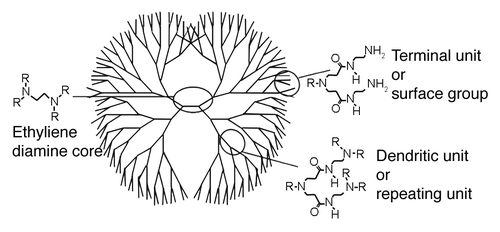
Despite their immense potential, many crucial questions surrounding dendrimer prion interactions remained unanswered at the time we began our study. For example the chemical properties of a dendrimer which define its anti-prion activity and the exact mechanism by which dendrimers eliminate PrPSc intracellularly were both unknown. In a number of recent manuscripts we have addressed these questions and advanced our understanding of the complex factors governing dendrimer/prion interactions.
Properties Which Govern Anti-Prion Activity of Dendrimers
The anti-prion efficacy of a dendrimer was traditionally believed to be based upon its cationic charge density.Citation6 However in a recent study we demonstrated that anionic, cationic and polar dendrimers are all capable of exhibiting anti-prion activity and that a high density of reactive surface groups is a necessity.Citation7
To achieve this, we produced a number of novel dendrimers through the surface group modification of a neutral maltose poly(propyleneimine) dendrimer (mPPI). The modified dendrimers varied in size, structure, charge and surface group composition and were examined against a range of prion strains to determine the effect of the modifications on dendrimer efficacy. Surprisingly, we found that dendrimers with an anti-prion effect could be cationic, anionic or neutral in charge as long as a high density of reactive surface groups was present, indicating dendrimers may interact with patches of minor surface charge on the PrPSc molecule opposite to the major surface charge. Support for dendrimers performing such a feat can be found in previous studies with proteins other than PrP.Citation8 Even more surprising was the observation that the same prion strains were always the most susceptible to elimination, regardless of dendrimer charge. This suggests that upon interaction with protease resistant PrPSc, dendrimers destabilize the protein through a somewhat generic mechanism. One possible explanation is that dendrimers act as water structure perturbing solutes—a highly destabilizing event for most protein tertiary structures. Again support for dendrimers achieving this feat can be found in previous studies with proteins other than PrP.Citation9
Having established the properties of a dendrimer which influence its in vitro anti-prion activity, we next investigated the efficacy of a particularly promising maltose dendrimer in an intracellular setting.
Anti-Prion Efficacy of mPPIg5
Maltose poly(propyleneimine) generation five (mPPIg5) is a particularly promising dendrimer with remarkably low cytotoxicity.Citation3 We investigated its efficacy in eliminating PrPSc from scrapie infected neuroblastoma cells. This is traditionally assessed by monitoring the elimination of protease resistant PrPSc over time through the use of PK digestion (to remove PrPC) and immunoblotting (to detect the remaining protease resistant PrPSc). As immunoblotting is a laborious technique and at best semi-quantitative, we developed a high throughput and highly quantitative assay known as the modified scrapie cell assay (mSCA), to measure the efficacy of anti-prion drugs in a cell culture setting.Citation10 The mSCA is based upon the standard scrapie cell assay (sSCA), an Elispot based assay for determining the level of infectious prions in a sample.Citation11 In the sSCA, susceptible cells are exposed to prion-containing samples, grown to confluence and passaged three times. The cells are subsequently attached to an Elispot plate and undergo numerous treatments including a protease digestion step to remove PrPC and an immuno-detection step to detect protease resistant PrPSc. We altered the protease digestion step and some of the incubation stages of the sSCA to produce an assay more suitable for detecting a decrease in prion infected cells over time following treatment with a therapeutic agent. The result was the creation of the mSCA, a high-throughput and fully quantitative method for measuring the efficacy of anti-prion drugs. This assay was used to demonstrate that despite its low cytotoxicity, mPPIg5 is a highly efficient anti-prion therapeutic with an activity comparable to the more established anti-prion compounds STI571 and suramin.Citation10
