
Abstract
Plants have evolved sophisticated defense mechanisms to overcome their sessile nature. One remarkable strategy is the inter-plant communication mediated by volatile organic compounds (VOCs). Quantity and quality of plant VOCs are intricately regulated by biotic and abiotic stresses, and the alterations facilitate plant community to optimize their growth, development, and endogenous physiology to environmental fluctuations. Here, we report that Arabidopsis thaliana plants that experience high salinity emit VOCs and trigger induction of high salt resistance in neighboring plants. VOC emission of emitter plants is likely correlative to the plant damages to high salt, and VOC-fumigated receiver plants acquire high salt tolerance. The VOC-induced stress tolerance is independent of conventional abscisic acid (ABA) and salt stress signaling pathways. Together, this study demonstrates that salt-induced Arabidopsis VOCs are relevant in priming stress tolerance in neighboring plants. In addition, it also provides insight into how VOCs elicit stress responses in plant community.
Plants are exposed to adverse abiotic and biotic stresses, such as low temperature, drought, high salinity, wounding, and pathogen attack. They have evolutionarily evolved a variety of defense mechanisms to cope with the environmental challenges. Conventional adaptive mechanisms include intra-plant communication that activates and primes plant tolerance to environmental stresses, establishing systemic acquired resistance (SAR).Citation1,Citation2,Citation3 Intriguingly, plants have also developed inter-plant communication through volatile signals to share awareness of the environmental changes.Citation4,Citation5,Citation6
More than 100,000 chemicals are synthesized in plants,Citation7 and at least 1700 of which are released into the atmosphere in significant amounts.Citation8 In particular, a significant step forward has been recently made in understanding ecological and physiological roles of volatile organic compounds (VOCs) that are defined as organic lipophilic compounds with high vapor pressure,Citation9 including methanol, ethanol, formaldehyde, acetaldehyde, methane, ethylene, amino acid-derived products, green leaf volatiles (GLVs), and terpenes (isoprenoids).Citation10,Citation11
The emission of VOCs is elaborately controlled by environmental factors and mediates a variety of interactions between plants and other organisms in ecosystems.Citation8,Citation11 Primary function of VOCs is to facilitate plant community to defend against herbivores and microorganisms.Citation12,Citation13 Indeed, the VOC exposure primes defense responses in neighboring plants, incurring fitness costs.Citation14,Citation15 It also enables indirect defense by attracting predators or parasitoids that increase predation pressure on herbivores.Citation16 In addition, particular emphasis has been placed on the role of VOCs in attracting beneficial insects or microbes, such as pollinators and symbiotic organisms,Citation17,Citation18 demonstrating that plant VOCs are crucial for plant survival and reproduction.
Abiotic stresses, such as drought, high salinity, reduced nutrient availability, temperature extremes, and UV radiation, are the most limiting factors for plant growth, development, and productivity. Several lines of evidence have illustrated that some VOC signals are potentially engaged in eliciting priming plant responses to abiotic stresses in order to minimize their damages.Citation8,Citation11 Nonetheless, the biological relevance and mechanism of inter-plant communication about the presence of upcoming environmental stress are still largely elusive.
We wanted to know whether abiotic stress induces emission of plant VOCs to alert neighboring plants and elicit their subsequent stress responses. To examine this, we applied salt stress, one of pervasive environmental stress, to emitter plants (emitters) and investigated the effect of airborne signals on stress adaptation of neighboring plants (receivers). We used two-compartment-dish plates for germinating Arabidopsis wild-type (Columbia-0) seeds on one compartment containing half-strength Murashige and Skoog (MS)-agar and the second compartment containing half-strength MS-agar with high salt (50, 100, or 150 mM NaCl) (). The plates that allow to exchange VOCs between compartments were sealed and incubated at 23 °C for 2 wk under long day (16-light/8-dark) conditions (120 μmol m−2 s−1 white light).
Figure 1. Effects of VOCs emitted from Arabidopsis plants grown under high salinity on salt tolerance in neighboring plants. Arabidopsis wild-type seeds (Col-0) were germinated in two-compartment dish plates that have one compartment containing half-strength MS-agar and another compartment containing half-strength MS-agar with high salt (50, 100, or 150 mM NaCl) (A). Growth and development of two-week-old receiver plants grown together with emitter plants were compared (B). Two-week-old receiver plants grown with emitter plants exposed to different concentrations of NaCl were transferred to MS-agar supplemented with 150 mM NaCl (C). Biological triplicates were averaged. Different letters represent a significant difference at P < 0.05 (oneway ANOVA with Fisher’s post hoc test). Bars indicate standard error of the mean.
