Abstract
Medicago truncatula and Sinorhizobium meliloti form a symbiotic association resulting in the formation of nitrogen-fixing nodules. In this organ, symbiotic cells contain large numbers of bacteroids. Remarkably, this chronic infection does not trigger visible defense reactions. Despite the importance of this phenomenon for potential transfer of the symbiotic capacity to non-legume plants, the molecular mechanisms underlying this tolerance are not understood.
We have characterized the dnf2 M. truncatula mutant blocked in the symbiotic process after bacterial infection of the symbiotic cells. Nodules formed by the mutant contain only few layers of infected cells. Furthermore, they exhibit defense-like reactions which clearly contrast with premature senescence frequently observed during inefficient symbioses. This atypical phenotype raises DNF2 as an exciting starting point to investigate the molecular basis of symbiotic repression of plant defenses.
Most legume species are able to form symbiotic interactions with soil bacteria referred to as rhizobia. During these associations, legumes differentiate a new organ dedicated to house bacteria: the nodule. Rhizobia often invade this organ using specialized symbiotic structures called infection threads (IT). These symbioses involve intimate contact between the two partners. The rhizobia reside in symbiotic nodule cells within organelle-like structures, separated from the host cell cytoplasm by a plant membrane. This structure is called symbiosome. In contrast to what is generally observed during microbial invasion, legumes do not elicit defense reactions during symbiosis despite that the bacterial population reaches massive densities in nodules. It has been suggested that the innate plant immune system might be suppressed in nodulesCitation1 and that this mechanisms is reversed as the nodules senesce.Citation2,Citation3 Not harming the rhizobium bacteria in the nodules is in the interest of the plants because rhizobia promote their growth by providing reduced nitrogen under the form of ammonium directly available for the plant. In exchange, bacteria receive carbon and energy supplies from the plants.
We recently characterized the DNF2 gene. dnf2 plant mutants develop nodules which are correctly invaded during the first stages of the symbiosis but in which the symbiotic process aborts once bacteria are released into the plant cell.Citation4 This arrest of the symbiotic process is accompanied with symbiosome degeneration, induction of a Pathogenesis Related 10 gene (PR10) and a senescence marker, necrosis due to phenolics accumulation as well as bacterial death.Citation4 These features are reminiscent of defense reactions and together clearly differ from early senescence which is commonly observed in non-fixing mutants (for instance nodules induced by a fix-minus rhizobium, ).
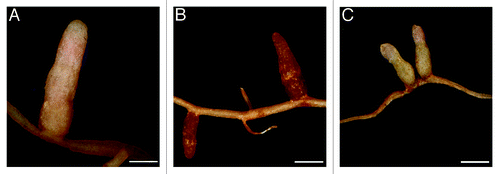
Figure 1.dnf2-4 displays typical features of defense reactions. Pictures show Wild Type (WT) R108 (A and C) and dnf2-4 (B) nodules 4 weeks after inoculation with Sinorhizobium meliloti 2011 (A and B) or its ΔnifH derivative (C). WT R108 shows big, pink nodules with WT strain and small, white nodules with Sm2011ΔnifH strain. In contrast to the fix- nodule shown in (C), dnf2 nodules show necrotic aspect. Scale bars: 1 cm.
Defense reactions in the frame of the rhizobium/legume symbiotic process have been discussed before.Citation5 Essentially all the previous studies related to defense reactions focused on early steps of the symbiotic process or on artificial cell culture system.Citation5
Here we highlight the interest to study the later steps of the symbiosis and particularly to understand how the plant cell can support the presence of so many bacteria into its cytoplasm without triggering defense reactions. Indeed, as the transfer of nitrogen-fixing symbiotic capacity to non-legume plants is an emerging goal,Citation6 studying the immune status of nodules should be one of the future priorities. In other words, it will be essential to understand how the plant cells and the thousands of bacteria within them stay alive and metabolically active during the symbiotic interaction.
The phenotype of the dnf2 mutantsCitation4 suggests an active mechanism responsible for the repression of plant defense within the symbiotic nodule cells. The requirement of such a specific mechanism taking place at the stage of cell invasion, by opposition to the one avoiding plant defenses in the IT, could be explained by the closer contact existing in symbiosomes between the bacterial cell envelope and the plant peribacteroid membrane. Indeed, the matrix that embedded bacteria in the ITCitation7 is drastically reduced in the symbiosomes leaving the bacterial cell surface and Pattern-Recognition Receptors (PRR) located on the plant plasma membrane in dangerous vicinity.
