Abstract
Under acidic soil conditions, aluminum (Al) becomes available to plants, which must cope with its toxicity by mechanisms involving both internal and external detoxification. Rice is the most Al-tolerant among the crop species, with Al detoxification being managed by both mechanisms. Recently, we focused on ASR (Abscisic acid, Stress and Ripening) gene expression analyses and observed increased ASR5 transcript levels in roots and shoots in response to Al. In addition, ASR5 RNAi knock down plants presented an Al-sensitive phenotype. A proteomic approach showed that ASR5 silencing affected several proteins related to photosynthesis in RNAi rice shoots. Furthermore, an ASR5-GFP fusion in rice protoplasts revealed for the first time a chloroplast localization of this protein. Because it is well known that Al induces photosynthetic dysfunction, here we discuss the hypothesis that ASR5 might be sequestered in the chloroplasts as an inactive transcription factor that could be released to the nucleus in response to Al to regulate genes related to photosynthesis.
Keywords: :
According to the FAO, aluminum (Al) toxicity is the most serious problem in the soil constraints on agriculture, second to erosion hazards.Citation1 Al is a very abundant metal in the earth’s crust, but it becomes a problem under acidic conditions under which Al becomes soluble and is taken up by plants. Plants have developed strategies to cope with Al via internal and external detoxification systems. The release of organic acids that form complexes and prevent Al entry into cells has been well documented.Citation2-Citation4 However, some plants have managed to mediate Al by chelation and storage in the vacuoles.Citation5,Citation6 Among crops, rice is the most Al-tolerant species due to its capacity to cope with Al using both mechanisms.Citation7,Citation8 Although rice can exclude Al from the roots, a portion of the Al can enter the cells and be transported to the shoots.Citation10
In recent years, different studies aiming to decipher Al-tolerance in rice permitted the identification of a few genes,Citation9 and our previous work has added a new association between ASR genes (Abscisic acid, Stress and Ripening) and Al tolerance.Citation11 The ASR genes are also involved in many abiotic and biotic stresses (for a review, see Citation12). The exact function, however, remains enigmatic as the possible roles of the ASR genes cannot be deduced by sequence homology with other known proteins.Citation12 Most of the ASR proteins reported to date are located in the nucleus and possess DNA-binding activities. ASR from grape, for example, was able to bind to the promoter of a hexose transporter gene and regulate its expression.Citation13 Selex-binding experiments showed that ASR1 from tomato exhibit DNA sequence-preferential binding.Citation14 Furthermore, ASR1 from tomato was able to compete for the transcription factor ABI4 binding motif when overexpressed in Arabidopsis.Citation15
We have previously shown that ASR5 transcripts are increased in response to Al in roots and shoots.Citation11 ASR5_RNAi plants presented a high Al sensitivity and a trichome-less phenotype. In addition, a proteomic approach revealed several proteins of which the expression was affected in shoots due to the silencing of ASR5. Furthermore, we confirmed the nuclear localization of ASR5 using rice calli transformed with an ASR5_GFP fusion construct. However, when this construct was transformed in protoplasts, ASR5 was accumulated in the chloroplast precursors (etioplastids) of rice leaves protoplasts grown under dark conditions () and chloroplasts () of rice leaves protoplasts grown under light conditions. GFP alone was used as positive control (). This new and intriguing localization for ASR5 led us to speculate that ASR5 plays a role in this organelle, most likely linking the signaling between the chloroplasts and nucleus and regulating the expression of chloroplast proteins. Al can enter rice chloroplastsCitation16 and decrease the photosynthetic ratio.Citation17,Citation18 The ASR5 silencing affected the expression of at least 41 proteins in rice leaves.Citation11 A total of 19 out of these 41 proteins contains the chloroplast transfer peptide (cTP) signal, as predicted by sequence analysesCitation19,Citation20 and, were found to be differentially expressed in our RNAi line. The identified proteins are involved in such processes as photosynthesis, electron transport and stress responses (). The overexpression of ASR5 (named ASR1 in the cited report) in rice, increased the tolerance to cold, and the plants presented approximately 2-fold higher Fv/Fm values for the photosynthetic efficiency when compared with non-transformed plants.Citation21 Out of the 19 proteins with a cTP signal found in our plants, 14 were decreased in the ASR5_RNAi line and might be helpful to explain the Al sensitivity of the RNAi plant. These genes may be regulated by ASR5 in the nucleus because most of the proteins with cTP signals are encoded in the nuclear genome, translated in the cytosol and post-translationally imported into the chloroplasts.Citation22
Figure 1. Rice protoplast transformation with ASR5-GFP fusion. From upper to lower – The ref fluorescence of chlorophyll, ASR5-YFP fluorescence and merged images using a confocal laser scanning microscope. (a) Protoplast from rice leaves grown under dark conditions. (b) Protoplast from rice leaves grown under light conditions. (c) Vector with GFP only used as positive control. No green fluorescence was detected in negative control (data not shown). Bar = 10µM.
