Abstract
In eukaryotes, nonsense-mediated mRNA decay (NMD) targets aberrant and selected non-aberrant mRNAs for destruction. A recent screen for mRNAs showing increased abundance in Arabidopsis NMD-deficient mutants revealed that most are associated with the salicylic acid (SA)-mediated defense pathway. mRNAs with conserved peptide upstream open reading frames (CpuORFs or CuORFs) are hugely overrepresented among the smaller class of NMD-regulated transcripts not associated with SA. Here we show that the common phenotypes observed in Arabidopsis NMD mutants are SA-dependent, whereas the upregulation of CpuORF-containing transcripts in NMD mutants is independent of SA. We speculate that CpuORFs could allow the conditional targeting of mRNAs for destruction using the NMD pathway.
In addition to its role in the destruction of aberrant mRNAs, nonsense-mediated mRNA decay (NMD) modulates gene expression by degrading endogenous transcripts bearing specific target features.Citation1-Citation6 Despite the presence of an NMD mechanism in all eukaryotes, few mRNAs have been identified as conserved NMD targets across evolution.Citation4,Citation5 The features that target a transcript to NMD are also not fully defined. In plants some, but not all, long 3′UTRs, 3′ UTR-located introns and upstream open reading frames (uORFs) result in NMD.Citation7-Citation12 In order to understand the role of NMD in plant gene regulation it is important to identify the transcripts that are targeted by NMD, the features that subject them to NMD and the effect of impairing NMD on growth and development.
Microarrays were used to identify 206 ‘common NMD transcripts’ that are coordinately upregulated in three Arabidopsis NMD mutants (upf1–5, upf3–1 and smg7–1).Citation13 These transcripts represent both direct and indirect NMD targets. Meta-analyses reveal that most of these transcripts (183) respond to pathogens in an SA dependent manner, whereas only 23 do not respond to pathogens or exogenously applied SA.Citation13 Furthermore, Arabidopsis NMD mutants are partially resistant to Pseudomonas syringae pathovar tomato DC3000 (DC3000) and accumulate more SA than their wild-type cohorts in the presence or absence of pathogen.Citation13 These findings suggest that one role for NMD in plants could be to facilitate a rapid pathogen response by changing the stability of pathogen responsive transcripts.
NMD mutant phenotypes in Arabidopsis
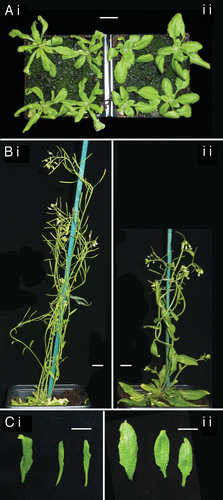
Plants deficient in different NMD effectors display similar phenotypes.Citation8,Citation9,Citation14,Citation15 The thin, twisted leaves of NMD mutants resemble SA accumulating mutants,Citation16 suggesting that some aspects of the NMD mutant phenotype may be caused by the constitutive pathogen response. To disentangle the effects of SA and NMD, NMD mutants were studied in an SA-deficient background. sid2–1 mutants are incapable of accumulating SA in response to pathogens.Citation17 The NMD impaired upf1–5 mutant exhibits the typical NMD phenotype of thin leaves with twisted petioles.Citation9 However, upf1–5 sid2–1 double mutants appear wild-type, with wider, flatter leaves (). This suggests that the distinctive leaf-shape phenotype of Arabidopsis NMD mutants is caused by accumulation of SA, consistent with a previously published finding that disruption of PAD4, which is upstream of SID2, also supresses the NMD mutant phenotype.Citation14
Figure 1. Phenotypes of NMD mutant plants with and without a functional ICS1 allele. All plants shown are homozygous for the mutant upf1–5 allele and therefore impaired in NMD. (i) Wild-type ICS1. (ii) Homozygous for the sid2–1 mutant allele of ICS1. Plants are 3 weeks (A), and 6 weeks (B and C) old. The white scale bar indicates 1 cm.
