Abstract
A baffling aspect of metazoan proteostasis is the lack of an Hsp104 ortholog that rapidly disaggregates and reactivates misfolded polypeptides trapped in stress induced disordered aggregates, preamyloid oligomers, or amyloid fibrils. By contrast, in bacteria, protozoa, chromista, fungi, and plants, Hsp104 orthologs are highly conserved and confer huge selective advantages in stress tolerance. Moreover, in fungi, the amyloid remodeling activity of Hsp104 has enabled deployment of prions for various beneficial modalities. Thus, a longstanding conundrum has remained unanswered: how do metazoan cells renature aggregated proteins or resolve amyloid fibrils without Hsp104? Here, we highlight recent advances that unveil the metazoan protein-disaggregase machinery, comprising Hsp110, Hsp70, and Hsp40, which synergize to dissolve disordered aggregates, but are unable to rapidly solubilize stable amyloid fibrils. However, Hsp110, Hsp70, and Hsp40 exploit the slow monomer exchange dynamics of amyloid, and can slowly depolymerize amyloid fibrils from their ends in a manner that is stimulated by small heat shock proteins. Upregulation of this system could have key therapeutic applications in various protein-misfolding disorders. Intriguingly, yeast Hsp104 can interface with metazoan Hsp110, Hsp70, and Hsp40 to rapidly eliminate disease associated amyloid. Thus, metazoan proteostasis is receptive to augmentation with exogenous disaggregases, which opens a number of therapeutic opportunities.
Keywords: :
Introduction
Controlling the “quality” of proteins beyond translation is key in assuring correct cellular functioning and preserving organismal health.Citation1 For a nascent polypeptide chain, folding into the correct native structure and maintaining the correct native form is challenging within the crowded environment of a cell.Citation2-Citation4 Thus, proteins occasionally assume defective, nonfunctional conformations. To circumvent this fundamental problem of cell biology, nature has devised several mechanisms to prevent or resolve protein misfolding in the cell. For instance, molecular chaperones block protein aggregation and actively help proteins reach their native conformations.Citation5 If a protein has been damaged beyond repair, it can be cleared by specialized degradation systems.Citation6-Citation8 Alas, protein misfolding and aggregation can overcome these systems, especially upon environmental stress, which can even elicit aging and disease.Citation9 Indeed, protein misfolding underpins several devastating neurodegenerative diseases, including Alzheimer disease, Parkinson disease, Creutzfeldt-Jakob disease, and Huntington's disease.Citation10,Citation11
In the 1990s, Susan Lindquist and coworkers unequivocally defined a new branch in protein quality control: protein disaggregation coupled to protein reactivation.Citation12-Citation14 Although it had been speculated that protein disaggregation and reactivation might occur,Citation15 it had never been convincingly demonstrated. Lindquist and colleagues discovered a new heat shock protein (Hsp) in Saccharomyces cerevisiae, Hsp104, which was found to have a key role in allowing cells to survive severe stress after heat treatment (thermotolerance).Citation14 In subsequent investigations, Hsp104 was found to solubilize large protein aggregates resulting from severe heat stress and recover enzymatically active proteins from these aggregates.Citation12 Accordingly, yeast cells lacking Hsp104 were no longer able to rapidly solubilize and reactivate proteins from an aggregated state following thermal stress.Citation13,Citation16
Since then, we have learned much about the way Hsp104 functions. Hsp104 is a ring-shaped homohexamer with two AAA+ nucleotide-binding domains (NBDs) per subunit that couple ATP binding and hydrolysis to protein disaggregation.Citation17-Citation22 Hsp104 is thought to drive protein disaggregation by threading substrates through its central channel to solution.Citation23-Citation26 Hsp104 disaggregates a diverse array of structures, ranging from stable amyloid to less stable disordered aggregates.Citation12,Citation13,Citation27-Citation33 Hsp104 hexamers adapt different mechanisms of intersubunit collaboration to disaggregate stress-induced aggregates vs. amyloid.Citation27,Citation34 Hsp104 acts alone or in concert with other molecular chaperones to rescue aggregated polypeptides.Citation12,Citation20,Citation28,Citation32,Citation34-Citation36 In particular, Hsp70, Hsp40, and small heat shock proteins (Hsp26 and Hsp42) can synergize with Hsp104 to promote the reactivation of protein aggregates.Citation35,Citation37,Citation38 Hsp104 is also essential for the formation and propagation of several yeast prions; protein-based genetic elements comprised of amyloid fibers that can confer advantageous self-perpetuating changes in protein structure and function.Citation32,Citation39-Citation46
Hsp104 is highly conserved in eubacteria and eukaryotes. Inexplicably, however, Hsp104 has no exact homolog or ortholog in metazoa.Citation47 NBD2, but not other parts of Hsp104, appears to be partially conserved in the four ER-resident AAA+ proteins: torsin A, B, 2A, and 3A, as well as the mitochondrial AAA+ protein, SKD3.Citation48-Citation50 This deficiency of Hsp104 in animals is puzzling, as a protein that reverses protein aggregation and restores protein function would be pivotal in combating aberrant protein aggregation.Citation17,Citation47 The reason underlying the loss of Hsp104 is unknown, and is even more baffling because Hsp104 is well tolerated and even neuroprotective in animal systems.Citation30,Citation51-Citation55 For example, Hsp104 rescues α-synuclein aggregation and dopaminergic neurodegeneration in a rat model of Parkinson disease.Citation30 Whether mammals boast an equivalent protein disaggregase has endured as a persistent unanswered question.