At least three processes can be envisage to prevent defense reactions in symbiotic cells and to maintain bacteria alive: (1) Bacteria are not detectable, (co)evolution might have led rhizobia to stop producing molecules eliciting plant defenses; (2) Detection systems are off; one can imagine that all PRRs, the receptors dedicated to detect the microbial presence, are not expressed in the symbiotic cells; (3) signal transduction might be compromised downstream of active PRRs either by the rhizobia or by the plant it-self via a nodule specific system. Examples of each of these possibilities are described in the literature: (1) Defense reactions are elicited in Lotus japonicus root systems in response to flg22 the active epitope of flagellin. In contrast, flagellin purified from Mesorhizobium loti does not elicit such defenses.Citation8 Modification of this Microbial Associated Molecular Pattern (MAMP) making it undetectable by the plant could be a general feature of rhizobia;Citation9 (2) The Pathogenesis Related Receptor FLS2 is drastically downregulated in L. japonicus nodules.Citation8 This might reflect a general downregulation of PRRs in the symbiotic organ (note that it seems futile to repress the gene required for flagellin repression if rhizobial flagellin cannot elicit defense reactions); (3) Bacterial exopolysaccharides are negatively charged polymers that chelate divalent cations and by this property they can inhibit Ca2+ influx required to develop MAMP triggered immunity.Citation10 It will be a priority for future research to determine whether the role of DNF2 fits in one of these scenarios. DNF2 encodes a PhosphoInositol-PhosphoLipaseC-XD-containing protein. It could function by masking or removing a bacterial surface MAMP (scenario 1), by interfering with PRR expression on symbiosome membranes (scenario 2) or by interfering with signal transduction (scenario 3).
To investigate further defense-like reactions development, studies should be conducted on both “normal” and altered rhizobia/legume interactions. Beyond the “normal” wild type situation, these studies should include: (1) mutant plants developing nodule necrosis, such as dnf2,Citation4 TRV36,Citation1 NF4608, NF1320Citation11 and latD/NIP;Citation12 (2) bacterial mutants triggering similar phenotype such as mutants altered in the general stress response regulator phyR/ecfGCitation13 as well as auxotrophic mutants;Citation14 (3) environmental conditions that trigger necrotic nodules such as boron deficiency which also results in induction of pathogenesis-related proteins from the PR10 family.Citation15 Characterization of the defense-like reactions developed in these different situations should be very informative and indicate if each situation requires a different signaling pathway. Are Rhizobia the eliciting agents? Are the defense-like reactions identical in all these situations? Do they induce priming of the defense reactions at distance? Are they primed by defenses related phytohormones? Under efficient symbiotic situation, exciting questions should also be addressed such as: is the immune system totally KO in the symbiotic cells and how can the plant do this? Are nodules responsive to priming by Induced Systemic Resistance or Systemic Aquired Resistance mechanisms?
The characterization of the dnf2 mutant and the corresponding DNF2 gene unravel a new aspect of the symbiotic interaction and the further unraveling of its mode of action will allow a better understanding of how plants can accommodate thousands of bacteria in a single cell without triggering defense reactions.