CpuORFs as targets for NMD
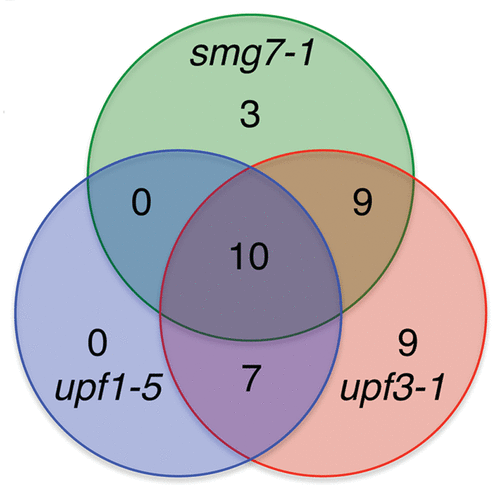
While it is likely that many of the 183 SA-responsive transcripts are indirectly affected by NMD, we previously suggested that the remaining 23 transcripts that do not respond to pathogens are enriched for direct NMD targets.Citation13 Eight of these 23 transcripts have CpuORFs; short open reading frames in the leader sequences that are conserved between Arabidopsis and rice.Citation13,Citation18 This is a striking overrepresentation given that only 44 Arabidopsis genes show this feature.Citation18 Comparisons with other species reveal that 82 Arabidopsis genes harbour CpuORFs.Citation18-Citation20 Of the 77 CpuORF-containing genes represented on the ATH1 GeneChip microarray, 49% are upregulated in at least one NMD mutant line (, Table S1). Enrichment for CpuORFs among NMD targets suggests that this feature is recognized by NMD.
Figure 2. Upregulation of CpuORF-containing transcripts in Arabidopsis plants deficient in NMD. The universe list for the Venn diagram is Arabidopsis loci that are represented on the ATH1 GeneChip microarray and that have CpuORFs that are conserved between Arabidopsis and another plant species (77 transcripts, Hayden and Jorgensen, 2007, Tran, Schiltz and Bauman, 2008 and Takahashi et al., 2012).Citation18-Citation20 Each circle represents transcripts that are upregulated at least 1.5-fold p < 0.05.
Many uORFs are not recognized by NMD and it is unclear why CpuORFs should differ from other uORFs in this respect. Although CpuORFs tend to be among the longest uORFs,Citation18,Citation21 length does not account for the differential representation of uORFs and CpuORFs in the list of NMD-regulated genes. CpuORFs are over-represented among the common NMD genes, even as a proportion of transcripts with a uORF exceeding 42 codons (p < 0.05). This suggests that the conserved sequence of the CpuORF may be important for recognition by NMD. In diverse organisms, nascent peptides of uORFs exert translational control of downstream ORFs by ribosome stalling at the uORF, sometimes in response to changing cellular conditions.Citation22-Citation24 Perhaps the short peptides encoded by the CpuORFs of NMD-sensitive transcripts expose the transcript to NMD by stalling the ribosome at the CpuORF, decreasing the efficiency of re-initiation of translation at the downstream ORF or making the transcript vulnerable to a change in cellular competence for translation reinitiation. Since recognition by NMD would depend on the translation of the CpuORF, this provides a mechanism by which individual transcripts could be targeted to, or protected from, NMD in a condition dependent manner. The is a precedent for this; in Saccharomyces cerevisiae, the CPA1 transcript harbours an uORF that encodes arginine attenuator peptide (AAP). In the presence of arginine, nascent AAP stalls ribosomes at the uORF targeting the transcript to NMD and downregulating CPA1 expression.Citation25
Publically available microarray data indicates that CpuORF-containing transcripts are mainly independent of SA and could therefore represent direct NMD targets. Meta-analyses across multiple microarray experiments show no association between CpuORFs and responses to SA. Furthermore, a specific search for transcripts that show increased abundance in smg7–1 mutants that are compromised in their ability to accumulate SA (smg7–1 pad4–1 vs pad4–1; PAD4 being a gene that mediates numerous responses to pathogens including SA accumulation via ICS1Citation26,Citation27) reveals 340 such transcripts (fold change > 1.5, p > 0.05). This list includes 16 CpuORF-containing transcripts, 8 of which are also commonly upregulated in the upf1–5, upf3–1 and smg7–1 NMD mutants. The PAD4-independence of the upregulation of these CpuORF transcripts supports the view that they are targeted directly by NMD, rather than responding indirectly via elevated SA.
Here we provide further evidence that CpuORFs are direct NMD targets in Arabidopsis. The high degree of conservation of CpuORFs suggests that they could also be direct NMD targets in other species. The presence of a CpuORF in the leaders of specific mRNAs could provide a mechanism whereby those mRNAs are conditionally moved into and out of the influence of NMD, raising the possibility that environmental and cellular conditions could act through CpuORFs to effectively alter the expression of the associated major transcripts.