Hsp110, Hsp70 and Hsp40 as a novel protein disaggregase system
We have recently shed some light on this issue and have identified the mammalian disaggregase system via biochemical fractionation of mammalian cytosol and reconstitution with pure components.Citation56 The mammalian disaggregase system is comprised of an Hsp110 (Apg-2), an Hsp70 (Hsc70 or Hsp70), and an Hsp40 (Hdj1 or Hdj2).Citation56 The combination of these three proteins was found to establish an active disaggregase system in the mammalian cytosol prepared from rat liver or sHeLa cells.Citation56 Hsp110, Hsp70, and Hsp40 were able to refold proteins from large chemically or thermally denatured protein aggregates.Citation56 Using pure proteins, we established that Hsp70 and Hsp40 alone are not sufficient for robust disaggregase activity, but must be supplemented with Hsp110.Citation56 Hsp110 homologs are found in all eukaryotes and contribute toward thermotolerance in mammalian cells.Citation57,Citation58 Hsp110 can serve as a nucleotide exchange factor (NEF) for Hsp70 but also displays chaperone activity.Citation59-Citation64 We established that Hsp110-Hsp70-Hsp40 disaggregase activity was most effective against disordered, amorphous aggregates.Citation56 Indeed, Hsp110, Hsp70 and Hsp40 were unable to rapidly disaggregate Sup35 prions or amyloid forms of α-synuclein.Citation56 Disaggregase activity was conserved to the yeast homologs.Citation56 Thus, Sse1 (Hsp110), Ssa1 (Hsp70), and Sis1 or Ydj1 (Hsp40) could synergize to rescue proteins from large disordered aggregates.Citation56 This activity was slow in comparison to Hsp104- catalyzed protein disaggregation, which might help explain why minimal disaggregase activity is observed in yeast lacking Hsp104 immediately after heat shock.Citation13 In yeast, Sse1 contributes to prion propagationCitation65-Citation67 and might also be involved in the dissolution of “Q-bodies” or “stress foci”: punctate cytoplasmic structures where misfolded proteins are collected prior to maturation into larger inclusions.Citation68-Citation70 Using a series of Sse1 mutants,Citation61 we determined that Sse1 must engage both substrate and Hsp70, promote nucleotide exchange on Hsp70, and bind and hydrolyze ATP itself to promote disaggregation of disordered aggregates. Thus, simply providing Hsp70 with a NEF, such as Fes1 or Snl1ΔN, in place of Sse1 was insufficient to promote protein disaggregation.Citation56 Likewise, using a series of Ssa1 mutants, we determined that Hsp70 must engage substrate and Hsp110, and hydrolyze ATP for protein disaggregation.Citation56 Hsp40 must harbor a functional J domain to promote protein disaggregation, but the J domain alone is insufficient.Citation56 Optimal disaggregase activity was achieved when the Hsp40 could stimulate Hsp110 and Hsp70 ATPase activity.Citation56 Finally, while Hsp110, Hsp70, and Hsp40 were unable to rapidly resolve amyloid conformers directly, they enhanced disaggregation of Sup35 prions and α-synuclein amyloid fibrils by Hsp104.Citation56
About a year later, a subsequent study confirmed the metazoan disaggregation activity exerted by Hsp110 (Apg-1, Apg-2, or Hsp105), Hsp70 (Hsc70 or Hsp70), and Hsp40 (Hdj1 or DNAJA2) in vitro.Citation71 Curiously, under the in vitro conditions employed the ATPase activity of Hsp110 was not required to promote protein disaggregation.Citation71 Hsp110 appeared to contribute primarily by acting as a nucleotide exchange factor (NEF) for Hsp70. Mild aggregation conditions were even established where Hsc70 and the alternative Hsp40, DNAJA2, could disaggregate substrates if provided with the Hsp70 NEFs Bag-1 or Snl1ΔN instead of Hsp110.Citation71 Under these circumstances, stimulation of Hsp70 nucleotide exchange was sufficient for disaggregation.Citation71
Using C. elegans as a model system, knockdown of Hsp110 in briefly heatshocked C. elegans resulted in persistent luciferase-YFP aggregates and a drastically reduced lifespan.Citation71 The persistence of protein aggregates and lifespan reduction could reflect a requirement for Hsp110 in the solubilization of protein aggregates in vivo.Citation71
Alternatively, it might point to a role for Hsp110 in the inhibition of ongoing aggregation after the transient heat shock. Unfortunately, the experiments performed could not differentiate between these two possibilities, as the expression of luciferase-YFP was not shut down after the transient heat shock. Thus, it is unclear whether the persistence of luciferase-YFP aggregates reflects a failure to disaggregate pre-existing luciferase-YFP or whether newly synthesized luciferase-YFP continued to aggregate after the transient heat shock (perhaps due to seeding by preformed aggregates).
Originally, to convincingly establish the disaggregase activity of Hsp104 in vivo, it was necessary to stringently shut down protein synthesis using cycloheximide immediately after the heat shock.Citation13 In this way, one could be absolutely certain that any protein reactivation that occurred was due to recovery of previously aggregated protein and not due to the accumulation of newly synthesized material.Citation13 A similar strategy has been employed to demonstrate that Hsp104 can solubilize amyloid in vivo.Citation29 Thus, in the study by Rampelt et al. one cannot be certain what proportion of the soluble luciferase- YFP observed in wild-type C. elegans after a 12h or 24h recovery from heat shock represents newly synthesized protein or bona fide resolubilized protein.Citation71 A similar issue arises in more recent experiments that also aimed to demonstrate in vivo disaggregase activity of Hsp110 using Drosophila S2 cells, but again the critical cycloheximide control was also omitted.Citation72 As such, although the foregoing experiments provide compelling indications,Citation71,Citation72 we still await an unequivocal in vivo demonstration of Hsp110, Hsp70, and Hsp40 disaggregase activity.
