Abstract
The Arabidopsis thaliana PHYTOALEXIN-DEFICIENT4 (PAD4) protein, which has homology to lipases, is required for phloem-based resistance against the green peach aphid (GPA; Myzus persicae Sülzer). PAD4 modulates antibiotic and antixenotic defenses against GPA. PAD4 in conjunction with its interacting partner ENHANCED DISEASE SUSCEPTIBILITY1 (EDS1) also functions in basal resistance to bacterial and oomycete pathogens by promoting salicylic acid (SA)-dependent and SA-independent defenses. By contrast, neither EDS1 nor SA is required for PAD4-controlled defense against GPA. Distinct molecular activities of PAD4 are involved in different aspects of Arabidopsis defense against GPA and pathogens. Histochemical analysis of plants containing a PAD4p:GUS chimera, which expresses the GUS reporter from the PAD4 promoter, indicated strong PAD4 promoter activity at the site of penetration of the vasculature by the insect stylet. GUS activity was also observed in non-vascular tissues of GPA-infested leaves, thus raising the possibility that a combination of distinct PAD4 activities in vascular and non-vascular tissues contribute to Arabidopsis defense against GPA.
Aphids (Hemiptera: Aphididae) are important pests of plants that use their slender stylets to feed on phloem sap.Citation1 The penetration path of aphid stylet is predominantly intercellular. However, occasionally, the aphid stylet pierces host cells. The green peach aphid (GPA), Myzus persicae (Sülzer), is one of the most damaging pests of many crop plants.Citation2 It has a host range covering more than hundred families of plants.Citation1 In addition, GPA is an important vector for more than hundred economically important plant viruses.Citation3 The interaction between Arabidopsis thaliana and GPA has provided an excellent model system to study the molecular mechanism of plant defense against aphids.Citation4 We have previously shown that expression of the Arabidopsis PHYTOALEXIN-DEFICIENT4 (PAD4) gene, which encodes a protein with homology to acyl hydrolases/lipases, is induced in response to GPA infestation.Citation5 PAD4 is an important modulator of antibiotic defenses that adversely impact GPA fecundity, and antixenotic defenses that impact plant choice by insect and feeding behavior.Citation5-Citation8 Electrical monitoring of insect feeding behavior confirmed a vital role for PAD4 in providing phloem-based resistance against GPA.Citation6 Recently, we established that serine at position 118 (S118) in PAD4, which corresponds to a key active site residue in other eukaryotic lipases, is critical for limiting GPA feeding from the sieve elements and for controlling aphid fecundity.Citation8 However, S118 was dispensable for deterring insect settling on plants.Citation8
The PAD4 Interacting Partner ENHANCED DISEASE SUSCEPTIBILITY1 is Not Required for Providing Antixenotic Defenses Against GPA
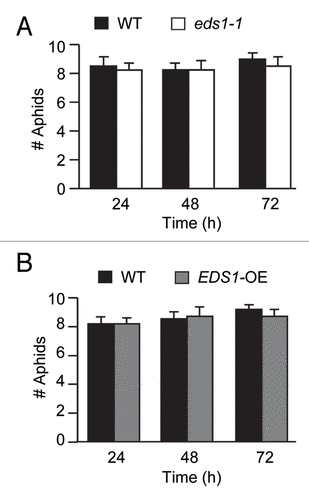
In Arabidopsis, the PAD4 protein along with its interacting partner, EDS1 (ENHANCED DISEASE SUSCEPTIBILITY1), which like PAD4 contains a region with homology to active site of acyl hydrolases/lipases, are also required for promoting defense against bacterial and oomycete pathogens.Citation9-Citation11 EDS1 is required for accumulation of PAD4 protein in pathogen-infected tissues.Citation10 The PAD4-EDS1 activity promotes accumulation of the defense signaling molecule salicylic acid (SA) in pathogen-infected plants. Like PAD4, EDS1 expression is also induced in response to GPA infestation.Citation6,Citation8,Citation10 However, PAD4’s involvement in Arabidopsis defense against GPA is molecularly distinct from its involvement in defense against pathogens.Citation8 By contrast to the larger population size of GPA on the pad4 mutant, aphid numbers were comparably low on the eds1 mutant and wild-type (WT) plants, thus indicating that EDS1 is not critical for the PAD4-dependent mechanism that controls GPA population size on Arabidopsis.Citation6 Furthermore, we have shown that redundancy between EDS1 and SID2 (SALICYLIC ACID INDUCTION DEFICIENT2), which is involved in the synthesis of SA, is not critical for controlling GPA population size on Arabidopsis. To determine if EDS1 has a role in antixenosis that impacts plant choice by GPA, insects were given a choice between the WT and the eds1 mutant plants. As shown in , the average number of GPAs that had settled on the WT was comparable to that on the eds1 mutant. Similarly, the insects showed no preference for the WT over the EDS1-OE transgenic plant in which EDS1 was constitutively overexpressed from the Cauliflower mosaic virus 35S gene promoter ().Citation8 These results further support our suggestion that PAD4 functions independent of EDS1, in mediating both antibiotic and antixenotic defenses against GPA.