Additional material
Download Zip (96.6 KB)Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
This work was supported by the BBSRC (BB/H00775X/1). The authors thank James Lloyd for critical comments and gratefully acknowledge Karel Riha for making the array data ‘NASCARRAYS-455’ publicly available. Array data for pad4–1 mutants was obtained from the NASCArrays repository.28
Related Research Data
References
- Maquat LE. Nonsense-mediated mRNA decay in mammals. J Cell Sci 2005; 118:1773 - 6; http://www.ncbi.nlm.nih.gov/pubmed/15860725 10.1242/jcs.01701; PMID: 15860725
- He F, Li XR, Spatrick P, Casillo R, Dong SY, Jacobson A. Genome-wide analysis of mRNAs regulated by the nonsense-mediated and 5′ to 3′ mRNA decay pathways in yeast. Molecular Cell 20003; 12:1439-52; PMID:14690598; http://www.ncbi.nlm.nih.gov/pubmed/14690598
- Mendell JT, Sharifi NA, Meyers JL, Martinez-Murillo F, Dietz HC. Nonsense surveillance regulates expression of diverse classes of mammalian transcripts and mutes genomic noise. Nat Genet 2004; 36:1073 - 8; http://www.ncbi.nlm.nih.gov/pubmed/15448691 10.1038/ng1429; PMID: 15448691
- Rehwinkel A, Letunic I, Raes J, Bork P, Izaurralde E. Nonsense-mediated mRNA decay factors act in concert to regulate common mRNA targets. RNA-a Publication of the RNA Society 2005; 11:1530-44; PMID:16199763; http://www.ncbi.nlm.nih.gov/pubmed/16199763
- Guan QN, Zheng W, Tang SJ, Liu XS, Zinkel RA, Tsui KW, et al. Impact of nonsense-mediated mRNA decay on the global expression profile of budding yeast. PLoS Genet 2006; 2:e203; http://www.ncbi.nlm.nih.gov/pubmed/17166056 10.1371/journal.pgen.0020203; PMID: 17166056
- Kurihara Y, Matsui A, Hanada K, Kawashima M, Ishida J, Morosawa T, et al. Genome-wide suppression of aberrant mRNA-like noncoding RNAs by NMD in Arabidopsis. Proc Natl Acad Sci U S A 2009; 106:2453 - 8; http://www.ncbi.nlm.nih.gov/pubmed/19181858 10.1073/pnas.0808902106; PMID: 19181858
- Hori K, Watanabe Y. UPF3 supresses aberrant spliced mRNA in Arabidopsis. Plant J 2006; 43:530 - 40; http://www.ncbi.nlm.nih.gov/pubmed/16098107 10.1111/j.1365-313X.2005.02473.x
- Yoine M, Nishii T, Nakamura K. Arabidopsis UPF1 RNA helicase for nonsense-mediated mRNA decay is involved in seed size control and is essential for growth. Plant Cell Physiol 2006; 47:572 - 80; http://www.ncbi.nlm.nih.gov/pubmed/16540482 10.1093/pcp/pcj035; PMID: 16540482
- Arciga-Reyes L, Wootton L, Kieffer M, Davies B. UPF1 is required for nonsense-mediated mRNA decay (NMD) and RNAi in Arabidopsis. Plant J 2006; 47:480 - 9; http://www.ncbi.nlm.nih.gov/pubmed/16813578 10.1111/j.1365-313X.2006.02802.x; PMID: 16813578
- Kerényi Z, Mérai Z, Hiripi L, Benkovics A, Gyula P, Lacomme C, et al. Inter-kingdom conservation of mechanism of nonsense-mediated mRNA decay. EMBO J 2008; 27:1585 - 95; http://www.ncbi.nlm.nih.gov/pubmed/18451801 10.1038/emboj.2008.88; PMID: 18451801
- Kertész S, Kerényi Z, Mérai Z, Bartos I, Pálfy T, Barta E, et al. Both introns and long 3′-UTRs operate as cis-acting elements to trigger nonsense-mediated decay in plants. Nucleic Acids Res 2006; 34:6147 - 57; http://www.ncbi.nlm.nih.gov/pubmed/17088291 10.1093/nar/gkl737; PMID: 17088291
- Nyikó T, Sonkoly B, Mérai Z, Benkovics AH, Silhavy D. Plant upstream ORFs can trigger nonsense-mediated mRNA decay in a size-dependent manner. Plant Mol Biol 2009; 71:367 - 78; http://www.ncbi.nlm.nih.gov/pubmed/19653106 10.1007/s11103-009-9528-4; PMID: 19653106
- Rayson S, Arciga-Reyes L, Wootton L, De Torres Zabala M, Truman W, Graham N, et al. A role for nonsense-mediated mRNA decay in plants: pathogen responses are induced in Arabidopsis thaliana NMD mutants. PLoS One 2012; 7:e31917; http://www.ncbi.nlm.nih.gov/pubmed/22384098 10.1371/journal.pone.0031917; PMID: 22384098
- Riehs-Kearnan N, Gloggnitzer J, Dekrout B, Jonak C, Riha K. Aberrant growth and lethality of Arabidopsis deficient in nonsense-mediated RNA decay factors is caused by autoimmune-like response. Nucleic Acids Research 2012; doi: 10.1093/nar/gks195; PMID:22379136; http://www.ncbi.nlm.nih.gov/pubmed/22379136
- Shi C, Baldwin IT, Wu J. Arabidopsis plants having defects in nonsense-mediated mRNA decay factors UPF1, UPF2, and UPF3 show photoperiod-dependent phenotypes in development and stress responses. J Integr Plant Biol 2012; 54:99 - 114; http://www.ncbi.nlm.nih.gov/pubmed/22353561 10.1111/j.1744-7909.2012.01093.x; PMID: 22353561
- Jeong H-J, Kim YJ, Kim SH, Kim Y-H, Lee I-J, Kim YK. Nonsense-mediated mRNA decay factors, UPF1 and UPF3, contribute to plant defense. Plant & Cell Physiology 2011; 52: 2147-56; PMID: 22025558; http://www.ncbi.nlm.nih.gov/pubmed/ 22025558.