| Abbreviations: | ||
| IT | = | infection threads |
| PR10 | = | Pathogenesis Related 10 gene |
| PRR | = | Pattern-Recognition Receptors |
| MAMP | = | Microbial Associated Molecular Pattern |
Acknowledgments
We are grateful to Peter Mergaert for comments and suggestions on the manuscript. This work was supported by the Centre National de La Recherche Scientifique (CNRS) and the grant Agence Nationale de la Recherche (ANR) Blanc International SVSE 6.2010.1 (LEGUMICS) from the Agence National de la Recherche to P.R. M.B. was supported by a PhD fellowship from the French Ministry of Research. This work has benefited from the facilities and expertise of the Imagif Cell Biology Unit of the Gif campus (www.imagif.cnrs.fr) which is supported by the Conseil Général de l'Essonne.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Related Research Data
References
- Maunoury N, Redondo-Nieto M, Bourcy M, Van de Velde W, Alunni B, Laporte P, et al. Differentiation of symbiotic cells and endosymbionts in Medicago truncatula nodulation are coupled to two transcriptome-switches. PLoS ONE 2010; 5:e9519; http://dx.doi.org/10.1371/journal.pone.0009519; PMID: 20209049
- Puppo A, Groten K, Bastian F, Carzaniga R, Soussi M, Lucas MM, et al. Legume nodule senescence: roles for redox and hormone signalling in the orchestration of the natural aging process. New Phytol 2005; 165:683 - 701; http://dx.doi.org/10.1111/j.1469-8137.2004.01285.x; PMID: 15720680
- Mithöfer A. Suppression of plant defence in rhizobia-legume symbiosis. Trends Plant Sci 2002; 7:440 - 4; http://dx.doi.org/10.1016/S1360-1385(02)02336-1; PMID: 12399178
- Bourcy M, Brocard L, Pislariu CI, Cosson V, Mergaert P, Tadege M, et al. Medicago truncatula DNF2 is a PI-PLC-XD-containing protein required for bacteroid persistence and prevention of nodule early senescence and defense-like reactions. New Phytol 2013; 197:1250 - 61; PMID: 23278348
- Zamioudis C, Pieterse CM. Modulation of host immunity by beneficial microbes. Mol Plant Microbe Interact 2012; 25:139 - 50; http://dx.doi.org/10.1094/MPMI-06-11-0179; PMID: 21995763
- Charpentier M, Oldroyd G. How close are we to nitrogen-fixing cereals?. Curr Opin Plant Biol 2010; 13:556 - 64; http://dx.doi.org/10.1016/j.pbi.2010.08.003; PMID: 20817544
- Brewin NJ. Plant cell wall remodelling in the Rhizobiumum-Legume symbiosis. Crit Rev Plant Sci 2004; 23:293 - 316; http://dx.doi.org/10.1080/07352680490480734
- Lopez-Gomez M, Sandal N, Stougaard J, Boller T. Interplay of flg22-induced defence responses and nodulation in Lotus japonicus.. J Exp Bot 2012; 63:393 - 401; http://dx.doi.org/10.1093/jxb/err291; PMID: 21934117
- Felix G, Duran JD, Volko S, Boller T. Plants have a sensitive perception system for the most conserved domain of bacterial flagellin. Plant J 1999; 18:265 - 76; http://dx.doi.org/10.1046/j.1365-313X.1999.00265.x; PMID: 10377992
- Aslam SN, Newman M-A, Erbs G, Morrissey KL, Chinchilla D, Boller T, et al. Bacterial polysaccharides suppress induced innate immunity by calcium chelation. Curr Biol 2008; 18:1078 - 83; http://dx.doi.org/10.1016/j.cub.2008.06.061; PMID: 18639458
- Pislariu CI, Murray JD, Wen J, Cosson V, Muni RR, Wang M, et al. A Medicago truncatula tobacco retrotransposon insertion mutant collection with defects in nodule development and symbiotic nitrogen fixation. Plant Physiol 2012; 159:1686 - 99; http://dx.doi.org/10.1104/pp.112.197061; PMID: 22679222
- Veereshlingam H, Haynes JG, Penmetsa RV, Cook DR, Sherrier DJ, Dickstein R. nip, a symbiotic Medicago truncatula mutant that forms root nodules with aberrant infection threads and plant defense-like response. Plant Physiol 2004; 136:3692 - 702; http://dx.doi.org/10.1104/pp.104.049064; PMID: 15516506
- Gourion B, Sulser S, Frunzke J, Francez-Charlot A, Stiefel P, Pessi G, et al. The PhyR-sigma(EcfG) signalling cascade is involved in stress response and symbiotic efficiency in Bradyrhizobium japonicum.. Mol Microbiol 2009; 73:291 - 305; http://dx.doi.org/10.1111/j.1365-2958.2009.06769.x; PMID: 19555458
- Ferraioli S, Tatè R, Cermola M, Favre R, Iaccarino M, Patriarca EJ. Auxotrophic mutant strains of Rhizobium etli reveal new nodule development phenotypes. Mol Plant Microbe Interact 2002; 15:501 - 10; http://dx.doi.org/10.1094/MPMI.2002.15.5.501; PMID: 12036281
- Reguera M, Bonilla I, Bolaños L. Boron deficiency results in induction of pathogenesis-related proteins from the PR-10 family during the legume-rhizobia interaction. J Plant Physiol 2010; 167:625 - 32; http://dx.doi.org/10.1016/j.jplph.2009.11.017; PMID: 20138685