More recently, Goloubinoff and colleagues corroborated that the cytosol and the endoplasmic reticulum of mammalian cells contain Hsp110 and Hsp70 machineries that can unfold and solubilize stably misfolded and aggregated protein.Citation62 Outstandingly, Hsp110 (Hsp105) was found to be an ATP-dependent unfoldase that can prevent aggregation, catalyze the unfolding of misfolded polypeptides, and favor their conversion into native protein on its own.Citation62 Thus, Hsp110 can act as a bona fide chaperone with unfolding activity, and does not simply serve as a NEF for Hsp70.Citation62 Furthermore, titration of the ATP- and Hsp40-dependent refolding activity in the presence of various amounts of Hsp110 and Hsp70 showed optimal disaggregation activity at a 1:1 ratio.Citation62
Intriguingly, even without ATP, Hsp110 promotes the release of a prebound substrate from Hsp70, and Hsp70 could activate the release of a prebound substrate from Hsp110.Citation62 Together with our study,Citation56 these findings conflict with the notion that the only function for Hsp110 in protein disaggregation is as a NEF for Hsp70.Citation62 Indeed, it should also be noted that several key in vivo functions of Sse1 require its ATPase activity.Citation73,Citation74
Collectively, these results from three different groups independently corroborate the existence of a disaggregase system, consisting of Hsp110, Hsp70, and Hsp40, that couple protein disaggregation to protein renaturation in metazoa (). Discrepancies in the details of the system regarding the requirement for Hsp110 ATPase activity in disaggregation and the relative level of activity of the system are likely the result of different experimental conditions. For instance, we used equal concentrations of Hsp70 and Hsp110,Citation56 while Rampelt and colleagues used a 1:10 ratio of Hsp110 to Hsp70.Citation71 Analogously to our work, Goloubinoff and coworkers identified an optimal 1:1 ratio for the concentrations of Hsp110 and Hsp70, which might even suggest an Hsp110:Hsp70 heterodimer that co-operatively drives disaggregation via synergistic entropic pulling.Citation56,Citation62,Citation75-Citation77 Indeed, Sse1 has been co-crystallized in 1:1 complex with the Hsc70 nucleotide-binding domain, suggesting that a 1:1 complex could be critical.Citation78
Figure 1. Hsp110, Hsp70, Hsp40 and sHsps are a disaggregation system in metazoan cells. Heat shock proteins Hsp110, Hsp70, and Hsp40 are capable of dissolving disordered aggregates. For labile aggregates, Hsp110 may only need to operate as a nucleotide exchange factor (NEF) for Hsp70, whereas for more stable aggregates it may need to serve as a NEF for Hsp70, engage substrate, and bind and hydrolyze ATP. Hsp110, Hsp70, and Hsp40 can also slowly depolymerize ordered amyloid substrates from their ends. Rapid amyloid dissolution can be achieved by supplementing Hsp110, Hsp70 and Hsp40 with exogenous Hsp104. Here, fibrils can be fragmented and monomers extracted from anywhere in the fibril (not just the ends), which leads to more rapid dissolution. sHsps can stimulate all of these protein disaggregation reactions, but are not absolutely required.
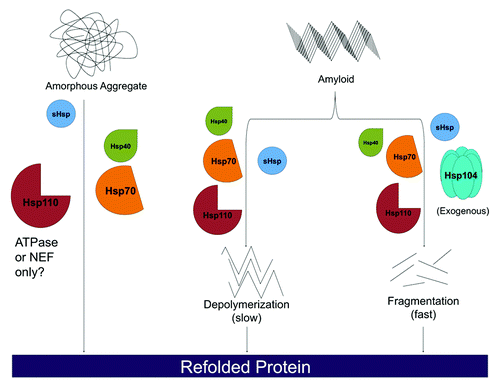
Moreover, Ssa1 and Sse1 display high affinity for different peptides,Citation79 indicating that they might interact and exert force on different regions of the polypeptide to cooperatively drive disaggregation. In each of these studies,Citation56,Citation62,Citation71 different conditions were used to generate the protein aggregates studied. For instance, in our study, more severe chemical or thermal denaturation was used to generate aggregates.Citation56 Hence, it is highly probable that different aggregated conformers were studied in each case. We suggest that proteins can adopt a wide variety of conformations in the aggregated state, some of which are more labile and do not require the full chaperone repertoire of Hsp110 for disaggregation (NEF activity is sufficient), whereas others are more recalcitrant and require the full complement of Hsp110 activities encompassing: substrate binding, Hsp70 binding, promotion of nucleotide exchange on Hsp70, and ATP binding and hydrolysis. Additional studies are required to explore this hypothesis further. However, the Hsp70 NEF, Snl1ΔN, could not substitute for Hsp110 under our more stringent aggregation conditions,Citation56 whereas it could under much milder aggregation conditions.Citation71 An interesting parallel may be drawn with Hsp104, which employs distinct mechanisms to dissolve labile aggregates vs. stable amyloid.Citation27 Hsp104 subunits within the hexamer can function independently to resolve disordered aggregates.Citation27 Thus, even a single subunit within the hexamer can drive disaggregation of disordered aggregates.Citation27 By contrast, multiple Hsp104 subunits must work together within the hexamer in a co-operative manner to drive amyloid dissolution.Citation27 Some very stable amyloid conformations even require direct co-operation between two Hsp104 hexamers.Citation27,Citation56 By analogy, the Hsp110-Hsp70-Hsp40 system might also exhibit mechanistic plasticity in protein disaggregation. We suggest that more stable aggregated structures might necessitate the full repertoire of Hsp110 modalities, whereas the NEF activity might suffice for more facile conformers.
Amyloid Disaggregation in Metazoa
In addition to amorphous aggregates, misfolded proteins in the cell can form amyloids and prions.Citation10,Citation80,Citation81 Amyloids are self-templating protein conformers.Citation10,Citation80,Citation81 They form long, stable fibers by self-replicating their “cross-β” conformation at their growing ends and by converting other copies of the same protein to the “cross-β” amyloid form.Citation10,Citation80,Citation81 When amyloid fibers become infectious, they are termed prions.Citation10,Citation80,Citation81 Initially, we found Hsp110, Hsp70, and Hsp40 were unable to rapidly remodel amyloid in the absence of Hsp104.Citation56 However, in a later study, we found Hsp110, Hsp70, and Hsp40 especially in conjunction with small heat shock proteins (sHsps), can very slowly depolymerize amyloid fibers (Sup35 prions or α-synuclein fibrils) from their ends,Citation35 providing a pathway for amyloid disaggregation in metazoans in the absence of Hsp104. Amyloid depolymerization is a lengthy process that occurs on a similar timeframe to molecular recycling within amyloid fibers (days).Citation35,Citation82,Citation83 The disaggregase system involving Hsp110, Hsp70, Hsp40, and sHsps might exploit this process to slowly eliminate amyloid by accelerating monomer dissociation or by capturing released monomers or by sealing off fibril ends once a monomer has been released thereby preventing monomer reassociation.Citation35 While newly released monomers could hypothetically collect into toxic oligomers, the chaperone system would likely prevent any toxic oligomer formation. Remarkably, we found this activity to be conserved in humans.Citation35 Thus, Hsp110 (Apg-2), Hsp70 (Hsc70), Hsp40 (Hdj1) and a small heat shock protein (HspB5) slowly depolymerized α-synuclein fibrils, which are connected to Parkinson disease.Citation19 These data suggest that in metazoa, which lack an Hsp104 homolog, Hsp110, Hsp70, and Hsp40 can slowly eliminate amyloid forms by specifically hijacking their intrinsic monomer recycling process.Citation35,Citation82,Citation83
Treating Neurodegenerative Disease: Can We Give Hsp110 a Boost?