Intracellular Method of Action of mPPIg5
Having established the efficacy of mPPIg5, we next set about determining its intracellular mode of action. Previous studies suggested that dendrimers with an anti-prion effect act directly on PrPSc in lysosomes to render them sensitive to proteases. This assumption was based on three principle pieces of evidence:
Dendrimers (PPI) localize in lysosomes.Citation4
The alkalising lysomotrophic agent chloroquine, inhibits the elimination of prions by branched polyamines (PEI).Citation1
The in vitro elimination of protease resistant PrPSc by dendritic polymers (PEI and PPI) was greatest at an acidic pH.Citation1,Citation4
Collectively, these findings certainly suggest that dendrimers with anti-prion activity achieve their effect by increasing the degradation of PrPSc within lysosomes. However when examined individually, each of the arguments is less robust. For example, PPI dendrimers localizing predominantly in lysosomes is not surprising as most endocytosed materials will end up there. Chloroquine’s inhibitory effect on branched polyamines (PEI) cannot be automatically attributed to a rise in lysosomal pH as it has numerous other effects on cells such as altering protein trafficking and endocytosis.Citation12 Furthermore, NH4Cl, another alkalising lysomotrophic agent, does not inhibit the effect of PEICitation1 suggesting alkalisation of acidic compartments alone is not sufficient. Finally, the finding that the in vitro elimination of protease resistant PrPSc by dendrimers was greatest at an acidic pH is also not conclusive as the in vitro anti-prion action of dendrimers appears to involve the denaturation of PrPSc and so, an acidic pH enhancing this is not surprising. In addition, an acidic pH is not necessary for the in vitro anti-prion effect dendrimers, it just optimizes it, and so the necessity of an acidic environment intracellularly cannot be automatically assumed.Citation7,Citation13 Most pertinently, a small number of dendrimers, including mPPIg5, have been shown to be effective in an intracellular environment but not always in an in vitro one, even when tested against the same prion strain.Citation10,Citation14 If dendrimers do indeed act directly against protease resistant PrPSc to render it sensitive to protease degradation, how can one explain a dendrimer working intracellularly on PrPSc but not in vitro? Was it possible that dendrimers act through a different mechanism intracellularly? To examine this and to determine how mPPIg5 was eliminating prions from cells, we took advantage of the unique intracellular processing of PrPSc.
Exploiting the Intracellular Cleavage of PrPSc
PrPSc has residues 23–88 cleaved from its N-terminus approximately one hour after its production.Citation15 The truncated PrPSc formed by this event is known as PrP27-30 and accumulates in lysosomes.Citation15 For the sake of clarity, we use the term “Full Length PrPSc (FL PrPSc)” to refer to non-truncated protease resistant PrPSc and PrP27-30 to refer to truncated protease resistant PrPSc. PrPSc is used to refer to all disease isoforms of the prion protein.
In summary: PrPC → FL PrPSc → PrP27-30
As FL PrPSc is short lived, its intracellular level is closely related to its production rate. If the synthesis of FL PrPSc is interfered with, the level of detectable FL PrPSc drops rapidly. Thus the level of FL PrPSc can be seen as a marker of the conversion of PrPC to misfolded forms. NH4CL treatment of cells inhibits the truncation of FL PrPSc.Citation15 Thus in prion infected cells NH4Cl treatment leads to a rapid increase in FL PrPSc as long as the conversion of PrPC to FL PrPSc is not inhibited.
We exploited this fact to determine how a drug eliminates PrPSc from cells. We reasoned, if a compound inhibits the conversion of PrPC to FL PrPSc, no difference between cells treated in the presence and absence of NH4Cl should be observed; FL PrPSc is not being produced so NH4Cl induced inhibition of truncation has no effect.