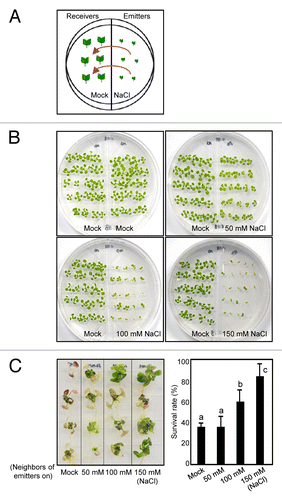
We first compared growth and development of receiver plants that were grown with emitter plants exposed to different concentration of high salt. While emitter plants were significantly influenced by high salt, growth patterns of receiver plants were not greatly altered (). We then investigated whether VOCs released from emitter plants contribute to establishing induced resistance of receiver plants to high salinity. Two-week-old receiver plants were transferred to half-strength MS-agar plates supplemented with 150 mM NaCl. Receiver plants grown with emitter plants on 150 mM NaCl were more tolerant than those that have emitter plants on lower NaCl concentrations (), indicating that salt-elicited VOCs are relevant in priming salt tolerance in neighboring plants and that emission of VOCs is probably correlative to high salt concentration.
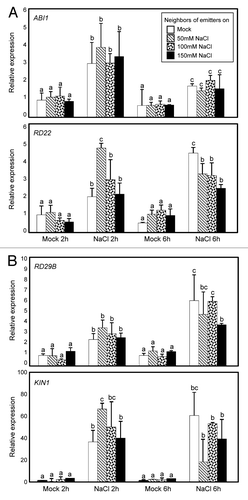
To understand molecular mechanisms underlying the induced resistance of receiver plants to high salinity, we incubated 2-wk-old receiver plants in MS-liquid medium supplemented with 150 mM NaCl for up to 6 h and monitored expression of stress-responsive genes. ABA responses substantially overlap with responses to high salinity,Citation19 but ABA signaling genes, such as ABA INSENSITIVE 1 (ABII) and RESPONSIVE TO DESICCATION 22 (RD22), were uninfluenced in receiver plants, which were exposed to a suite of salt-elicited VOCs, compared with wild-type plants (). In addition, key signaling integrators for abiotic stress pathways, such as RD29B and KOLD-INDUCED 1 (KINI), were also unaffected (). These results suggest that VOCs prime stress tolerance of receiver plants through as-yet-unidentified signaling pathways and mechanisms.
Figure 2. Effects of high salt-elicited plant VOCs on transcript accumulation of salt-responsive genes in neighboring plants. Two-week-old receiver plants grown with emitter plants exposed to different concentrations of high salt were transferred to MS-liquid medium supplemented with 150 mM NaCl and incubated for up to 6h. Transcript accumulation of ABA- (A) and salt-responsive (B) genes was analyzed by quantitative RT-PCR (qRT-PCR). Biological triplicates were averaged. Different letters represent a significant difference at P < 0.05 (oneway ANOVA with Fisher’s post hoc test). Bars indicate standard error of the mean.
It has been considered that Arabidopsis may not be a good species to study chemical exchanges between plants, because their VOCs are not abundant and colorful. Nonetheless, Arabidopsis is still beneficial considering the accessibilities of genomic information and diverse genetic resources. As an initial attempt, this study demonstrates that airborne signals emitted from Arabidopsis plants exposed to high salinity are biologically relevant in eliciting stress adaptive responses in neighboring plants yet experienced environmental stress. The VOCs seem to provoke priming responses, ensuring faster and stronger activation of physiological responses upon exposure to environmental challenges.Citation13,Citation14
Environmental stresses increase abundance of VOC emission and alter quality of VOCs.Citation8 Hence, one of the major challenges is to identify exchanged VOCs and elucidate roles of the individual compound. While high salinity-elicited chemicals have not been intensively identified, a few classes of VOCs are postulated to be involved in salt stress responses. A variety of terpene volatiles that are the major constituents of the plant volatilome, such as isoprenes/hemiterpenes (C5), monoterpenes (C10), and sesquiterpenes (C15), play a particular role in plant protection to thermal and oxidative stresses in a range of plant species.Citation9 Volatile terpene emission is stimulated by a wide range of environmental stresses.Citation8,Citation11 Then they neutralize reactive oxygen species (ROS), possibly due to the presence of conjugated double bonds, and protect plants from negative effects caused by environmental stresses, indicating the antioxidant properties.Citation20,Citation21 In agreement with this, inhibition of isoprene and monoterpene biosynthesis results in enhanced oxidative damages and reduced photosynthetic performance.Citation22 By contrast, plants emitting abundant isoprene show enhanced tolerance to high light, heat stress, ozone, and other ROS.Citation23 Given that virtually all environmental stresses involve ROS accumulation, terpenes may also contribute to relieving damages to abiotic stresses, including high salt, in plant community.
