
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.Abstract
We investigated electrical circuitry of the Venus flytrap, Mimosa pudica and Aloe vera. The goal was to discover if these plants might have a new electrical component—a resistor with memory. This element has attracted great interest recently and the researchers were looking for its presence in different systems. The analysis was based on cyclic current-voltage characteristic where the resistor with memory should manifest itself. We found that the electrostimulation of plants by bipolar sinusoidal or triangle periodic waves induces electrical responses in the Venus flytrap, Mimosa pudica and Aloe vera with fingerprints of memristors. Tetraethylammonium chloride, an inhibitor of voltage gated K+ channels, transforms a memristor to a resistor in plant tissue. Our results demonstrate that a voltage gated K+ channel in the excitable tissue of plants has properties of a memristor. This study can be a starting point for understanding mechanisms of memory, learning, circadian rhythms, and biological clocks.
Introduction
Electrical signals play important roles in electrophysiology of plants. These signals propagate along sophisticated electrical circuitry of plants consisting of many electrical components developed by nature. The standard electrical circuits comprise three basic elements: a resistor, a capacitor and an inductor. In addition to these elements, ChuaCitation1 postulated the existence of a new element, which has electrical memory. This is the fourth basic circuit element, a memristor or a resistor with memory. In spite of initial skeptical attitude to this idea, the first solid-state memristor was built already in 2008.Citation2 Memristors are memory circuit elements whose properties depend on the history and state of the system. Since then, researchers have been looking for the presence of memristors in different systems.Citation3-Citation12 They found a number of examples but up to now nobody was able to detect it in plants. Here we are going to demonstrate the existence of memristors in plants.
A memristor is a nano-scale memory device, which carries huge perspective technical applications. For example, they can replace flash memories and DRAMs in electronic appliances.Citation13 A memristor is a nonlinear element because its current-voltage characteristic is similar to that of a Lissajous pattern. No combination of nonlinear resistors, capacitors and inductors can reproduce this Lissajous behavior of the memristor. It is a fundamental 2-terminal described by the state dependent Ohm’s Law. Mathematically it can be described by an equation:
where ϕ and q denote the flux and charge, respectively.
Electrical circuits in plants operate over large distances.Citation14-Citation18 The activation of these circuits can lead to various physiological and biochemical responses.Citation19-Citation25 The cells of many biological organs generate electric potentials that can result in the flow of electric currentsCitation14,Citation26,Citation27 and propagation of action potentials. The Hodgkin-Huxley axon model describing action potentials is based on voltage gated channels. These channels can be identified as a potassium ion-channel memristor and a sodium ion-channel memristor.Citation3,Citation4 Since plants and animals have similar voltage gated K+ channels, it would be interesting to investigate the possible presence of memristors in plants. Chua et al.Citation3,Citation4 and Adhikari et al.Citation28 found that a memristor has three characteristic fingerprints: “When driven by a bipolar periodic signal the device must exhibit a pinched hysteresis loop in the voltage-current plane, assuming the response is periodic; starting from some critical frequency, the hysteresis lobe area should decrease monotonically as the excitation frequency increases; the pinched hysteresis loop should shrink to a single-valued function when the frequency tends to infinity.” The pinched hysteresis loop of memory elements, when subject to a periodic stimulus, can be self crossing (type I memristor) or not (type II memristor).
Plant tissues have different forms of memory such as sensory, short, and long-term memory.Citation15 Plants have a biological clock and circadian rhythms, which involve electrical elements of memory.Citation21,Citation24,Citation29,Citation30 We selected for this analysis the Venus flytrap and the Mimosa pudica as dicots, and the Aloe vera as a monocot and CAM plant. Aloe vera (L.) is a member of the Asphodelaceae (Liliaceae) family with crassulacean acid metabolism (CAM). In Aloe vera, stomata are open at night and closed during the day. CO2 acquired by Aloe vera at night is temporarily stored as malic and other organic acids, and is decarboxylated the following day to provide CO2 for fixation in the Benson-Calvin cycle behind closed stomata. The main goal of this article is to find out if plants comprise memristors.