On the other hand, if an anti-prion compound solely achieves its effect by increasing the degradation of PrPSc isoforms (including FL PrPSc), prion infected cells treated in the presence of NH4Cl should produce a higher level of FL PrPSc than cells treated in the absence of NH4Cl. There are two possible reasons for this. (1) NH4Cl may raise the pH of the lysosome and consequently inhibit the anti-prion action of a drug. In this instance, PrPSc isoforms (including FL PrPSc) would not be degraded and so an increase in the level of FL PrPSc would be observed in cells treated in the presence of NH4Cl over cells treated in the absence of NH4Cl. (2) In the presence of NH4Cl, an anti-prion drug may still degrade PrPSc isoforms. However NH4Cl treated cells will possess an increased level of FL PrPSc. Cells treated with an anti-prion drug in the presence of NH4Cl will therefore display a higher level of FL PrPSc than cells treated in the absence of NH4Cl as there is more FL PrPSc to degrade.
Thus it can be seen that by comparing a compounds effect on cellular levels of FL PrPSc in the presence and absence of NH4Cl over a short time frame, we were able to determine whether a drug increases the degradation of PrPSc or inhibits the conversion of PrPC to the misfolded PrPSc.
nSCA for Determining Compounds Mode of Action
To produce a method capable of quantifying the level of FL PrPSc in treated cells, the N-terminal specific scrapie cell assay was created (nSCA). This is another modified version of the sSCA and specifically detects FL PrPSc—it does not detect PrPC or PrP27-30. Two drugs with a known method of action, suramin and STI571, were used to validate the assay and demonstrate that our system is capable of determining an anti-prion therapeutics’ mode of action. Finally, the system was used to examine how mPPIg5 eliminates PrPSc from cells. The assay demonstrated that mPPIg5 is a highly potent drug which inhibits the conversion of PrPC to PrPSc—the first time this has been demonstrated for a dendrimer. Through the use of pulse chase, confocal microscopy and phosphatidylinositol-specific phospholipase C we demonstrated that mPPIg5 does not interfere with the biogenesis or processing of the regular PrP isoform. Thus it can be seen that maltose dendrimers are a highly promising class of therapeutics, which specifically target misfolded PrP and prevent the conversion of PrPC to PrPSc.Citation10
There are a number of possible explanations for how mPPIg5 achieves this. mPPIg5 may competitively inhibit interactions between PrPC and PrPSc. It may interfere with PrPSc trafficking to such an extent that it is no longer available for the conversion of PrPC. Alternatively it may interfere with components of PrPSc formation which have not yet been identified such as short-lived PrPC → PrPSc intermediates or potential co-factors like RNA which are necessary for PrPSc formation.Citation16 However, in our opinion the most likely explanation is that mPPIg5 alters the structure of PrPSc to such an extent that it is no longer capable of initiating the mis-folding of PrPC.
Evidence for Dendrimers Altering Structure of PrPSc
Evidence for mPPIg5 and other dendrimers effecting the structure of PrPSc comes from in vitro studies with prion infected brain homogenate, performed in our laboratory and elsewhere, demonstrating that various dendrimers appear to denature protease resistant PrPSc and render it susceptible to proteolysis.Citation1,Citation4,Citation7,Citation13 Additionally, electron microscopy and Fourier transformed infrared spectroscopy studies have demonstrated that PPI dendrimer treatment of purified PrPSc leads to disaggregation and a loss in β-sheet structure.Citation4 It is possible that this change in conformation inhibits PrPSc’s ability to convert PrPC to misfolded forms. This is supported by in vitro fibrilization studies that demonstrate dendrimer treatment of PrP peptides inhibits their ability to aggregate.Citation2