The Hsp110-Hsp70-Hsp40 disaggregase system might prove to be an advantageous therapeutic target against the numerous neurological disorders connected to protein misfolding and aggregation.Citation10 Indeed, co-expression of Hsp110 and Hsp40 in Drosophila melanogaster suppresses the cytotoxicity of polyglutamine aggregation.Citation84 Importantly, the ATPase activity of Hsp110 was critical for this rescue.Citation84 A feasible explanation for this effect invokes Hsp110 and Hsp40 interfacing with members of the Hsp70 family to disassemble polyglutamine aggregates and thus reduce the cellular toxicity of protein aggregation. Polyglutamine aggregation is connected to several neurodegenerative diseases, including Huntington’s disease and Spinocerebellar Ataxias.Citation85 Similarly, Hsp110 was found to completely reverse a vesicle transport defect produced by a mutant (G85R) of Superoxide Dismutase 1 (SOD1) associated with amyotrophic lateral sclerosis (ALS) in the isolated axoplasm from the giant axon of the squid Loligo pealei.Citation86 Hsp110 appears to either directly bind to the mutant SOD1, or to associate with the mutant protein via Hsp70, and occlude binding surfaces that would otherwise interact with endogenous proteins leading to a gain of toxic function. Further studies are warranted to determine whether Hsp110, Hsp70, and Hsp40 can also disaggregate misfolded SOD1 conformers connected to ALS or polyglutamine fibrils and oligomers connected to Huntington’s disease and Spinocerebellar Ataxias. Treatment of several neurodegenerative disorders could entail the activation of the Hsp110-Hsp70-Hsp40 disaggregase system. As Hsp110, Hsp70, and Hsp40 enhanced amyloid remodeling by Hsp104,Citation56 one alternative possibility to achieve such activation would be to supplement metazoan cells with Hsp104.Citation47 For instance, Hsp104 prevented the aggregation and toxicity of polyglutamine in C. elegans.Citation54 In mouse and rat, Hsp104 expression resulted in extension the animal’s lifespan and rescue of striatal dysfunction respectively.Citation53,Citation55 We have recently introduced Hsp104 into Drosophila models of Spinocerebellar Ataxia Type-3.Citation51 Notably, Hsp104 suppressed toxicity of a C-terminal ataxin-3 fragment when expressed even after the onset of polyglutamine-induced degeneration.Citation51 This constitutes the first disaggregase treatment that halts disease progression after the start of pathogenic degeneration.Citation51 Notably, induction of Hsp70 after polyglutamine-mediated degeneration had already initiated was unable to significantly mitigate disease progression.Citation51 It is possible that simultaneous induction of not only Hsp70, but also Hsp110 and Hsp40 is necessary to achieve toxicity suppression after degeneration and aggregation have initiated.Citation84
Another possibility to enhance clearance of harmful amyloids would be to boost sHsp levels or activity to facilitate the action of endogenous human Hsp110, Hsp70, and Hsp40. Thus, small molecules that induce the expression of these proteins without compromising other components of the stress response could be critical.Citation11,Citation87,Citation88 For example, the ability to stimulate the dissolution of α-synuclein fibers in patients with Parkinson disease might provide an unprecedented therapeutic leap in the treatment of this disease. Although released monomers could theoretically reassemble into toxic oligomers, the proteostasis network would likely prevent this situation. Lastly, direct pharmacological activation of Hsp110 or Hsp70 is another attractive possibility.Citation89 While pharmaceutical discovery efforts generally focus on protein inhibition, protein activation is an emerging field.Citation90
In conclusion, the metazoan disaggregase machinery is comprised of the heat shock proteins Hsp110, Hsp70, and Hsp40, which dissolve disordered aggregates.Citation56,Citation62,Citation71 Hsp110, Hsp70, and Hsp40 exploit the exchange dynamics of amyloid, and can slowly depolymerize amyloid fibrils from their ends.Citation35 This amyloid depolymerase activity as well as the disaggregation of disordered aggregates is stimulated by sHsps.Citation35,Citation71 Augmentation of this disaggregase network could have key applications in various neurological disorders linked to protein misfolding. Fascinatingly, this metazoan disaggregation network is amenable to augmentation with exogenous disaggregases, which opens several exciting avenues for potential treatments.Citation47
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
We thank Laura Castellano, Jamie DeNizio, Dr Meredith Jackrel, and Elizabeth Sweeny for helpful comments on the manuscript. MPT is supported by a PENN-PORT postdoctoral fellowship (K12GM081259). JS is supported by grants: NIH Director's New Innovator Award DP2OD002177, R21HD074510, R01GM099836, a Muscular Dystrophy Association Research Award (MDA277268), Packard Center for ALS Research at Johns Hopkins University, Target ALS, and an Ellison Medical Foundation New Scholar in Aging Award.