Results
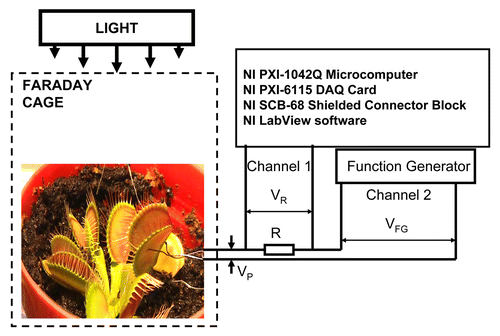
Experimental setup is shown in . Bipolar sinusoidal or triangle periodic waves with amplitude VFG were applied from a function generator. To measure electrical current we included in the circuit additional 47 kΩ resistor R, so that electrical current was found as I = VR/R. Potential difference, VP, between electrodes in plants is equal to VP = VFG – VR ().
Figure 1. Block diagram of the data acquisition and electrostimulation system.
The Venus flytrap is a carnivorous plant. The trap can be closed by mechanical stimulation of trigger hairs or by an electrical pulse between the midrib and a lobe of the upper leaf.Citation19
We recorded the current flowing through the plant and generated by a bipolar triangle wave with frequency of 0.001 Hz () for closed trap (panel A) and for an open trap (panel B). In both cases we obtained a pinched hysteresis loop in the voltage-current plane with one important difference. If the trap is closed, the plot displays a common pinched point with self-crossing between curves when I = 0 μA and VP = 0 V (). If the trap is open, the plot also displays a common pinched point but without self-crossing between curves with coordinates I = 1.5 μA and VP = 0.16 V (). At the first glance it might be not obvious that the curves in self-cross. So, to avoid the confusion we added the arrows to this panel. If one just follows these arrows, the crossing of the curves becomes immediately clear.
Figure 2. Dependencies of electrical current, I, in the Venus flytrap on VP induced by triangle voltage wave VFG from a function generator (A,B,C); Frequency of triangle voltage VFG scanning was 0.001 Hz (A,B) and 10 Hz (c); R = 47 kΩ. Position of Ag/AgCl electrodes in the Venus flytrap is shown. The trap was open in (B,C) or closed in (A). These results were reproduced 16 times.
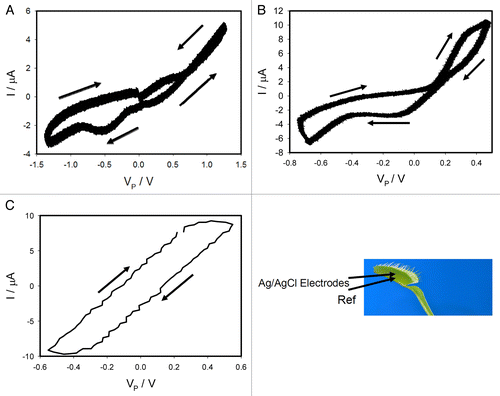
The behavior in can be described as “kiss-and-go”: the two branches touch each other but do not cross. The deviation of coordinates VP and I from zero value can be caused by the action potential propagation in the trap, which has amplitude of 0.16 V.Citation20,Citation31 Increasing of a triangle wave frequency to 1 kHz changes the shape of the line: it is still a loop but without a pinched point for both open () and closed traps. The branches of the loop go parallel to each other. So, the electrostimulation of the Venus flytrap by a periodic wave induces electrical responses in the Venus flytrap with fingerprints of a memristor.
Voltage gated ionic channels regulate generation and transduction of electrical signals. For their analysis there are very efficient tools - blockers of ionic channels.Citation29,Citation30 It was intriguing to investigate if these blockers would change characteristics or even the presence of memristors in plants. Tetraethylammonium chloride (TEACl) is known as a blocker of a voltage gated K+ channel.Citation30 We found that deposition of 20 μL of 10 mM TEACl on the midrib of the Venus flytrap decreased the amplitude of electrical current and the hysteresis shrank (). This can be caused by the increasing of resistance in the plant tissue. An inhibitor of voltage gated K+ channels tetraethylammonium chloride transforms a memristor to a simple resistor in plant tissue. Increasing of a triangle wave frequency to 10 Hz changes the shape of the line: it is still a loop but without a pinched point (). These results demonstrate that a voltage gated K+ channel in the excitable tissue of the Venus flytrap is an essential component of plant memristor.
Figure 3. Effects of voltage gated channel inhibitor TEACl deposited on the midrib of the Venus flytrap 5 h before electrical measurements. Frequency of triangle voltage VFG scanning was 0.001 Hz (A) and 10 Hz (B); R = 47 kΩ. Position of Ag/AgCl electrodes in the Venus flytrap is shown. The trap was open. These results were reproduced 16 times.