Dendrimers altering the conformation of PrPSc may explain all anti-prion activity that has been reported for dendrimers in both an in vitro and intracellular setting. One can envisage dendrimers interacting with PrPSc (both intracellularly and in vitro) and altering its structure in a process that is governed by the original confirmation of the PrPSc molecule (influenced by the type of prion strain)Citation13 and on the chemical nature of the dendrimer (dependant on a combination of the dendrimers size, structure, charge and density of reactive surface groups).Citation7 In some scenarios this interaction may alter the conformation of PrPSc to such an extent that it loses its ability to initiate the misfolding of PrPC and becomes susceptible to proteolysis. However, in other situations the change in conformation may be more subtle and PrPSc, while losing its ability to induce misfolding of PrPC, is not altered to the extent whereby it becomes sensitive to proteolysis. This would explain how some dendrimers fail to reduce protease resistant PrPSc in vitro but can be effective against the same prion strains intracellularly. Additionally, this model allows dendrimers to eliminate prions intracellularly by increasing their degradation as the original experiments in this area suggestCitation1,Citation4,Citation5 and/or eliminate prions by inhibiting the conversion of PrPC to PrPSc as some of the more recent studies in this area, including our own, suggest.Citation10,Citation14
Dendrimers altering the conformation of misfolded PrP may also explain in vitro studies which have shown that numerous dendrimers (including mPPI) inhibit the fibrilization of PrP peptides in vitro.Citation2,Citation17-Citation19 Dendrimers increasing the fibril breakage rate and dendrimers blocking the ends of growing fibrils are two of the explanations offered for this phenomena.Citation18 While both of these scenarios are possible in an intracellular setting, dendrimers blocking the end of growing fibrils cannot explain the in vitro ability of various dendrimers to reduce protease resistant PrPSc in prion infected brain homogenate.Citation1,Citation3-Citation5 Conversely, specific dendrimers increasing the fibril breakage rate can explain this ability and can again be linked back to a dendrimers ability to alter the conformation of misfolded PrP. While it is entirely possible that particular dendrimers achieve their anti-prion effect through a variety of mechanisms, we believe dendrimers altering the conformation of PrPSc is the most likely explanation as it can explain the anti-prion activity observed for dendrimers in a variety of settings.
Lack of mPPIg5 Effect on PrPC—Positive or Negative?
The lack of an effect of mPPIg5 on the biogenesis and processing of PrPC is particularly interesting as it can be viewed as a negative or positive feature of mPPIg5. On the positive side, the role of the highly conserved PrPC is not yet known and so alterations to its life cycle are undesirable. That said, studies with PrP−/− knockout mice demonstrate that such mice develop normally (though they are somewhat inconclusive in regards to synaptic function) and some of the most promising results in prolonging the incubation time of prion disease in animal models have involved the ablation of PrPC.Citation20 Additionally, a drugs ability to interfere with PrPC may mean it also has potential in other neurodegenerative diseases where PrPC is thought to play a role. For example PrPC is implicated in Alzheimer diseases as a potential receptor for Aβ and antibodies directed against PrPC have successfully inhibited the disruption of long-term potentiation (LTP) in rat hippocampus induced by Aβ-containing aqueous extracts of Alzheimer disease.Citation22 PrPC has even been implicated in mediating neurotoxic signaling by sensitizing cells to the toxic effects of β-sheet-rich conformers.Citation21 As far as prion diseases are concerned, mPPIg5′s specific effect on PrPSc and lack of an effect on PrPC appears to be the most promising outcome. While not necessarily the direct cause of neurodegeneration in prion diseases, PrPSc is certainly the main component of the infectious agentCitation23 and so a therapeutic that specifically eliminates PrPSc with minimum toxicity to the cell and with no detectable effect on endogenous PrPC is, from a theoretical viewpoint at least, ideal.
