
Abstract
Pathogens use effector proteins to suppress host immunity and promote infection. However, plants can recognize specific effectors and mount an effector-triggered immune response that suppresses pathogen growth. The YopJ/HopZ family of type III secreted effector proteins is broadly distributed in bacterial pathogens of both animals and plants. These effectors can either suppress host immunity or elicit defense responses depending on the host genotype. In a recent report, we identified an Arabidopsis thaliana pseudokinase ZED1 that is required for the recognition of the Pseudomonas syringae HopZ1a effector. Here we discuss the role of ZED1 in HopZ1a recognition, and present models of effector recognition in plants. We draw parallels between HopZ1a and YopJ effector proteins, and between ZED1 and other immunity-related kinases that can be targeted by pathogen effectors.
Pathogens and their hosts are engaged in a dynamic molecular arms race where the growth and reproductive success of one usually comes at a cost to the other. Pseudomonas syringae is a Gram-negative bacterial pathogen that infects a wide range of plant species. It employs a type III secretion system to secrete and translocate type III secreted effector (T3SE) proteins into its hosts. T3SEs primarily function to suppress plant immunity.Citation1,Citation2,Citation3 Plants protect themselves from pathogens using 2 layers of immunity. The first layer of immunity relies on recognition of microbe-associated molecular patterns (MAMPs; e.g., flagellin), and leads to pattern recognition receptor (PRR)-triggered immunity (PTI).Citation4 The second layer of immunity results from recognition of specific T3SEs by nucleotide-binding leucine-rich repeat receptors (NLRs), and leads to effector-triggered immunity (ETI).Citation5 ETI typically includes a rapid form of programmed cell death called the hypersensitive response (HR) that restricts bacterial proliferation.Citation5,Citation6
Recognition of T3SEs can occur through direct interaction between a T3SE and an NLR, or indirectly where the T3SE and NLR both interact with an intermediate protein. The “guard” model was first proposed to account for indirect recognition of a T3SE, and postulates that the modification of a host virulence target (“the guardee”) by a T3SE is recognized by an NLR.Citation5,Citation7 An extension of the guard model is the “decoy” model, in which an effector target undergoes duplication and neo- or non-functionalization to evolve a protein that has no inherent function except to serve as a sentinel for effector activity.Citation8,Citation9 Importantly, the decoy model predicts that modification of the decoy by a T3SE can activate an NLR-mediated ETI response, but does not promote pathogen growth in the absence of recognition.
The YopJ/HopZ family of bacterial T3SEs is evolutionarily diverse and found in both mammalian and plant pathogens.Citation10 The P. syringae HopZ1a T3SE is recognized by the Arabidopsis NLR protein ZAR1.Citation11 We hypothesized a model of indirect recognition of HopZ1a by ZAR1 since the acetyltransferase activity of HopZ1a is required to activate ZAR1.Citation11,Citation12,Citation13 To identify other host components necessary for HopZ1a recognition, we performed a forward genetic screen and identified multiple mutants in one locus that were deficient for HopZ1a ETI. We named this locus ZED1 to reflect the hopZ ETI-deficient mutant phenotype.Citation14 Sequence analysis of ZED1 indicates that it is a pseudokinase that lacks the catalytic aspartic acid residue of the conserved HRD kinase motif.Citation15 We showed that ZED1 interacts directly with both HopZ1a and ZAR1, and that HopZ1a acetylates ZED1 at threonine 125 and threonine 177. Importantly, while ZED1 is required for HopZ1a-associated ETI, loss of ZED1 does not alter HopZ1a-associated virulence or PTI. Consequently, we propose that ZED1 acts as a decoy for HopZ1a, allowing the immune system to trap HopZ1a into the ZAR1 recognition complex. We hypothesize that once in this complex, ZED1 acetylation by HopZ1a contributes to ZAR1-mediated resistance ().
Figure 1. Recognition of YopJ, HopZ1a, and an unknown Xanthomonas T3SE. (A) Yersinia spp. injects YopJ into mammalian host cells. YopJ interferes with mitogen-activated protein kinase kinase (MAPKK) cascades via acetylation of MAPKK and IKKβ in the kinase binding site, thereby blocking downstream signaling and suppressing immune signaling. (B) Pseudomonas syringae injects HopZ1a into plant cells, where it is myristoylated and membrane-associated. HopZ1a acetylates ZED1, which triggers ZAR1-mediated immunity. (C) Xanthomonas spp. injects an unknown T3SE into plant cells that is recognized by ZRK1/RKS1 (a homolog of ZED1). The T3SE may modify ZRK1 to trigger immunity by an unknown NLR protein.