In addition to their role as antioxidant agents, volatile terpenes also stabilize cellular membranes,Citation24 because of their lipophilic nature. They are efficiently incorporated into phospholipid bilayer of cellular membranes and ensure structural persistency of the membranes by enhancing hydrophobic interactions. Consistently, plants fumigated with isoprenes and monoterpenes are resistant to heat stress that disrupts cellular membranes.Citation25 In this regard, sustained volatile terpene synthesis under high salinity may also be relevant in priming salt tolerance in neighboring plants.Citation26
Oxylipins are another important chemical blend of VOCs. They are derived from polyunsaturated fatty acids through the action of lipoxygenase (LOX) enzymes.Citation27 The oxylipins are subsequently cleaved into GLVs (C6- or LOX-products) by 13-(S)-hydroperoxide lyase (HPL).Citation8 GLVs include saturated or monounsaturated aldehydes, alcohols and esters, and their diverse configuration enable to have different sensory properties.Citation8 Emission of GLVs is rapidly elicited upon mechanical damages and herbivory attack, which accompany membrane denaturations and/or damages.Citation28 Consistent with this, those are also released with significant level in response to drought and salt stresses that entails cellular damages,Citation29 supposing the role of GLVs in salt stress adaptation.
Transient induction of methanol and/or methane can be engaged in the plant-plant communication under high salinity. Both volatile compounds are derived from degradation of cell wall pectins. Methanol emission, which is catalyzed by pectin methylesterases (PMEs), is induced by cellular mechanical damages, such as herbivore feeding and wounding.Citation30 Methoxyl groups of pectin can be a source of aerobic methane. It has been shown that UV radiation induces plants to produce methane though ROS-mediated cell wall breakdown.Citation31 Since environmental stresses also involve cellular damages as well as ROS accumulation, aerobic methanol and methane production can be speculated in salt-damaged plants.
In addition, the gaseous phytohormone ethylene can also be responsible for high salt adaptation. Ethylene is significantly induced by ABA, drought, and high salinity.Citation32 The rate-limiting enzyme of ethylene synthesis, ACC synthase (ACS) that catalyzes production of ethylene precursor 1-aminocyclopropane-1-carboxylic acid (ACC), is also enzymatically activated in response to osmotic stress.Citation33 Transgenic plants overexpressing ethylene responsive transcriptional regulators exhibit increased salt tolerance,Citation34,Citation35 supporting that ethylene signaling is intimately related to priming salt responses in neighboring plants.
While further investigations are required to elucidate volatile compounds responsible for the plant-plant communications, it is still obvious that VOCs emitted from plant exposed to environmental stresses are relevant in priming stress tolerance in neighboring plants. Metabolic analyses and genetic manipulation will provide comprehensive views of inter-plant communication and biological mechanisms underlying volatile signal perception and interpretation.
| Abbreviations: | ||
| GLV | = | green leaf volatile |
| HPL | = | 13-(S)-hydroperoxide lyase |
| LOX | = | lipoxygenase |
| PME | = | pectin methylesterase |
| ROS | = | reactive oxygen species |
| SAR | = | systemic acquired resistance |
| VOC | = | volatile organic compound |
Disclosure of Potential Conflicts of Interest
No potential conflicts of interests were disclosed.
Acknowledgments
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2013R1A1A1004831). K.L. was supported by BK21 PLUS program in the Department of Bioactive Material Sciences.