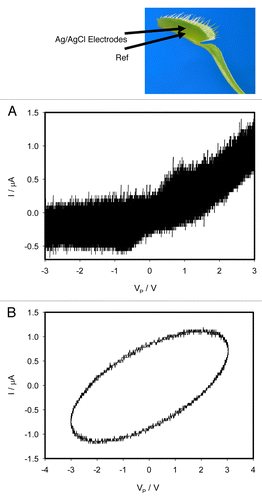
The Mimosa pudica is a nyctinastic plant that closes its’ leaves in the evening. The leaves open in the morning due to a circadian rhythm, which is regulated by a biological clock with a cycle of about 24 h. Leaf movement in the Mimosa pudica appear to be regulated by electrical signal transduction. Mechanics of these movements are hidden in the specialized organ - the pulvinus.Citation17,Citation25 We have found that the movements of the petiole, or pinnules, connected to pulvinus are accompanied by a change of the pulvinus morphing structures. This structure has rather peculiar electrical properties similar to the electrical synapse in the animal nerves.Citation25 Therefore, it was interesting to investigate electrical circuitry of this organ. shows dependencies of electrical current induced by bipolar sinusoidal wave with amplitude of ± 3.5V (a) or ± 4.5V (b) and frequency of 0.001 Hz, where platinum electrodes are inserted along the pulvinus of Mimosa pudica.
Figure 4. Dependencies of electrical current, I, on voltage VP applied along a pulvinus. Frequency of sinusoidal voltage scanning was 0.001 Hz. R = 47 kΩ. Position of Pt electrodes in the pulvinus of the Mimosa pudica is shown. Amplitude of applied voltage VFG from a function generator varies from -3.5 V to + 3.5 V (A) and from -4.5 V to + 4.5 V (B). These results were reproduced 27 times.
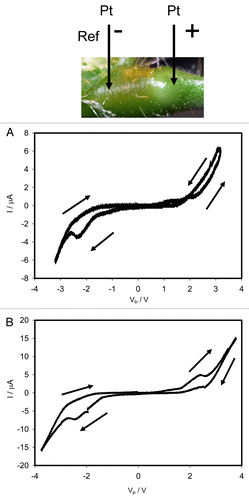
There is a pinched point in hysteresis loop at low frequency of sinusoidal wave in the voltage-current plane when I = 0 μA and VP = 0 V, which is a typical sign of a memristor. shows that there is a self-crossing between curves when amplitude of the applied sinusoidal voltage is between ± 3.5V. Increase of the amplitude of the sinusoidal voltage to 4.5 V changes the curve (): the plot displays a common pinched point without self-crossing. Increasing of a sinusoidal wave frequency to 1 kHz changes the shape of a hysteresis loop and a pinched point disappears. Similar dependencies of electrical current on voltage were observed in a stem of the Mimosa pudica. There is a pinched point but without self-crossing between curves when I = 0 μA and VP = 0 V.
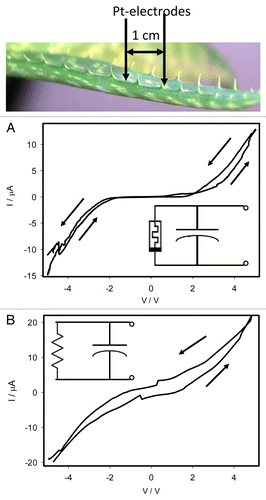
The Aloe vera (L.) is a member of the Asphodelaceae (Liliaceae) family with crassulacean acid metabolism (CAM). shows cyclic voltammetry in the leaf of Aloe vera. At a low frequency of scanning, there is a pinched point with self-crossing between curves when I = 0 μA and VP = 0 V, which is a typical fingerprint of a memristor (). There is also an additional pinched point at the potential of 2 V. Increasing the speed of scanning from 2 mV/s (2x10−4 Hz) to 500 mV/s (0.05 Hz) leads to the disappearance of pinched points in the hysteresis loop (). The simple possible equivalent electrical circuits are shown in inserts.