Dendrimer Potential in Treatment of Other Neurodegenerative Diseases
In the realm of neurodegenerative disorders, the therapeutic potential of dendrimers is not just limited to prion diseases. In vitro studies have demonstrated that dendrimers can interact with various other polypeptides and that they have a particular affinity for peptides of amyloid structure.Citation24 This suggests dendrimers have potential against a wide range of disease associated proteins such as SOD1 mutants in Amyotrophic Lateral Sclerosis, α-synuclein aggregates in Parkinson disease and Htt protein in Huntington disease. Dendrimers have already been shown to have an in vitro effect on Aβ peptides. Aβ is the main constituent of amyloid plaques in Alzheimer disease and Aβ oligomers are believed to be neurotoxic. Studies have shown that various dendrimers (including mPPI) can inhibit the in vitro fibrilization of Aβ1–28 (a fragment of Aβ) and reduce the toxicity of the fibrils formed.Citation18,Citation25-Citation27 In addition, phosphorous dendrimers have recently been shown to affect the aggregation of MAP-Tau protein.Citation27 Hyper-phosphorylated tau is a major component of the neurofibrillary tangles present in Alzheimer disease, frontotemporal dementia and the tauopathies.Citation28 The ability of dendrimers to effect the in vitro aggregation of Aβ and MAP-tau is very promising as an almost identical effect is observed for various dendrimers acting against PrP peptides.Citation2,Citation17-Citation19 This raises hopes that dendrimers may be able to accelerate the clearance and/or prevent the aggregation of a host of misfolded proteins intracellularly.
Conclusion
Dendrimer/prion interactions represent a fascinating area in neurodegenerative disease research which has allowed us to address many questions about the fundamental nature of prions. The therapeutic potential of dendrimers is immense due to the fact that numerous varieties target amyloid proteins and maltotriose modified PPI dendrimers have recently been shown to be capable of crossing the blood to brain barrier.Citation29
In our recent work we have demonstrated that anti-prion activity is not limited to cationic dendrimers and that a high density of reactive surface groups is a necessity. Furthermore we showed for the first time that a dendrimer is capable of inhibiting the intracellular conversion of PrPC to PrPSc. Understanding how a drug works is a vital component in maximising its performance. By establishing that mPPIg5 interferes with the conversion of PrPC to PrPSc, our study should help determine what other drugs may enhance this effect. Future work will require the use of animal models of prion disease to fully determine the potential of dendrimers in the treatment of prion and other neurodegenerative disorders.
| Abbreviations: | ||
| Aβ | = | Amyloid Beta peptide |
| mPPI | = | maltose modified poly(propyleneimine) dendrimer |
| mPPIg5 | = | maltose modified poly(propyleneimine) generation five dendrimer |
| PAMAM | = | poly(amidoamine) dendrimer |
| PEI | = | hyperbranched poly(ethyleneimine) |
| PPI | = | poly(propyleneimine) dendrimer |
| SOD1 | = | superoxide dismutase 1 |
| FL PrPSc | = | non-truncated protease resistant PrPSc. PrP27-30 (in this manuscript) = N-terminally truncated protease resistant PrPSc |
Acknowledgments
This work was supported by University College Dublin, the Irish Research Council for Science, Engineering and Technology (IRCSET) the European Union (Dendrimers in Biomedical Applications—COST TD0802, Neuroprion—FOOD-CT-2004–506579), the Irish Department of Agriculture, Food and Rural Development (FIRM 01-R&D-D-160), the Saxon Ministry for Science and Art and The German Ministry for Education and Science, as well as the DFG SFB 596 (to Jorg Tatzelt).
Disclosure of Potential Conflicts of Interest
No potential conflict of interest was disclosed.