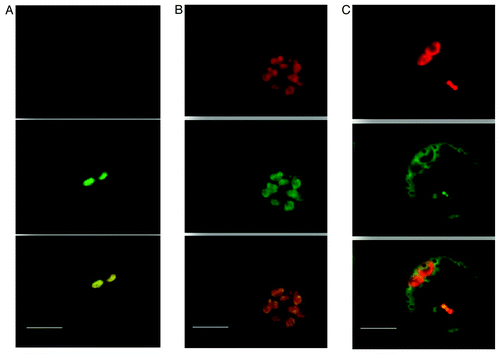
Table 1. Proteins differentially expressed in RNAi_ASR rice plants
Our hypothesis is that ASR5 is stored inside the chloroplast until specific conditions (such as Al stress) require their activity in the nucleus; a scenario proposed for certain other dual-target transcription factors.Citation23 For example, Whirly1 is localized in both the nucleus and plastids of the same cells in barley leaves.Citation24
Curiously, the ASR5 N-terminal region lacks the expected cTP sequence required to target this protein to the chloroplast. Notwithstanding, in Arabidopsis chloroplast preparations, a total of 604 proteins encoded in the nuclear genome were found, and surprisingly, only 376 were predicted to have the cTP when analyzed by a chloroplast-targeting prediction tool.Citation25 Proteins with cTP signals are targeted to the chloroplast surface and imported across the double-membrane envelope by translocons in the outer and inner envelope membranes, termed TOC and TIC, respectively.Citation26 However, increasing evidence shows that some proteins can enter the chloroplast by a TOC-TIC independent pathway.Citation27-Citation29 The transcriptional responses to abrupt environmental changes often have to occur rapidly and might require a release of pre-produced inactive transcription factors. A controlled sequestering of proteins at intracellular membranes seems to be an established way of controlling gene expression and intracellular communication.Citation23 Furthermore, a potential myristoylation site in rice ASR5 sequence is conserved among several ASR proteins at the same location when aligned.Citation30 The myristoyl residue may bind to a hydrophobic pocket and thus confer structural stability to a protein.Citation31 The most obvious function of the myristoyl moieties is to mediate membrane bindingCitation32 and is frequently found in proteins shuttling across membranes, where the myristoyl residue inserts into the inner lipid layer of the plasma membrane and facilitates membrane interaction.Citation30
In tobacco, Al accumulated in the chloroplasts after entering the cells, reacted with or replaced the non-heme iron between the QA and QB binding sites and blocked PSII electron transport, resulting in PSII photochemical damage and the inhibition of photosynthesis.Citation33 Thus, Al could be the signal for ASR5 release from the chloroplasts to regulate photosynthesis related gene expression and cope with Al toxicity in rice shoots.
Materials and Methods
The complete sequence of ASR5 was amplified using cDNA library of rice Japonica cultivar, sequenced and cloned fusionated with YFP coding sequence at its N-terminus into the Gateway vector pART7-YFP.Citation34 Protoplast isolation was performed according to Chen et al. (2006)Citation35 and protoplast transformation according to Tao et al. (2002).Citation36 Transformed rice protoplasts grown under dark and under light conditions were incubated in the dark for 24–48 h at 27°C prior to imaging. Fluorescence microscopy was performed with an Olympus FluoView 1000 confocal laser-scanning microscope equipped with a set of filters capable of distinguishing green and yellow fluorescent proteins (EGFP and EYFP, respectively) and plastid autofluorescence. Images were captured with a high-sensitivity photomultiplier tube detector. A vector containing only GFP was used as positive control and an empty vector was used as a negative control. The experiment was performed in biological triplicate.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
References
- Bot AJ, Nachtergaele FO, Young A. Land resource potential and constraints at regional and country level. F A O Land and Water Development Division, FAO, Rome 2000; Available at: http://www.fao.org/AG/agl/agll/terrastat.