References
- Lindquist SL, Kelly JW. Chemical and biological approaches for adapting proteostasis to ameliorate protein misfolding and aggregation diseases: progress and prognosis. Cold Spring Harb Perspect Biol 2011; 3:3; http://dx.doi.org/10.1101/cshperspect.a004507; PMID: 21900404
- Sarkar M, Smith AE, Pielak GJ. Impact of reconstituted cytosol on protein stability. Proc Natl Acad Sci U S A 2013; 110:19342 - 7; http://dx.doi.org/10.1073/pnas.1312678110; PMID: 24218610
- Uversky VN, M Cooper E, Bower KS, Li J, Fink AL. Accelerated alpha-synuclein fibrillation in crowded milieu. FEBS Lett 2002; 515:99 - 103; http://dx.doi.org/10.1016/S0014-5793(02)02446-8; PMID: 11943202
- van den Berg B, Ellis RJ, Dobson CM. Effects of macromolecular crowding on protein folding and aggregation. EMBO J 1999; 18:6927 - 33; http://dx.doi.org/10.1093/emboj/18.24.6927; PMID: 10601015
- Kim YE, Hipp MS, Bracher A, Hayer-Hartl M, Hartl FU. Molecular chaperone functions in protein folding and proteostasis. Annu Rev Biochem 2013; 82:323 - 55; http://dx.doi.org/10.1146/annurev-biochem-060208-092442; PMID: 23746257
- Wong E, Cuervo AM. Integration of clearance mechanisms: the proteasome and autophagy. Cold Spring Harb Perspect Biol 2010; 2:a006734; http://dx.doi.org/10.1101/cshperspect.a006734; PMID: 21068151
- Maday S, Holzbaur EL. Autophagosome assembly and cargo capture in the distal axon. Autophagy 2012; 8:858 - 60; http://dx.doi.org/10.4161/auto.20055; PMID: 22617438
- Maday S, Wallace KE, Holzbaur EL. Autophagosomes initiate distally and mature during transport toward the cell soma in primary neurons. J Cell Biol 2012; 196:407 - 17; http://dx.doi.org/10.1083/jcb.201106120; PMID: 22331844
- Coelho M, Dereli A, Haese A, Kühn S, Malinovska L, DeSantis ME, Shorter J, Alberti S, Gross T, Tolić-Nørrelykke IM. Fission yeast does not age under favorable conditions, but does so after stress. Curr Biol 2013; 23:1844 - 52; http://dx.doi.org/10.1016/j.cub.2013.07.084; PMID: 24035542
- Cushman M, Johnson BS, King OD, Gitler AD, Shorter J. Prion-like disorders: blurring the divide between transmissibility and infectivity. J Cell Sci 2010; 123:1191 - 201; http://dx.doi.org/10.1242/jcs.051672; PMID: 20356930
- Jackrel ME, Shorter J. Shock and awe: unleashing the heat shock response to treat Huntington disease. J Clin Invest 2011; 121:2972 - 5; http://dx.doi.org/10.1172/JCI59190; PMID: 21785212
- Glover JR, Lindquist S. Hsp104, Hsp70, and Hsp40: a novel chaperone system that rescues previously aggregated proteins. Cell 1998; 94:73 - 82; http://dx.doi.org/10.1016/S0092-8674(00)81223-4; PMID: 9674429
- Parsell DA, Kowal AS, Singer MA, Lindquist S. Protein disaggregation mediated by heat-shock protein Hsp104. Nature 1994; 372:475 - 8; http://dx.doi.org/10.1038/372475a0; PMID: 7984243
- Sanchez Y, Lindquist SL. HSP104 required for induced thermotolerance. Science 1990; 248:1112 - 5; http://dx.doi.org/10.1126/science.2188365; PMID: 2188365
- Pelham HR. Speculations on the functions of the major heat shock and glucose-regulated proteins. Cell 1986; 46:959 - 61; http://dx.doi.org/10.1016/0092-8674(86)90693-8; PMID: 2944601
- Vogel JL, Parsell DA, Lindquist S. Heat-shock proteins Hsp104 and Hsp70 reactivate mRNA splicing after heat inactivation. Curr Biol 1995; 5:306 - 17; http://dx.doi.org/10.1016/S0960-9822(95)00061-3; PMID: 7780741
- Vashist S, Cushman M, Shorter J. Applying Hsp104 to protein-misfolding disorders. Biochem Cell Biol 2010; 88:1 - 13; http://dx.doi.org/10.1139/O09-121; PMID: 20130674
- DeSantis ME, Shorter J. The elusive middle domain of Hsp104 and ClpB: location and function. Biochim Biophys Acta 2012; 1823:29 - 39; http://dx.doi.org/10.1016/j.bbamcr.2011.07.014; PMID: 21843558
- Saibil H. Chaperone machines for protein folding, unfolding and disaggregation. Nat Rev Mol Cell Biol 2013; 14:630 - 42; http://dx.doi.org/10.1038/nrm3658; PMID: 24026055
- Desantis ME, Sweeny EA, Snead D, Leung EH, Go MS, Gupta K, Wendler P, Shorter J. Conserved distal loop residues in the Hsp104 and ClpB middle domain contact nucleotide-binding domain 2 and enable Hsp70-dependent protein disaggregation. J Biol Chem 2014; 289:848 - 67; http://dx.doi.org/10.1074/jbc.M113.520759; PMID: 24280225
- Doyle SM, Genest O, Wickner S. Protein rescue from aggregates by powerful molecular chaperone machines. Nat Rev Mol Cell Biol 2013; 14:617 - 29; http://dx.doi.org/10.1038/nrm3660; PMID: 24061228
- Wendler P, Shorter J, Plisson C, Cashikar AG, Lindquist S, Saibil HR. Atypical AAA+ subunit packing creates an expanded cavity for disaggregation by the protein-remodeling factor Hsp104. Cell 2007; 131:1366 - 77; http://dx.doi.org/10.1016/j.cell.2007.10.047; PMID: 18160044
- Shorter J, Lindquist S. Navigating the ClpB channel to solution. Nat Struct Mol Biol 2005; 12:4 - 6; http://dx.doi.org/10.1038/nsmb0105-4; PMID: 15689967
- Wendler P, Shorter J, Snead D, Plisson C, Clare DK, Lindquist S, Saibil HR. Motor mechanism for protein threading through Hsp104. Mol Cell 2009; 34:81 - 92; http://dx.doi.org/10.1016/j.molcel.2009.02.026; PMID: 19362537
- Lum R, Niggemann M, Glover JR. Peptide and protein binding in the axial channel of Hsp104. Insights into the mechanism of protein unfolding. J Biol Chem 2008; 283:30139 - 50; http://dx.doi.org/10.1074/jbc.M804849200; PMID: 18755692