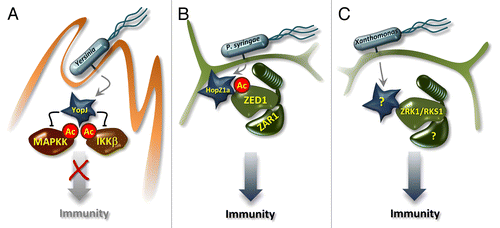
ZED1 is a member of a clade of closely related kinases we named ZED1-related kinases (ZRKs). Seven ZRKs and ZED1 are co-localized in a 14kbp region of Arabidopsis chromosome 3. At least some of the ZRK family members are predicted to be functional kinases, including ZRK10, which we validated experimentally. As discussed above, recognition of HopZ1a requires both ZED1 and ZAR1,Citation11,Citation14 but the absence of ZED1 does not eliminate HopZ1a’s ability to promote bacterial growth. This strongly suggests that ZED1 is not the virulence target of HopZ1a, but is instead a decoy.Citation11,Citation14 Consequently, we speculate that one or several other kinases, possibly in the ZED1/ZRK genomic cluster, are the true virulence targets of HopZ1a. We predict that HopZ1a acetylates these kinases, and that this acetylation promotes bacterial growth. This would be similar to the function of YopJ from Yersinia pestis, which acetylates and inactivates kinases to suppress their immunity-related functions ().Citation14,Citation15,Citation16
One of the ZRK family has recently been implicated in immunity against Xanthomonas campestris. Huard-Chauveau and colleagues independently identified resistance related kinase 1 (RKS1/ZRK1) as contributing to quantitative resistance against several strains of Xanthomonas campestris.Citation16 Like ZED1, ZRK1 appears to be a pseudokinase, although while ZED1 lacks a key catalytic residue, ZRK1 lacks residues involved in ATP binding.Citation14,Citation16 It is possible that ZRK1 triggers ETI when modified by an unidentified X. campestris T3SE ().
While pseudokinases such as ZED1 and ZRK1 are catalytically dead, they are not necessarily non-functional. Some pseudokinases lack catalytic sites but retain binding faces for protein-protein interaction.Citation17 Pseudokinases can also act as scaffolding proteins,Citation15 or bind ATP, and may also act as allosteric switches to regulate functional kinases.Citation18,Citation19 With respect to ZED1, it is unlikely that it only acts as a scaffold to help recruit ZAR1 or other kinases to a resistance signaling complex since loss-of-function point mutations occur in the predicted ATP binding pocket of ZED1, suggesting that an ATP-dependent ZED1 function may be involved in HopZ1a recognition.
Pseudokinases are emerging as crucial components of numerous biological processes, demonstrating that these catalytically inactive proteins are anything but non-functional.Citation15 The studies discussed here have extended the roles of pseudokinases to plant immunity. Given the inherent structural conservation of kinase active sites,Citation20 the deployment of pseudokinase decoys may prove to be irresistible traps for pathogen effectors that promiscuously target kinases to promote pathogen virulence. Further studies will show if plants commonly use this elegant mechanism to effectively turn the activity of a foreign molecule against itself.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
Work on HopZ function and recognition is supported by Natural Sciences and Engineering Research Council of Canada awards to Guttman DS and Desveaux D; a Canada Research Chair in Plant-Microbe Systems Biology (Desveaux D) or Comparative Genomics (Guttman DS); the Centre for the Analysis of Genome Evolution and Function (Desveaux D and Guttman DS); United States Department of Agriculture Agricultural Research Service 5335–21000–040–00D (Lewis JD).