Figure 5. Cyclic voltammetry in a leaf of Aloe vera with a scanning rate of periodic triangle wave of 2 mV/s (A) and 500 mV/s (B). Position of Pt electrodes in the Aloe vera is shown. The simplest equivalent electrical circuits are shown in inserts. Electrodes were located along the leaf on the distance of 1 cm. These results were reproduced 16 times.
Discussion
In the Venus flytrap, Mimosa pudica, and Aloe vera plants we found the presence of resistors with memory. It is truly fascinating that even plants exhibit memristor fingerprints. When driven by a bipolar periodic sinusoidal or triangle signal plants exhibit a pinched hysteresis loop in the voltage-current plane (,,). Starting from some critical frequency, the hysteresis loop changes shape and a pinched hysteresis loop transforms to a non-pinched hysteresis (, ) as the excitation frequency increases. Adhikari et al.Citation28 have shown that there are two types of pinched hysteresis loops in memristors: a transversal type where two branches of the hysteresis loop are self-crossing and a non-transversal or tangential type where two branches of the hysteresis loop are tangent at the pinched point. Voltage gated channels can exhibit more than one self-intersection points.Citation5 The third fingerprint of a memristor discovered by ChuaCitation13 and Adhikari et al.Citation5: “the pinched hysteresis loop should shrink to a single-valued function when the frequency tends to infinity” is correct for ideal memristors. For a plant tissue, the pinched hysteresis loop transforms to a non-pinched hysteresis loop instead of a single line I = V/R at high frequencies of the applied voltage because the amplitude of electrical current hysteresis depends also on capacitance of a plant tissue and electrodes, frequency and direction of scanning.
shows simple equivalent electrical circuits for low frequency measurements with a memristor () and for high frequency measurements with a resistor (). The goal of this work is not to offer an exact memristor model of the plants, which would be subject of future research, but to point out for the first time, that even plants exhibit memristive characteristics. The physiology of plants must include memristors as essential model building blocks in electrical networks in plants. The memristor is an “ideal” circuit element and no real-world biodevices can be exactly mimicked by an ideal circuit model. There are different models of increasing accuracy that can be developed at the cost of increasing complexity.
Materials and Methods
Plants
The Dionaea muscipula Ellis (Venus flytrap) plants were purchased from Fly-Trap Farm Supply (North Carolina, USA) and grown in well drained peat moss in plastic pots at 22 °C with 12:12h light: dark photoperiod. The soil was treated with distilled water.
Fifty Aloe vera L plants were grown in clay pots. Aloe vera plants had 25 – 35 cm leaves.
The seeds of Mimosa pudica L. were soaked in warm water (30 °C) for 48 h. They were then grown in well drained peat moss at 21 °C with a 12:12 h light:dark photoperiod. After growing for two weeks, the seedlings were transplanted into pots and placed inside a plant growing chamber. The plants were watered every day. Two-three month old plants were used for the experiments.
Irradiance was 700–800 μmol photons m−2s−1. The humidity averaged 45 – 50%. All experiments were performed on healthy adult specimens.
Chemicals
Tetraethylammonium chloride (TEACl) was obtained from Fluka (New York).
Electrodes for extracellular measurements
Ag/AgCl electrodes were prepared in the dark from Teflon coated silver wires (A-M Systems, Inc., Sequim, WA, USA) with a diameter of 0.2 mm by electrolysis of 5 mm long silver wire tip without Teflon coating in a 0.1 M KCl aqueous solution. The response time of Ag/AgCl electrodes was less than 0.1 μs. We also used platinum electrodes instead of Ag/AgCl electrodes and acquired the same results. Platinum electrodes were prepared from Teflon coated platinum wires (A-M Systems, Inc..) with diameter of 0.076 mm. In all experiments we used identical (Pt or Ag/AgCl) electrodes as a measuring and as reference (Ref) electrodes.
Data acquisition
All measurements were conducted in the laboratory at constant room temperature of 22 oС inside a Faraday cage which was mounted on a vibration-stabilized table (). High speed data acquisition of low-pass filtered signals was performed using microcomputer NI-PXI-1042Q (National Instruments) with simultaneous multifunction I/O plug-in data acquisition board NI-PXI-6115 (National Instruments) interfaced through a NI SCB-68 shielded connector block to electrodes.
Plant electrostimulation
The function generator FG300 (Yokagawa, Japan) was interfaced to NI-PXI-1042Q microcomputer and used for electrostimulation of plants. Resistance between electrodes in plants in all our experiments was between 1.1 MΩ and 1.5 MΩ. We selected a resistor R = 47 kΩ for measuring of voltage, VR, for estimation of electrical current I.