- Lum R, Tkach JM, Vierling E, Glover JR. Evidence for an unfolding/threading mechanism for protein disaggregation by Saccharomyces cerevisiae Hsp104. J Biol Chem 2004; 279:29139 - 46; http://dx.doi.org/10.1074/jbc.M403777200; PMID: 15128736
- DeSantis ME, Leung EH, Sweeny EA, Jackrel ME, Cushman-Nick M, Neuhaus-Follini A, Vashist S, Sochor MA, Knight MN, Shorter J. Operational plasticity enables hsp104 to disaggregate diverse amyloid and nonamyloid clients. Cell 2012; 151:778 - 93; http://dx.doi.org/10.1016/j.cell.2012.09.038; PMID: 23141537
- DeSantis ME, Shorter J. Hsp104 drives “protein-only” positive selection of Sup35 prion strains encoding strong [PSI(+)]. Chem Biol 2012; 19:1400 - 10; http://dx.doi.org/10.1016/j.chembiol.2012.09.013; PMID: 23177195
- DiSalvo S, Derdowski A, Pezza JA, Serio TR. Dominant prion mutants induce curing through pathways that promote chaperone-mediated disaggregation. Nat Struct Mol Biol 2011; 18:486 - 92; http://dx.doi.org/10.1038/nsmb.2031; PMID: 21423195
- Lo Bianco C, Shorter J, Régulier E, Lashuel H, Iwatsubo T, Lindquist S, Aebischer P. Hsp104 antagonizes alpha-synuclein aggregation and reduces dopaminergic degeneration in a rat model of Parkinson disease. J Clin Invest 2008; 118:3087 - 97; http://dx.doi.org/10.1172/JCI35781; PMID: 18704197
- Shorter J, Lindquist S. Hsp104 catalyzes formation and elimination of self-replicating Sup35 prion conformers. Science 2004; 304:1793 - 7; http://dx.doi.org/10.1126/science.1098007; PMID: 15155912
- Sweeny EA, Shorter J. Prion proteostasis: Hsp104 meets its supporting cast. Prion 2008; 2:135 - 40; http://dx.doi.org/10.4161/pri.2.4.7952; PMID: 19242125
- Shorter J, Lindquist S. Destruction or potentiation of different prions catalyzed by similar Hsp104 remodeling activities. Mol Cell 2006; 23:425 - 38; http://dx.doi.org/10.1016/j.molcel.2006.05.042; PMID: 16885031
- Doyle SM, Shorter J, Zolkiewski M, Hoskins JR, Lindquist S, Wickner S. Asymmetric deceleration of ClpB or Hsp104 ATPase activity unleashes protein-remodeling activity. Nat Struct Mol Biol 2007; 14:114 - 22; http://dx.doi.org/10.1038/nsmb1198; PMID: 17259993
- Duennwald ML, Echeverria A, Shorter J. Small heat shock proteins potentiate amyloid dissolution by protein disaggregases from yeast and humans. PLoS Biol 2012; 10:e1001346; http://dx.doi.org/10.1371/journal.pbio.1001346; PMID: 22723742
- Shorter J, Lindquist S. Hsp104, Hsp70 and Hsp40 interplay regulates formation, growth and elimination of Sup35 prions. EMBO J 2008; 27:2712 - 24; http://dx.doi.org/10.1038/emboj.2008.194; PMID: 18833196
- Cashikar AG, Duennwald M, Lindquist SL. A chaperone pathway in protein disaggregation. Hsp26 alters the nature of protein aggregates to facilitate reactivation by Hsp104. J Biol Chem 2005; 280:23869 - 75; http://dx.doi.org/10.1074/jbc.M502854200; PMID: 15845535
- Haslbeck M, Miess A, Stromer T, Walter S, Buchner J. Disassembling protein aggregates in the yeast cytosol. The cooperation of Hsp26 with Ssa1 and Hsp104. J Biol Chem 2005; 280:23861 - 8; http://dx.doi.org/10.1074/jbc.M502697200; PMID: 15843375
- Newby GA, Lindquist S. Blessings in disguise: biological benefits of prion-like mechanisms. Trends Cell Biol 2013; 23:251 - 9; http://dx.doi.org/10.1016/j.tcb.2013.01.007; PMID: 23485338
- True HL, Berlin I, Lindquist SL. Epigenetic regulation of translation reveals hidden genetic variation to produce complex traits. Nature 2004; 431:184 - 7; http://dx.doi.org/10.1038/nature02885; PMID: 15311209
- True HL, Lindquist SL. A yeast prion provides a mechanism for genetic variation and phenotypic diversity. Nature 2000; 407:477 - 83; http://dx.doi.org/10.1038/35035005; PMID: 11028992
- Halfmann R, Jarosz DF, Jones SK, Chang A, Lancaster AK, Lindquist S. Prions are a common mechanism for phenotypic inheritance in wild yeasts. Nature 2012; 482:363 - 8; http://dx.doi.org/10.1038/nature10875; PMID: 22337056
- Alberti S, Halfmann R, King O, Kapila A, Lindquist S. A systematic survey identifies prions and illuminates sequence features of prionogenic proteins. Cell 2009; 137:146 - 58; http://dx.doi.org/10.1016/j.cell.2009.02.044; PMID: 19345193
- Chernoff YO, Lindquist SL, Ono B, Inge-Vechtomov SG, Liebman SW. Role of the chaperone protein Hsp104 in propagation of the yeast prion-like factor [psi+]. [psi+] Science 1995; 268:880 - 4; http://dx.doi.org/10.1126/science.7754373; PMID: 7754373
- Liebman SW, Chernoff YO. Prions in yeast. Genetics 2012; 191:1041 - 72; http://dx.doi.org/10.1534/genetics.111.137760; PMID: 22879407
- Paushkin SV, Kushnirov VV, Smirnov VN, Ter-Avanesyan MD. Propagation of the yeast prion-like [psi+] determinant is mediated by oligomerization of the SUP35-encoded polypeptide chain release factor. EMBO J 1996; 15:3127 - 34; PMID: 8670813
- Shorter J. Hsp104: a weapon to combat diverse neurodegenerative disorders. Neurosignals 2008; 16:63 - 74; http://dx.doi.org/10.1159/000109760; PMID: 18097161
- Murdock DG, Boone BE, Esposito LA, Wallace DC. Up-regulation of nuclear and mitochondrial genes in the skeletal muscle of mice lacking the heart/muscle isoform of the adenine nucleotide translocator. J Biol Chem 1999; 274:14429 - 33; http://dx.doi.org/10.1074/jbc.274.20.14429; PMID: 10318868