References
- Supattapone S, Nguyen HO, Cohen FE, Prusiner SB, Scott MR. Elimination of prions by branched polyamines and implications for therapeutics. Proc Natl Acad Sci U S A 1999; 96:14529 - 34; http://dx.doi.org/10.1073/pnas.96.25.14529; PMID: 10588739
- Klajnert B, Appelhans D, Komber H, Morgner N, Schwarz S, Richter S, et al. The influence of densely organized maltose shells on the biological properties of poly(propyleneimine) dendrimers: new effects dependent on hydrogen bonding. Chemistry 2008; 14:7030 - 41; http://dx.doi.org/10.1002/chem.200800342; PMID: 18576443
- Fischer M, Appelhans D, Schwarz S, Klajnert B, Bryszewska M, Voit B, et al. Influence of surface functionality of poly(propyleneimine) dendrimers on protease resistance and propagation of the scrapie prion protein. Biomacromolecules 2010; 11:1314 - 25; http://dx.doi.org/10.1021/bm100101s; PMID: 20405854
- Supattapone S, Wille H, Uyechi L, Safar J, Tremblay P, Szoka FC, et al. Branched polyamines cure prion-infected neuroblastoma cells. J Virol 2001; 75:3453 - 61; http://dx.doi.org/10.1128/JVI.75.7.3453-3461.2001; PMID: 11238871
- Solassol J, Crozet C, Perrier V, Leclaire J, Béranger F, Caminade AM, et al. Cationic phosphorus-containing dendrimers reduce prion replication both in cell culture and in mice infected with scrapie. J Gen Virol 2004; 85:1791 - 9; http://dx.doi.org/10.1099/vir.0.19726-0; PMID: 15166465
- Sebestik J, Niederhafner P, Jezek J. Peptide and glycopeptide dendrimers and analogous dendrimeric structures and their biomedical applications. Amino Acids 2011; 40:301 - 70; http://dx.doi.org/10.1007/s00726-010-0707-z; PMID: 21058024
- McCarthy JM, Rasines Moreno B, Filippini D, Komber H, Maly M, Cernescu M, et al. Influence of surface groups on poly(propyleneimine) dendrimers antiprion activity. Biomacromolecules 2013; 14:27 - 37; http://dx.doi.org/10.1021/bm301165u; PMID: 23234313
- Ciolkowski M, Pałecz B, Appelhans D, Voit B, Klajnert B, Bryszewska M. The influence of maltose modified poly(propyleneimine) dendrimers on hen egg white lysozyme structure and thermal stability. Colloids Surf B Biointerfaces 2012; 95:103 - 8; http://dx.doi.org/10.1016/j.colsurfb.2012.02.021; PMID: 22410344
- Boas U, Christensen JB, Heegaard PMH. Dendrimers in medicine and biotechnology: new molecular tools. Cambridge, U.K.: Royal Society of Chemistry, 2006.
- McCarthy JM, Franke M, Resenberger UK, Waldron S, Simpson JC, Tatzelt J, et al. Anti-prion drug mPPIg5 inhibits PrP(C) conversion to PrP(Sc). PLoS One 2013; 8:e55282; http://dx.doi.org/10.1371/journal.pone.0055282; PMID: 23383136
- Klöhn PC, Stoltze L, Flechsig E, Enari M, Weissmann C. A quantitative, highly sensitive cell-based infectivity assay for mouse scrapie prions. Proc Natl Acad Sci U S A 2003; 100:11666 - 71; http://dx.doi.org/10.1073/pnas.1834432100; PMID: 14504404
- Pless DD, Wellner RB. In vitro fusion of endocytic vesicles: effects of reagents that alter endosomal pH. J Cell Biochem 1996; 62:27 - 39; http://dx.doi.org/10.1002/(SICI)1097-4644(199607)62:1<27::AID-JCB4>3.0.CO;2-3; PMID: 8836873
- McCarthy JM, Rasines B, Appelhans D, Rogers M. Differentiating prion strains using dendrimers. Adv Healthc Mater 2012; 1:768 - 72; http://dx.doi.org/10.1002/adhm.201200151; PMID: 23184829
- Lim YB, Mays CE, Kim Y, Titlow WB, Ryou C. The inhibition of prions through blocking prion conversion by permanently charged branched polyamines of low cytotoxicity. Biomaterials 2010; 31:2025 - 33; http://dx.doi.org/10.1016/j.biomaterials.2009.11.085; PMID: 20022103