Figure 1.EDS1 is not required for plant choice by insect. Choice assay: Twenty adult apterous (wingless) aphids were released at the center of each pot containing one WT accession Wassilewskija (Ws-0) and one eds1–1 plant (A), or one WT Ws-0 and one EDS1-OE plant (B), equidistant from the two plants. The numbers of released insects that had settled on plants of each genotype was determined at 24, 48 and 72 h post release. Values are the average of aphid counts on a minimum of eight plants of each genotype for each time point. Error bars represent SE. The means were separated using pooled χ2 test. There were no significant difference between insect numbers on the WT and the eds1–1, or WT and the EDS1-OE plants. The experiments in A and B were independently conducted two times.
PAD4 Promoter is Activated at Site of Penetration of Vascular Tissue by the Insect Stylet
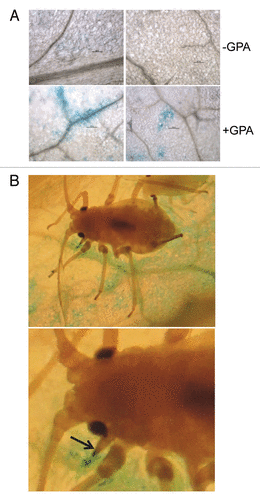
PAD4 activity is likely required in the vasculature to control GPA infestation. Alternatively, although not exclusively, PAD4 activity in another tissue type(s) may contribute to basal resistance against GPA. To determine if PAD4 expression is induced in the vascular tissues in response to GPA infestation, we generated transgenic Arabidopsis plants (accession Columbia) containing a chimeric reporter consisting of approximately 1.6 Kb of the PAD4 promoter (PAD4p) cloned upstream of the bacterial β-glucuronidase (GUS) coding sequence. GUS activity was monitored in situ with the synthetic substrate 5-bromo-4-chloro-3-indolyl glucuronide (X-Gluc) in un-infested and GPA-infested leaves of PAD4p:GUS plants. As shown in , by comparison to the un-infested leaves, which showed negligible GUS activity, blue coloration indicative of GUS activity was observed at several places around the vasculature in GPA-infested leaves. Strongest staining was observed at sites of penetration of vascular tissue by the insect stylet (). These results are supportive of our suggestion that PAD4 functions in the vascular tissues to control insect infestation.Citation6,Citation7,Citation12 Additional experiments are required to determine if expression of PAD4 in the vascular tissues is sufficient to complement one or more pad4 mutant defects in plant defense. Since GUS activity was also observed in clusters of non-vascular cells (), it is likely that PAD4 function in defense against GPA is required in other tissue types, as well. Our recent studies with a missense version of PAD4 in which serine at position 118 was replaced with alanine, suggested that distinct molecular activities of PAD4 contribute to different aspects of defense against GPA. Whether these distinct PAD4 activities are manifested in different tissue types, as alluded by the expression pattern of PAD4p:GUS in GPA-infested leaves (), remains to be determined.
Figure 2.PAD4p:GUS expression in un-infested and GPA-infested leaves. (A) Histochemical staining for GUS activity in un-infested (-GPA) (two upper panels) and GPA-infested (+GPA) (two lower panels) leaves, 48 h post release of insects. Left lower panel shows GUS activity around vasculature, and right lower panel shows GUS activity in cells distinct from vascular tissues. (B) Histochemical staining for GUS activity around vascular tissue near an insect (upper panel). Lower panel shows a close up view of GUS activity at site of penetration of plant tissue by the insect stylet. The aphid rostrum, the appendage to which the stylet is attached, is indicated by a black arrow.