- Ozelius LJ, Page CE, Klein C, Hewett JW, Mineta M, Leung J, Shalish C, Bressman SB, de Leon D, Brin MF, et al. The TOR1A (DYT1) gene family and its role in early onset torsion dystonia. Genomics 1999; 62:377 - 84; http://dx.doi.org/10.1006/geno.1999.6039; PMID: 10644435
- Périer F, Radeke CM, Raab-Graham KF, Vandenberg CA. Expression of a putative ATPase suppresses the growth defect of a yeast potassium transport mutant: identification of a mammalian member of the Clp/HSP104 family. Gene 1995; 152:157 - 63; http://dx.doi.org/10.1016/0378-1119(94)00697-Q; PMID: 7835694
- Cushman-Nick M, Bonini NM, Shorter J. Hsp104 suppresses polyglutamine-induced degeneration post onset in a drosophila MJD/SCA3 model. PLoS Genet 2013; 9:e1003781; http://dx.doi.org/10.1371/journal.pgen.1003781; PMID: 24039611
- Dandoy-Dron F, Bogdanova A, Beringue V, Bailly Y, Tovey MG, Laude H, Dron M. Infection by ME7 prion is not modified in transgenic mice expressing the yeast chaperone Hsp104 in neurons. Neurosci Lett 2006; 405:181 - 5; http://dx.doi.org/10.1016/j.neulet.2006.05.066; PMID: 16884849
- Perrin V, Régulier E, Abbas-Terki T, Hassig R, Brouillet E, Aebischer P, Luthi-Carter R, Déglon N. Neuroprotection by Hsp104 and Hsp27 in lentiviral-based rat models of Huntington’s disease. Mol Ther 2007; 15:903 - 11; http://dx.doi.org/10.1038/mt.sj.6300141; PMID: 17375066
- Satyal SH, Schmidt E, Kitagawa K, Sondheimer N, Lindquist S, Kramer JM, Morimoto RI. Polyglutamine aggregates alter protein folding homeostasis in Caenorhabditis elegans.. Proc Natl Acad Sci U S A 2000; 97:5750 - 5; http://dx.doi.org/10.1073/pnas.100107297; PMID: 10811890
- Vacher C, Garcia-Oroz L, Rubinsztein DC. Overexpression of yeast hsp104 reduces polyglutamine aggregation and prolongs survival of a transgenic mouse model of Huntington’s disease. Hum Mol Genet 2005; 14:3425 - 33; http://dx.doi.org/10.1093/hmg/ddi372; PMID: 16204350
- Shorter J. The mammalian disaggregase machinery: Hsp110 synergizes with Hsp70 and Hsp40 to catalyze protein disaggregation and reactivation in a cell-free system. PLoS One 2011; 6:e26319; http://dx.doi.org/10.1371/journal.pone.0026319; PMID: 22022600
- Easton DP, Kaneko Y, Subjeck JR. The hsp110 and Grp1 70 stress proteins: newly recognized relatives of the Hsp70s. Cell Stress Chaperones 2000; 5:276 - 90; http://dx.doi.org/10.1379/1466-1268(2000)005<0276:THAGSP>2.0.CO;2; PMID: 11048651
- Oh HJ, Chen X, Subjeck JR. Hsp110 protects heat-denatured proteins and confers cellular thermoresistance. J Biol Chem 1997; 272:31636 - 40; http://dx.doi.org/10.1074/jbc.272.50.31636; PMID: 9395504
- Raviol H, Sadlish H, Rodriguez F, Mayer MP, Bukau B. Chaperone network in the yeast cytosol: Hsp110 is revealed as an Hsp70 nucleotide exchange factor. EMBO J 2006; 25:2510 - 8; http://dx.doi.org/10.1038/sj.emboj.7601139; PMID: 16688211
- Dragovic Z, Broadley SA, Shomura Y, Bracher A, Hartl FU. Molecular chaperones of the Hsp110 family act as nucleotide exchange factors of Hsp70s. EMBO J 2006; 25:2519 - 28; http://dx.doi.org/10.1038/sj.emboj.7601138; PMID: 16688212
- Polier S, Dragovic Z, Hartl FU, Bracher A. Structural basis for the cooperation of Hsp70 and Hsp110 chaperones in protein folding. Cell 2008; 133:1068 - 79; http://dx.doi.org/10.1016/j.cell.2008.05.022; PMID: 18555782
- Mattoo RU, Sharma SK, Priya S, Finka A, Goloubinoff P. Hsp110 is a bona fide chaperone using ATP to unfold stable misfolded polypeptides and reciprocally collaborate with Hsp70 to solubilize protein aggregates. J Biol Chem 2013; 288:21399 - 411; http://dx.doi.org/10.1074/jbc.M113.479253; PMID: 23737532
- Polier S, Hartl FU, Bracher A. Interaction of the Hsp110 molecular chaperones from S. cerevisiae with substrate protein. J Mol Biol 2010; 401:696 - 707; http://dx.doi.org/10.1016/j.jmb.2010.07.004; PMID: 20624400
- Shaner L, Sousa R, Morano KA. Characterization of Hsp70 binding and nucleotide exchange by the yeast Hsp110 chaperone Sse1. Biochemistry 2006; 45:15075 - 84; http://dx.doi.org/10.1021/bi061279k; PMID: 17154545
- Fan Q, Park KW, Du Z, Morano KA, Li L. The role of Sse1 in the de novo formation and variant determination of the [PSI+] prion. Genetics 2007; 177:1583 - 93; http://dx.doi.org/10.1534/genetics.107.077982; PMID: 18039878
- Moran C, Kinsella GK, Zhang ZR, Perrett S, Jones GW. Mutational analysis of Sse1 (Hsp110) suggests an integral role for this chaperone in yeast prion propagation in vivo. G3 (Bethesda) 2013; 3:1409 - 18; http://dx.doi.org/10.1534/g3.113.007112; PMID: 23797105
- Sadlish H, Rampelt H, Shorter J, Wegrzyn RD, Andréasson C, Lindquist S, Bukau B. Hsp110 chaperones regulate prion formation and propagation in S. cerevisiae by two discrete activities. PLoS One 2008; 3:e1763; http://dx.doi.org/10.1371/journal.pone.0001763; PMID: 18335038
- Escusa-Toret S, Vonk WI, Frydman J. Spatial sequestration of misfolded proteins by a dynamic chaperone pathway enhances cellular fitness during stress. Nat Cell Biol 2013; 15:1231 - 43; http://dx.doi.org/10.1038/ncb2838; PMID: 24036477
- Spokoini R, Moldavski O, Nahmias Y, England JL, Schuldiner M, Kaganovich D. Confinement to organelle-associated inclusion structures mediates asymmetric inheritance of aggregated protein in budding yeast. Cell Rep 2012; 2:738 - 47; http://dx.doi.org/10.1016/j.celrep.2012.08.024; PMID: 23022486