Cyclic voltammetry
The SI1287 Advanced Electrochemical Interface (Solartron, Hampshire, England)) was connected via the GPIB to PC and used in two terminal configuration for cyclic voltammetry.
Statistics
All experimental results were reproduced 25 times on different plants. Software SigmaPlot 12 (Systat Software, Inc.) was used for statistical analysis of experimental data.
| Abbreviations: | ||
| C | = | capacitance |
| DAQ | = | data acquisition |
| PXI | = | PCI eXtensions for Instrumentation |
| I | = | electrical current |
| V | = | voltage between electrodes in plants, VFG, voltage of an function generator VR,voltage on resistor R |
| Vp | = | voltage between electrodes in plants |
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
This work was supported in part by the National Science Foundation (grant no. CBET-1064160) and in part by the U. S. Army Research Office (contract/grant no W911NF-11-1-0132). L. Chua's research is supported by AFOSR grant number FA9550-13-1-0136.
References
- Chua L. Memristor – The missing circuit element. IEE Transactions Circuit Theory 1971; 18:507 - 19; http://dx.doi.org/10.1109/TCT.1971.1083337
- Strukov DB, Snider GS, Stewart DR, Williams RS. The missing memristor found. Nature 2008; 453:80 - 3; http://dx.doi.org/10.1038/nature06932; PMID: 18451858
- Chua L, Sbitnev V, Kim H.. Hodgkin-Huxley axon is made of memristors. Internat J Bifurcation Chaos 2012; 22:1230011 - 48; http://dx.doi.org/10.1142/S021812741230011X
- Chua L, Sbitnev V, Kim H.. Neurons are poised near the edge of chaos. Internat J Bifurcation Chaos 2012; 22:1250098 - 49; http://dx.doi.org/10.1142/S0218127412500988
- Sah M, Kim H, Chua L. Brains are made of memristors. IEEE Circuits Systems 2014; 14 In press
- Katz E, MacVittie K. Electrochemical systems with memimpedance properties. J Phys Chem C 2013; 117:24943 - 7; http://dx.doi.org/10.1021/jp409257v
- Pershin YV, La Fontaine S, Di Ventra M. Memristive model of amoeba learning. Phys Rev E Stat Nonlin Soft Matter Phys 2009; 80:021926; http://dx.doi.org/10.1103/PhysRevE.80.021926; PMID: 19792170
- Chua L. Memristor, Hodgkin-Huxley, and edge of chaos. Nanotechnology 2013; 24:383001; http://dx.doi.org/10.1088/0957-4484/24/38/383001; PMID: 23999613
- Borghetti J, Snider GS, Kuekes PJ, Yang JJ, Stewart DR, Williams RS. ‘Memristive’ switches enable ‘stateful’ logic operations via material implication. Nature 2010; 464:873 - 6; http://dx.doi.org/10.1038/nature08940; PMID: 20376145
- Smerieri A, Berzina T, Erokhin V, Fontana MP. Polymeric electrochemical elements for adaptive networks: Pulse mode. J Appl Phys 2008; 104:114513 - 8; http://dx.doi.org/10.1063/1.3033399
- Jo SH, Chang T, Ebong I, Bhadviya BB, Mazumder P, Lu W. Nanoscale memristor device as synapse in neuromorphic systems. Nano Lett 2010; 10:1297 - 301; http://dx.doi.org/10.1021/nl904092h; PMID: 20192230
- Jacobsen T, Zachau-Christiansen B, Bay L, Jorgensen MJ. Hysteresis in the solid oxide fuel cell cathode reaction. . Electrochim Acta 2001; 46:1019 - 24; http://dx.doi.org/10.1016/S0013-4686(00)00689-7
- Chua L. Resistance switching memories are memristors. Appl Phys, A Mater Sci Process 2011; 102:765 - 83; http://dx.doi.org/10.1007/s00339-011-6264-9
- Volkov AG. Green Plants: Electrochemical Interfaces. J Electroanal Chem 2000; 483:150 - 156; http://dx.doi.org/10.1016/S0022-0728(99)00497-0