- Sasaki T, Yamamoto Y, Ezaki B, Katsuhara M, Ahn SJ, Ryan PR, et al. A wheat gene encoding an aluminum-activated malate transporter. Plant J 2004; 37:645 - 53; http://dx.doi.org/10.1111/j.1365-313X.2003.01991.x; PMID: 14871306
- Ryan PR, Delhaize E, Jones D. Function and Mechanism of Organic Anion Exudation From Plant Roots. Annu Rev Plant Physiol Plant Mol Biol 2001; 52:527 - 60; http://dx.doi.org/10.1146/annurev.arplant.52.1.527; PMID: 11337408
- Ma JF, Furukawa J. Recent progress in the research of external Al detoxification in higher plants: a minireview. J Inorg Biochem 2003; 97:46 - 51; http://dx.doi.org/10.1016/S0162-0134(03)00245-9; PMID: 14507459
- Matsumoto H, Ma JF, Matsumoto H, Jian Zheng S, Feng Ma J. High aluminum resistance in buckwheat. I. Al-induced specific secretion of oxalic acid from root tips. Plant Physiol 1998; 117:745 - 51; http://dx.doi.org/10.1104/pp.117.3.745; PMID: 9662517
- Ma JF, Hiradate S, Nomoto K, Iwashita T, Matsumoto H. Internal Detoxification Mechanism of Al in Hydrangea (Identification of Al Form in the Leaves). Plant Physiol 1997; 113:1033 - 9; PMID: 12223659
- Huang CF, Yamaji N, Mitani N, Yano M, Nagamura Y, Ma JF. A bacterial-type ABC transporter is involved in aluminum tolerance in rice. Plant Cell 2009; 21:655 - 67; http://dx.doi.org/10.1105/tpc.108.064543; PMID: 19244140
- Yokosho K, Yamaji N, Ma JF. An Al-inducible MATE gene is involved in external detoxification of Al in rice. Plant J 2011; 68:1061 - 9; http://dx.doi.org/10.1111/j.1365-313X.2011.04757.x; PMID: 21880027
- Delhaize E, Ma JF, Ryan PR. Transcriptional regulation of aluminium tolerance genes. Trends Plant Sci 2012; 17:341 - 8; http://dx.doi.org/10.1016/j.tplants.2012.02.008; PMID: 22459757
- Meriga B, Reddy BK, Rao KR, Reddy LA, Kishor PBK. Aluminium-induced production of oxygen radicals, lipid peroxidation and DNA damage in seedlings of rice (Oryza sativa). J Plant Physiol 2004;161:63-8; PMID: 15002665; DOI: 0176-1617/04/161/01-63.
- Arenhart RA, Lima JC, Pedron M, Carvalho FEL, da Silveira JAG, Rosa SB, et al. Involvement of ASR genes in aluminum tolerance mechanisms in rice. Plant Cell Environ 2012; 1; http://dx.doi.org/10.1111/j.1365-3040.2012.02553.x
- Virlouvet L, Jacquemot MP, Gerentes D, Corti H, Bouton S, Gilard F, et al. The ZmASR1 protein influences branched-chain amino acid biosynthesis and maintains kernel yield in maize under water-limited conditions. Plant Physiol 2011; 157:917 - 36; http://dx.doi.org/10.1104/pp.111.176818; PMID: 21852416
- Çakir B, Agasse A, Gaillard C, Saumonneau A, Delrot S, Atanassova R. A grape ASR protein involved in sugar and abscisic acid signaling. Plant Cell 2003; 15:2165 - 80; http://dx.doi.org/10.1105/tpc.013854; PMID: 12953118
- Kalifa Y, Gilad A, Konrad Z, Zaccai M, Scolnik PA, Bar-Zvi D. The water- and salt-stress regulated Asr1 gene encodes a zinc-dependent DNA-binding protein. Biochem J 2004; •••:1 - 26; http://dx.doi.org/10.1042/BJ20031800; PMID: 15101820
- Shkolnik D, Bar-Zvi D. Tomato ASR1 abrogates the response to abscisic acid and glucose in Arabidopsis by competing with ABI4 for DNA binding. Plant Biotechnol J 2008; 6:368 - 78; http://dx.doi.org/10.1111/j.1467-7652.2008.00328.x; PMID: 18363631
- Jinqi L, Houtian L. Effect of aluminum on NAD kinase activity in chloroplast fraction from leaves of rice seedlings. J Environ Sci (China) 1995; 7:24 - 30
- Ali B, Hasan SA, Hayat S, Hayat Q, Yadav S, Fariduddin Q, et al. A role for brassinosteroids in the amelioration of aluminium stress through antioxidant system in mung bean (Vigna radiata L. Wilczek). Environ Exp Bot 2008; 62:153 - 9; http://dx.doi.org/10.1016/j.envexpbot.2007.07.014
- Zhang WH, Ryan PR, Tyerman SD. Malate-permeable channels and cation channels activated by aluminum in the apical cells of wheat roots. Plant Physiol 2001; 125:1459 - 72; http://dx.doi.org/10.1104/pp.125.3.1459; PMID: 11244125