Disclosure of Potential Conflict of Interest
The authors have no potential conflicts of interest
Acknowledgments
This work was funded by a grant (IOS-0919192) from the National Science Foundation. We thank Jane Parker for providing the eds1–1 and EDS1-OE seeds.
Note
GenBank IDs: PAD4 (At3g52430), EDS1 (At3g48090)
Related Research Data
References
- Blackman RL, Eastop VF. Aphids on the World’s Crops. An Indentification and Information Guide, Chichester, UK: John Wiley 2002; 2.
- Baker JR. Insect and related pests of flowers and foliage plants: Some important, common, and potential pests in the southeastern United States. Raleigh, NC, U.S.A: North Carolina Cooperative Extension Service Publication AG-136 1994.
- Kennedy JS, Day MF, Eastop VF. A Conspectus of Aphids as Vectors of Plant Viruses, London, UK: Commonwealth Institute of Entomology1962.
- Louis J, Singh V, Shah J. Arabidopsis thaliana-Aphid Interaction. Arabidopsis Book 2012; 10:e0159; PMID: 22666177
- Pegadaraju V, Knepper C, Reese JC, Shah J. Premature leaf senescence modulated by the Arabidopsis PAD4 gene is associated with defense against the phloem-feeding green peach aphid. Plant Physiol 2005; 139:1927 - 34; 10.1104/pp.105.070433; PMID: 16299172
- Pegadaraju V, Louis J, Singh V, Reese JC, Bautor J, Feys BJ, et al. Phloem-based resistance to green peach aphid is controlled by Arabidopsis PHYTOALEXIN DEFICIENT4 without its signaling partner ENHANCED DISEASE SUSCEPTIBILITY1.. Plant J 2007; 52:332 - 41; 10.1111/j.1365-313X.2007.03241.x; PMID: 17725549
- Louis J, Leung Q, Pegadaraju V, Reese J, Shah J. PAD4-dependent antibiosis contributes to the ssi2-conferred hyper-resistance to the green peach aphid. Mol Plant Microbe Interact 2010; 23:618 - 27; 10.1094/MPMI-23-5-0618; PMID: 20367470
- Louis J, Gobbato E, Mondal HA, Feys BJ, Parker JE, Shah J. Discrimination of Arabidopsis PAD4 activities in defense against green peach aphid and pathogens. Plant Physiol 2012; 158:1860 - 72; 10.1104/pp.112.193417; PMID: 22353573
- Falk A, Feys BJ, Frost LN, Jones JDG, Daniels MJ, Parker JE. EDS1, an essential component of R gene-mediated disease resistance in Arabidopsis has homology to eukaryotic lipases. Proc Natl Acad Sci U S A 1999; 96:3292 - 7; 10.1073/pnas.96.6.3292; PMID: 10077677
- Feys BJ, Wiermer M, Bhat RA, Moisan LJ, Medina-Escobar N, Neu C, et al. Arabidopsis SENESCENCE-ASSOCIATED GENE101 stabilizes and signals within an ENHANCED DISEASE SUSCEPTIBILITY1 complex in plant innate immunity. Plant Cell 2005; 17:2601 - 13; 10.1105/tpc.105.033910; PMID: 16040633
- Rietz S, Stamm A, Malonek S, Wagner S, Becker D, Medina-Escobar N, et al. Different roles of Enhanced Disease Susceptibility1 (EDS1) bound to and dissociated from PHYTOALEXIN DEFICIENT4 (PAD4) in Arabidopsis immunity. New Phytol 2011; 191:107 - 19; 10.1111/j.1469-8137.2011.03675.x; PMID: 21434927
- Louis J, Lorenc-Kukula K, Singh V, Reese J, Jander G, Shah J. Antibiosis against the green peach aphid requires the Arabidopsis thaliana MYZUS PERSICAE-INDUCED LIPASE1 gene. Plant J 2010; 64:800 - 11; 10.1111/j.1365-313X.2010.04378.x; PMID: 21105927