- Wildermuth MC, Dewdney J, Wu G, Ausubel FM. Isochorismate synthase is required to synthesize salicylic acid for plant defence. Nature 2001; 414:562 - 5; http://www.ncbi.nlm.nih.gov/pubmed/11734859 10.1038/35107108; PMID: 11734859
- Hayden CA, Jorgensen RA. Identification of novel conserved peptide uORF homology groups in Arabidopsis and rice reveals ancient eukaryotic origin of select groups and preferential association with transcription factor-encoding genes. BMC Biol 2007; 5:32; http://www.ncbi.nlm.nih.gov/pubmed/17663791 10.1186/1741-7007-5-32; PMID: 17663791
- Tran MK, Schultz CJ, Baumann U. Conserved upstream open reading frames in higher plants. BMC Genomics 2008; 9:361; http://www.ncbi.nlm.nih.gov/pubmed/18667093 10.1186/1471-2164-9-361; PMID: 18667093
- Takahashi H, Takahashi A, Naito S, Onouchi H. BAIUCAS: a novel BLAST-based algorithm for the identification of upstream open reading frames with conserved amino acid sequences, and its application to the Arabidopsis thaliana genome. Bioinformatics 2012; PMID:22618534; http://www.ncbi.nlm.nih.gov/pubmed/22618534
- Kawaguchi R, Bailey-Serres J. mRNA sequence features that contribute to translational regulation in Arabidopsis.. Nucleic Acids Res 2005; 33:955 - 65; http://www.ncbi.nlm.nih.gov/pubmed/15716313 10.1093/nar/gki240; PMID: 15716313
- Cao J, Geballe AP. Inhibition of nascent-peptide release at translation termination. Mol Cell Biol 1996; 16:7109 - 14; http://www.ncbi.nlm.nih.gov/pubmed/8943366 PMID: 8943366
- Luo Z, Sachs MS. Role of an upstream open reading frame in mediating arginine-specific translational control in Neurospora crassa.. J Bacteriol 1996; 178:2172 - 7; http://www.ncbi.nlm.nih.gov/pubmed/8636015 PMID: 8636015
- Ruan H, Shantz LM, Pegg AE, Morris DR. The upstream open reading frame of the mRNA encoding S-adenosylmethionine decarboxylase is a polyamine-responsive translational control element. J Biol Chem 1996; 271:29576 - 82; http://www.ncbi.nlm.nih.gov/pubmed/8939886 10.1074/jbc.271.47.29576; PMID: 8939886
- Gaba A, Jacobson A, Sachs MS. Ribosome occupancy of the yeast CPA1 upstream open reading frame termination codon modulates nonsense-mediated mRNA decay. Mol Cell 2005; 20:449 - 60; http://www.ncbi.nlm.nih.gov/pubmed/16285926 10.1016/j.molcel.2005.09.019; PMID: 16285926
- Jirage D, Tootle TL, Reuber TL, Frost LN, Feys BJ, Parker JE, et al. Arabidopsis thaliana PAD4 encodes a lipase-like gene that is important for salicylic acid signaling. Proc Natl Acad Sci U S A 1999; 96:13583 - 8; http://www.ncbi.nlm.nih.gov/pubmed/10557364 10.1073/pnas.96.23.13583; PMID: 10557364
- Feys BJ, Moisan LJ, Newman M-A, Parker JE. Direct interaction between the Arabidopsis disease resistance signaling proteins, EDS1 and PAD4. EMBO J 2001; 20:5400 - 11; http://www.ncbi.nlm.nih.gov/pubmed/11574472 10.1093/emboj/20.19.5400; PMID: 11574472
- Craigon DJ, James N, Okyere J, Higgins J, Jotham J, May S. NASCArrays: a repository for microarray data generated by NASC’s transcriptomics service. Nucleic Acids Res 2004; 32:Database issue D575 - 7; http://www.ncbi.nlm.nih.gov/pubmed/14681484 10.1093/nar/gkh133; PMID: 14681484