- Volkov AG, Carrell H, Baldwin A, Markin VS. Electrical memory in Venus flytrap. Bioelectrochemistry 2009; 75:142 - 7; http://dx.doi.org/10.1016/j.bioelechem.2009.03.005; PMID: 19356999
- Volkov AG, Carrell H, Markin VS. Biologically closed electrical circuits in venus flytrap. Plant Physiol 2009; 149:1661 - 7; http://dx.doi.org/10.1104/pp.108.134536; PMID: 19211696
- Volkov AG, Foster JC, Ashby TA, Walker RK, Johnson JA, Markin VS. Mimosa pudica: Electrical and mechanical stimulation of plant movements. Plant Cell Environ 2010; 33:163 - 73; http://dx.doi.org/10.1111/j.1365-3040.2009.02066.x; PMID: 19895396
- Volkov AG, Foster JC, Markin VS. Signal transduction in Mimosa pudica: biologically closed electrical circuits. Plant Cell Environ 2010; 33:816 - 27; PMID: 20040063
- Volkov AG, Adesina T, Jovanov E. Closing of venus flytrap by electrical stimulation of motor cells. Plant Signal Behav 2007; 2:139 - 45; http://dx.doi.org/10.4161/psb.2.3.4217; PMID: 19516982
- Volkov AG, Adesina T, Markin VS, Jovanov E. Kinetics and mechanism of Dionaea muscipula trap closing. Plant Physiol 2008; 146:694 - 702; http://dx.doi.org/10.1104/pp.107.108241; PMID: 18065564
- Volkov AG, Baker K, Foster JC, Clemmons J, Jovanov E, Markin VS. Circadian variations in biologically closed electrochemical circuits in Aloe vera and Mimosa pudica.. Bioelectrochemistry 2011; 81:39 - 45; http://dx.doi.org/10.1016/j.bioelechem.2011.01.004; PMID: 21334987
- Volkov AG, Foster JC, Jovanov E, Markin VS. Anisotropy and nonlinear properties of electrochemical circuits in leaves of Aloe vera L. Bioelectrochemistry 2011; 81:4 - 9; http://dx.doi.org/10.1016/j.bioelechem.2010.11.001; PMID: 21167797
- Volkov AG, Pinnock MR, Lowe DC, Gay MS, Markin VS. Complete hunting cycle of Dionaea muscipula: consecutive steps and their electrical properties. J Plant Physiol 2011; 168:109 - 20; http://dx.doi.org/10.1016/j.jplph.2010.06.007; PMID: 20667624
- Volkov AG, Wooten JD, Waite AJ, Brown CR, Markin VS. Circadian rhythms in electrical circuits of Clivia miniata.. J Plant Physiol 2011; 168:1753 - 60; http://dx.doi.org/10.1016/j.jplph.2011.03.012; PMID: 21546115
- Volkov AG, O’Neal L, Volkova MI, Markin VS. Morphing structures and signal transduction in Mimosa pudica L. induced by localized thermal stress. J Plant Physiol 2013; 170:1317 - 27; http://dx.doi.org/10.1016/j.jplph.2013.05.003; PMID: 23747058
- Volkov AG, ed. Plant Electrophysiology. Methods and Cell Electrophysiology. Berlin: Springer; 2012.
- Volkov AG, ed. Plant Electrophysiology. Signaling and Responses. Berlin: Springer; 2012.
- Adhikaru AP, Sah MP, Kim H, Chua L. The fingerprints of memristor. IEEE Trans Circuits Systems 2013; http://dx.doi.org/10.1109/TCSI.2013.2256171
- Markin VS, Volkov AG. Morphing structures in the Venus flytrap. In: Volkov AG, ed. Plant Electrophysiology. Signaling and Responses. Berlin: Springer; 2012: 1-31
- Volkov AG, Coopwood KJ, Markin VS. Inhibition of the Dionaea muscipula Ellis trap closure by ion and water channels blockers and uncouplers. Plant Sci 2008; 175:642 - 9; http://dx.doi.org/10.1016/j.plantsci.2008.06.016
- Escalante-Pérez M, Krol E, Stange A, Geiger D, Al-Rasheid KAS, Hause B, Neher E, Hedrich R. A special pair of phytohormones controls excitability, slow closure, and external stomach formation in the Venus flytrap. Proc Natl Acad Sci U S A 2011; 108:15492 - 7; http://dx.doi.org/10.1073/pnas.1112535108; PMID: 21896747