- Kaganovich D, Kopito R, Frydman J. Misfolded proteins partition between two distinct quality control compartments. Nature 2008; 454:1088 - 95; http://dx.doi.org/10.1038/nature07195; PMID: 18756251
- Rampelt H, Kirstein-Miles J, Nillegoda NB, Chi K, Scholz SR, Morimoto RI, Bukau B. Metazoan Hsp70 machines use Hsp110 to power protein disaggregation. EMBO J 2012; 31:4221 - 35; http://dx.doi.org/10.1038/emboj.2012.264; PMID: 22990239
- Cherkasov V, Hofmann S, Druffel-Augustin S, Mogk A, Tyedmers J, Stoecklin G, Bukau B. Coordination of Translational Control and Protein Homeostasis during Severe Heat Stress. Curr Biol 2013; 23:2452 - 62; http://dx.doi.org/10.1016/j.cub.2013.09.058; PMID: 24291094
- Goeckeler JL, Stephens A, Lee P, Caplan AJ, Brodsky JL. Overexpression of yeast Hsp110 homolog Sse1p suppresses ydj1-151 thermosensitivity and restores Hsp90-dependent activity. Mol Biol Cell 2002; 13:2760 - 70; http://dx.doi.org/10.1091/mbc.02-04-0051; PMID: 12181344
- Hrizo SL, Gusarova V, Habiel DM, Goeckeler JL, Fisher EA, Brodsky JL. The Hsp110 molecular chaperone stabilizes apolipoprotein B from endoplasmic reticulum-associated degradation (ERAD). J Biol Chem 2007; 282:32665 - 75; http://dx.doi.org/10.1074/jbc.M705216200; PMID: 17823116
- Ben-Zvi A, De Los Rios P, Dietler G, Goloubinoff P. Active solubilization and refolding of stable protein aggregates by cooperative unfolding action of individual hsp70 chaperones. J Biol Chem 2004; 279:37298 - 303; http://dx.doi.org/10.1074/jbc.M405627200; PMID: 15201275
- De Los Rios P, Ben-Zvi A, Slutsky O, Azem A, Goloubinoff P. Hsp70 chaperones accelerate protein translocation and the unfolding of stable protein aggregates by entropic pulling. Proc Natl Acad Sci U S A 2006; 103:6166 - 71; http://dx.doi.org/10.1073/pnas.0510496103; PMID: 16606842
- Goloubinoff P, De Los Rios P. The mechanism of Hsp70 chaperones: (entropic) pulling the models together. Trends Biochem Sci 2007; 32:372 - 80; http://dx.doi.org/10.1016/j.tibs.2007.06.008; PMID: 17629485
- Schuermann JP, Jiang J, Cuellar J, Llorca O, Wang L, Gimenez LE, Jin S, Taylor AB, Demeler B, Morano KA, et al. Structure of the Hsp110:Hsc70 nucleotide exchange machine. Mol Cell 2008; 31:232 - 43; http://dx.doi.org/10.1016/j.molcel.2008.05.006; PMID: 18550409
- Goeckeler JL, Petruso AP, Aguirre J, Clement CC, Chiosis G, Brodsky JL. The yeast Hsp110, Sse1p, exhibits high-affinity peptide binding. FEBS Lett 2008; 582:2393 - 6; http://dx.doi.org/10.1016/j.febslet.2008.05.047; PMID: 18539149
- Eisenberg D, Jucker M. The amyloid state of proteins in human diseases. Cell 2012; 148:1188 - 203; http://dx.doi.org/10.1016/j.cell.2012.02.022; PMID: 22424229
- Shorter J. Emergence and natural selection of drug-resistant prions. Mol Biosyst 2010; 6:1115 - 30; http://dx.doi.org/10.1039/c004550k; PMID: 20422111
- Carulla N, Caddy GL, Hall DR, Zurdo J, Gairí M, Feliz M, Giralt E, Robinson CV, Dobson CM. Molecular recycling within amyloid fibrils. Nature 2005; 436:554 - 8; http://dx.doi.org/10.1038/nature03986; PMID: 16049488
- Carulla N, Zhou M, Giralt E, Robinson CV, Dobson CM. Structure and intermolecular dynamics of aggregates populated during amyloid fibril formation studied by hydrogen/deuterium exchange. Acc Chem Res 2010; 43:1072 - 9; http://dx.doi.org/10.1021/ar9002784; PMID: 20557067
- Kuo Y, Ren S, Lao U, Edgar BA, Wang T. Suppression of polyglutamine protein toxicity by co-expression of a heat-shock protein 40 and a heat-shock protein 110. Cell Death Dis 2013; 4:e833; http://dx.doi.org/10.1038/cddis.2013.351; PMID: 24091676
- Blum ES, Schwendeman AR, Shaham S. PolyQ disease: misfiring of a developmental cell death program?. Trends Cell Biol 2013; 23:168 - 74; http://dx.doi.org/10.1016/j.tcb.2012.11.003; PMID: 23228508
- Song Y, Nagy M, Ni W, Tyagi NK, Fenton WA, López-Giráldez F, Overton JD, Horwich AL, Brady ST. Molecular chaperone Hsp110 rescues a vesicle transport defect produced by an ALS-associated mutant SOD1 protein in squid axoplasm. Proc Natl Acad Sci U S A 2013; 110:5428 - 33; http://dx.doi.org/10.1073/pnas.1303279110; PMID: 23509252
- Calamini B, Silva MC, Madoux F, Hutt DM, Khanna S, Chalfant MA, Saldanha SA, Hodder P, Tait BD, Garza D, et al. Small-molecule proteostasis regulators for protein conformational diseases. Nat Chem Biol 2012; 8:185 - 96; http://dx.doi.org/10.1038/nchembio.763; PMID: 22198733
- Labbadia J, Cunliffe H, Weiss A, Katsyuba E, Sathasivam K, Seredenina T, Woodman B, Moussaoui S, Frentzel S, Luthi-Carter R, et al. Altered chromatin architecture underlies progressive impairment of the heat shock response in mouse models of Huntington disease. J Clin Invest 2011; 121:3306 - 19; http://dx.doi.org/10.1172/JCI57413; PMID: 21785217
- Wang AM, Miyata Y, Klinedinst S, Peng HM, Chua JP, Komiyama T, Li X, Morishima Y, Merry DE, Pratt WB, et al. Activation of Hsp70 reduces neurotoxicity by promoting polyglutamine protein degradation. Nat Chem Biol 2013; 9:112 - 8; http://dx.doi.org/10.1038/nchembio.1140; PMID: 23222885
- Zorn JA, Wells JA. Turning enzymes ON with small molecules. Nat Chem Biol 2010; 6:179 - 88; http://dx.doi.org/10.1038/nchembio.318; PMID: 20154666