- Caughey B, Raymond GJ, Ernst D, Race RE. N-terminal truncation of the scrapie-associated form of PrP by lysosomal protease(s): implications regarding the site of conversion of PrP to the protease-resistant state. J Virol 1991; 65:6597 - 603; PMID: 1682507
- Deleault NR, Kascsak R, Geoghegan JC, Supattapone S. Species-dependent differences in cofactor utilization for formation of the protease-resistant prion protein in vitro. Biochemistry 2010; 49:3928 - 34; http://dx.doi.org/10.1021/bi100370b; PMID: 20377181
- Heegaard PMH, Pedersen HG, Flink J, Boas U. Amyloid aggregates of the prion peptide PrP106-126 are destabilised by oxidation and by the action of dendrimers. FEBS Lett 2004; 577:127 - 33; http://dx.doi.org/10.1016/j.febslet.2004.09.073; PMID: 15527773
- Klajnert B, Cortijo-Arellano M, Cladera J, Bryszewska M. Influence of dendrimer’s structure on its activity against amyloid fibril formation. Biochem Biophys Res Commun 2006; 345:21 - 8; http://dx.doi.org/10.1016/j.bbrc.2006.04.041; PMID: 16674918
- Klajnert B, Cortijo-Arellano M, Cladera J, Majoral JP, Caminade AM, Bryszewska M. Influence of phosphorus dendrimers on the aggregation of the prion peptide PrP 185-208. Biochem Biophys Res Commun 2007; 364:20 - 5; http://dx.doi.org/10.1016/j.bbrc.2007.09.083; PMID: 17927954
- Colby DW, Prusiner SB. Prions. Cold Spring Harb Perspect Biol 2011; 3:a006833; http://dx.doi.org/10.1101/cshperspect.a006833; PMID: 21421910
- Resenberger UK, Harmeier A, Woerner AC, Goodman JL, Müller V, Krishnan R, et al. The cellular prion protein mediates neurotoxic signalling of β-sheet-rich conformers independent of prion replication. EMBO J 2011; 30:2057 - 70; http://dx.doi.org/10.1038/emboj.2011.86; PMID: 21441896
- Barry AE, Klyubin I, Mc Donald JM, Mably AJ, Farrell MA, Scott M, et al. Alzheimer’s disease brain-derived amyloid-β-mediated inhibition of LTP in vivo is prevented by immunotargeting cellular prion protein. J Neurosci 2011; 31:7259 - 63; http://dx.doi.org/10.1523/JNEUROSCI.6500-10.2011; PMID: 21593310
- Soto C. Prion hypothesis: the end of the controversy?. Trends Biochem Sci 2011; 36:151 - 8; http://dx.doi.org/10.1016/j.tibs.2010.11.001; PMID: 21130657
- Lane AR, Stanley CJ, Wilson SM. Binding of pathological forms of prion proteins. MICROSENS BIOPHAGE LTD (GB), 2010.
- Klajnert B, Cladera J, Bryszewska M. Molecular interactions of dendrimers with amyloid peptides: pH dependence. Biomacromolecules 2006; 7:2186 - 91; http://dx.doi.org/10.1021/bm060229s; PMID: 16827586
- Klajnert B, Cangiotti M, Calici S, Majoral JP, Caminade AM, Cladera J, et al. EPR study of the interactions between dendrimers and peptides involved in Alzheimer’s and prion diseases. Macromol Biosci 2007; 7:1065 - 74; http://dx.doi.org/10.1002/mabi.200700049; PMID: 17654761
- Wasiak T, Ionov M, Nieznanski K, Nieznanska H, Klementieva O, Granell M, et al. Phosphorus Dendrimers Affect Alzheimer’s (Aβ1–28) Peptide and MAP-Tau Protein Aggregation. Mol Pharm 2011; 9:458 - 69; http://dx.doi.org/10.1021/mp2005627
- Lee VM, Goedert M, Trojanowski JQ. Neurodegenerative tauopathies. Annu Rev Neurosci 2001; 24:1121 - 59; http://dx.doi.org/10.1146/annurev.neuro.24.1.1121; PMID: 11520930
- Janaszewska A, Ziemba B, Ciepluch K, Appelhans D, Voit B, Klajnert B, et al. The biodistribution of maltotriose modified poly (propyleneimine)(PPI) dendrimers conjugated with fluorescein—proofs of crossing blood–brain–barrier. New J Chem 2012; 36:350 - 3; http://dx.doi.org/10.1039/c1nj20444k