- Emanuelsson O, Nielsen H, von Heijne G. ChloroP, a neural network-based method for predicting chloroplast transit peptides and their cleavage sites. Protein Sci 1999; 8:978 - 84; http://dx.doi.org/10.1110/ps.8.5.978; PMID: 10338008
- Schein AI, Kissinger JC, Ungar LH. Chloroplast transit peptide prediction: a peek inside the black box. Nucleic Acids Res 2001; 29:E82; http://dx.doi.org/10.1093/nar/29.16.e82; PMID: 11504890
- Kim S-J, Lee SC, Hong SK, An K, An G, Kim SR. Ectopic expression of a cold-responsive OsAsr1 cDNA gives enhanced cold tolerance in transgenic rice plants. Mol Cells 2009; 27:449 - 58; http://dx.doi.org/10.1007/s10059-009-0055-6; PMID: 19390826
- Soll J, Tien R. Protein translocation into and across the chloroplastic envelope membranes. Plant Mol Biol 1998; 38:191 - 207; http://dx.doi.org/10.1023/A:1006034020192; PMID: 9738967
- Krause K, Krupinska K. Nuclear regulators with a second home in organelles. Trends Plant Sci 2009; 14:194 - 9; http://dx.doi.org/10.1016/j.tplants.2009.01.005; PMID: 19285907
- Grabowski E, Miao Y, Mulisch M, Krupinska K. Single-stranded DNA-binding protein Whirly1 in barley leaves is located in plastids and the nucleus of the same cell. Plant Physiol 2008; 147:1800 - 4; http://dx.doi.org/10.1104/pp.108.122796; PMID: 18678751
- Kleffmann T, Russenberger D, von Zychlinski A, Christopher W, Sjölander K, Gruissem W, et al. The Arabidopsis thaliana chloroplast proteome reveals pathway abundance and novel protein functions. Curr Biol 2004; 14:354 - 62; http://dx.doi.org/10.1016/j.cub.2004.02.039; PMID: 15028209
- Bédard J, Jarvis P. Recognition and envelope translocation of chloroplast preproteins. J Exp Bot 2005; 56:2287 - 320; http://dx.doi.org/10.1093/jxb/eri243; PMID: 16087701
- Nada A, Soll J. Inner envelope protein 32 is imported into chloroplasts by a novel pathway. J Cell Sci 2004; 117:3975 - 82; http://dx.doi.org/10.1242/jcs.01265; PMID: 15286175
- Reinbothe S, Mache R, Reinbothe C. A second, substrate-dependent site of protein import into chloroplasts. Proc Natl Acad Sci U S A 2000; 97:9795 - 800; http://dx.doi.org/10.1073/pnas.160242597; PMID: 10920193
- Kim C, Ham H, Apel K. Multiplicity of different cell- and organ-specific import routes for the NADPH-protochlorophyllide oxidoreductases A and B in plastids of Arabidopsis seedlings. Plant J 2005; 42:329 - 40; http://dx.doi.org/10.1111/j.1365-313X.2005.02374.x; PMID: 15842619
- Vaidyanathan R, Kuruvilla S, Thomas G. Characterization and expression pattern of an abscisic acid and osmotic stress responsive gene from rice. Plant Sci 1999; 140:21 - 30; http://dx.doi.org/10.1016/S0168-9452(98)00194-0
- Zheng J, Knighton DR, Xuong NH, Taylor SS, Sowadski JM, Ten Eyck LF. Crystal structures of the myristylated catalytic subunit of cAMP-dependent protein kinase reveal open and closed conformations. Protein Sci 1993; 2:1559 - 73; http://dx.doi.org/10.1002/pro.5560021003; PMID: 8251932
- Vergères G, Manenti S, Weber T, Stürzinger C. The myristoyl moiety of myristoylated alanine-rich C kinase substrate (MARCKS) and MARCKS-related protein is embedded in the membrane. J Biol Chem 1995; 270:19879 - 87; http://dx.doi.org/10.1074/jbc.270.34.19879; PMID: 7650001
- Li Z, Xing F, Xing D. Characterization of target site of aluminum phytotoxicity in photosynthetic electron transport by fluorescence techniques in tobacco leaves. Plant Cell Physiol 2012; 53:1295 - 309; http://dx.doi.org/10.1093/pcp/pcs076; PMID: 22611177
- Galván-Ampudia CS, Offringa R. Plant evolution: AGC kinases tell the auxin tale. Trends Plant Sci 2007; 12:541 - 7; http://dx.doi.org/10.1016/j.tplants.2007.10.004; PMID: 18024140
- Chen SB, Tao LZ, Zeng LR, Vega-Sanchez ME, Umemura K, Wang GL. A highly efficient transient protoplast system for analyzing defence gene expression and protein-protein interactions in rice. Mol Plant Pathol 2006; 7:417 - 27; http://dx.doi.org/10.1111/j.1364-3703.2006.00346.x; PMID: 20507457
- Tao LZ, Cheung AY, Wu HM. Plant rac-like gtpases are activated by auxin and mediate auxin responsive gene expression. Plant Cell 2002; 14: 2745-2760; DOI: 10.1105/tpc.006320.