Related Research Data
References
- Lewis JD, Guttman DS, Desveaux D. The targeting of plant cellular systems by injected type III effector proteins. Semin Cell Dev Biol 2009; 20:1055 - 63; http://dx.doi.org/10.1016/j.semcdb.2009.06.003; PMID: 19540926
- Deslandes L, Rivas S. Catch me if you can: bacterial effectors and plant targets. Trends Plant Sci 2012; 17:644 - 55; http://dx.doi.org/10.1016/j.tplants.2012.06.011; PMID: 22796464
- Block A, Li G, Fu ZQ, Alfano JR. Phytopathogen type III effector weaponry and their plant targets. Curr Opin Plant Biol 2008; 11:396 - 403; http://dx.doi.org/10.1016/j.pbi.2008.06.007; PMID: 18657470
- Segonzac C, Zipfel C. Activation of plant pattern-recognition receptors by bacteria. Curr Opin Microbiol 2011; 14:54 - 61; http://dx.doi.org/10.1016/j.mib.2010.12.005; PMID: 21215683
- Jones JDG, Dangl JL. The plant immune system. Nature 2006; 444:323 - 9; http://dx.doi.org/10.1038/nature05286; PMID: 17108957
- Heath MC. Hypersensitive response-related death. Plant Mol Biol 2000; 44:321 - 34; http://dx.doi.org/10.1023/A:1026592509060; PMID: 11199391
- Van der Hoorn RAL, De Wit PJ, Joosten MH. Balancing selection favors guarding resistance proteins. Trends Plant Sci 2002; 7:67 - 71; http://dx.doi.org/10.1016/S1360-1385(01)02188-4; PMID: 11832277
- van der Hoorn RAL, Kamoun S. From Guard to Decoy: a new model for perception of plant pathogen effectors. Plant Cell 2008; 20:2009 - 17; http://dx.doi.org/10.1105/tpc.108.060194; PMID: 18723576
- Block A, Alfano JR. Plant targets for Pseudomonas syringae type III effectors: virulence targets or guarded decoys?. Curr Opin Microbiol 2011; 14:39 - 46; http://dx.doi.org/10.1016/j.mib.2010.12.011; PMID: 21227738
- Lewis JD, Lee A, Ma W, Zhou H, Guttman DS, Desveaux D. The YopJ superfamily in plant-associated bacteria. Mol Plant Pathol 2011; 12:928 - 37; http://dx.doi.org/10.1111/j.1364-3703.2011.00719.x; PMID: 21726386
- Lewis JD, Wu R, Guttman DS, Desveaux D. Allele-specific virulence attenuation of the Pseudomonas syringae HopZ1a type III effector via the Arabidopsis ZAR1 resistance protein. PLoS Genet 2010; 6:e1000894; http://dx.doi.org/10.1371/journal.pgen.1000894; PMID: 20368970
- Lewis JD, Abada W, Ma W, Guttman DS, Desveaux D. The HopZ family of Pseudomonas syringae type III effectors require myristoylation for virulence and avirulence functions in Arabidopsis thaliana.. J Bacteriol 2008; 190:2880 - 91; http://dx.doi.org/10.1128/JB.01702-07; PMID: 18263728
- Lee AH-Y, Hurley B, Felsensteiner C, Yea C, Ckurshumova W, Bartetzko V, Wang PW, Quach V, Lewis JD, Liu YC, et al. A bacterial acetyltransferase destroys plant microtubule networks and blocks secretion. PLoS Pathog 2012; 8:e1002523; http://dx.doi.org/10.1371/journal.ppat.1002523; PMID: 22319451
- Lewis JD, Lee AH-Y, Hassan JA, Wan J, Hurley B, Jhingree JR, Wang PW, Lo T, Youn J-Y, Guttman DS, et al. The Arabidopsis ZED1 pseudokinase is required for ZAR1-mediated immunity induced by the Pseudomonas syringae type III effector HopZ1a. Proc Natl Acad Sci U S A 2013; 110:18722 - 7; http://dx.doi.org/10.1073/pnas.1315520110; PMID: 24170858
- Boudeau J, Miranda-Saavedra D, Barton GJ, Alessi DR. Emerging roles of pseudokinases. Trends Cell Biol 2006; 16:443 - 52; http://dx.doi.org/10.1016/j.tcb.2006.07.003; PMID: 16879967
- Huard-Chauveau C, Perchepied L, Debieu M, Rivas S, Kroj T, Kars I, Bergelson J, Roux F, Roby D. An atypical kinase under balancing selection confers broad-spectrum disease resistance in Arabidopsis.. PLoS Genet 2013; 9:e1003766; http://dx.doi.org/10.1371/journal.pgen.1003766; PMID: 24068949
- Scheeff ED, Eswaran J, Bunkoczi G, Knapp S, Manning G. Structure of the pseudokinase VRK3 reveals a degraded catalytic site, a highly conserved kinase fold, and a putative regulatory binding site. Structure 2009; 17:128 - 38; http://dx.doi.org/10.1016/j.str.2008.10.018; PMID: 19141289
- Taylor SS, Shaw A, Hu J, Meharena HS, Kornev A. Pseudokinases from a structural perspective. Biochem Soc Trans 2013; 41:981 - 6; http://dx.doi.org/10.1042/BST20130120; PMID: 23863167
- Iyer GH, Garrod S, Woods VL Jr., Taylor SS. Catalytic independent functions of a protein kinase as revealed by a kinase-dead mutant: study of the Lys72His mutant of cAMP-dependent kinase. J Mol Biol 2005; 351:1110 - 22; http://dx.doi.org/10.1016/j.jmb.2005.06.011; PMID: 16054648
- Hunter T. A thousand and one protein kinases. Cell 1987; 50:823 - 9; http://dx.doi.org/10.1016/0092-8674(87)90509-5; PMID: 3113737