References
- Baxter A, Mittler R, Suzuki N. ROS as key players in plant stress signalling. J Exp Bot 2013; http://dx.doi.org/10.1093/jxb/ert375; PMID: 24253197
- Zhang R, Zhu J, Cao HZ, An YR, Huang JJ, Chen XH, Mohammed N, Afrin S, Luo ZY. Molecular cloning and expression analysis of PDR1-like gene in ginseng subjected to salt and cold stresses or hormonal treatment. Plant Physiol Biochem 2013; 71:203 - 11; http://dx.doi.org/10.1016/j.plaphy.2013.07.011; PMID: 23968928
- Fu ZQ, Dong X. Systemic acquired resistance: turning local infection into global defense. Annu Rev Plant Biol 2013; 64:839 - 63; http://dx.doi.org/10.1146/annurev-arplant-042811-105606; PMID: 23373699
- Karban R, Shiojiri K, Huntzinger M, McCall AC. Damage-induced resistance in sagebrush: volatiles are key to intra- and interplant communication. Ecology 2006; 87:922 - 30; http://dx.doi.org/10.1890/0012-9658(2006)87[922:DRISVA]2.0.CO;2; PMID: 16676536
- Pearse IS, Hughes K, Shiojiri K, Ishizaki S, Karban R. Interplant volatile signaling in willows: revisiting the original talking trees. Oecologia 2013; 172:869 - 75; http://dx.doi.org/10.1007/s00442-013-2610-2; PMID: 23576105
- Yao Y, Danna CH, Zemp FJ, Titov V, Ciftci ON, Przybylski R, Ausubel FM, Kovalchuk I. UV-C-irradiated Arabidopsis and tobacco emit volatiles that trigger genomic instability in neighboring plants. Plant Cell 2011; 23:3842 - 52; http://dx.doi.org/10.1105/tpc.111.089003; PMID: 22028460
- Dicke M, Loreto F. Induced plant volatiles: from genes to climate change. Trends Plant Sci 2010; 15:115 - 7; http://dx.doi.org/10.1016/j.tplants.2010.01.007; PMID: 20137997
- Loreto F, Schnitzler JP. Abiotic stresses and induced BVOCs. Trends Plant Sci 2010; 15:154 - 66; http://dx.doi.org/10.1016/j.tplants.2009.12.006; PMID: 20133178
- Tholl D, Boland W, Hansel A, Loreto F, Röse US, Schnitzler JP. Practical approaches to plant volatile analysis. Plant J 2006; 45:540 - 60; http://dx.doi.org/10.1111/j.1365-313X.2005.02612.x; PMID: 16441348
- Maffei ME, Gertsch J, Appendino G. Plant volatiles: production, function and pharmacology. Nat Prod Rep 2011; 28:1359 - 80; http://dx.doi.org/10.1039/c1np00021g; PMID: 21670801
- Holopainen JK, Gershenzon J. Multiple stress factors and the emission of plant VOCs. Trends Plant Sci 2010; 15:176 - 84; http://dx.doi.org/10.1016/j.tplants.2010.01.006; PMID: 20144557
- Sarmento RA, Lemos F, Bleeker PM, Schuurink RC, Pallini A, Oliveira MG, Lima ER, Kant M, Sabelis MW, Janssen A. A herbivore that manipulates plant defence. Ecol Lett 2011; 14:229 - 36; http://dx.doi.org/10.1111/j.1461-0248.2010.01575.x; PMID: 21299823
- Unsicker SB, Kunert G, Gershenzon J. Protective perfumes: the role of vegetative volatiles in plant defense against herbivores. Curr Opin Plant Biol 2009; 12:479 - 85; http://dx.doi.org/10.1016/j.pbi.2009.04.001; PMID: 19467919
- Peng J, van Loon JJ, Zheng S, Dicke M. Herbivore-induced volatiles of cabbage (Brassica oleracea) prime defence responses in neighbouring intact plants. Plant Biol (Stuttg) 2011; 13:276 - 84; http://dx.doi.org/10.1111/j.1438-8677.2010.00364.x; PMID: 21309974
- Ali M, Sugimoto K, Ramadan A, Arimura G. Memory of plant communications for priming anti-herbivore responses. Sci Rep 2013; 3:1872; http://dx.doi.org/10.1038/srep01872; PMID: 23695148
- Degenhardt J. Indirect defense responses to herbivory in grasses. Plant Physiol 2009; 149:96 - 102; http://dx.doi.org/10.1104/pp.108.128975; PMID: 19126700
- Brodmann J, Emer D, Ayasse M. Pollinator attraction of the wasp-flower Scrophularia umbrosa (Scrophulariaceae). Plant Biol (Stuttg) 2012; 14:500 - 5; http://dx.doi.org/10.1111/j.1438-8677.2011.00525.x; PMID: 22188305
- Weber W, Daoud-El Baba M, Fussenegger M. Synthetic ecosystems based on airborne inter- and intrakingdom communication. Proc Natl Acad Sci U S A 2007; 104:10435 - 40; http://dx.doi.org/10.1073/pnas.0701382104; PMID: 17551014
- Uno Y, Furihata T, Abe H, Yoshida R, Shinozaki K, Yamaguchi-Shinozaki K. Arabidopsis basic leucine zipper transcription factors involved in an abscisic acid-dependent signal transduction pathway under drought and high-salinity conditions. Proc Natl Acad Sci U S A 2000; 97:11632 - 7; http://dx.doi.org/10.1073/pnas.190309197; PMID: 11005831
- Loreto F, Mannozzi M, Maris C, Nascetti P, Ferranti F, Pasqualini S. Ozone quenching properties of isoprene and its antioxidant role in leaves. Plant Physiol 2001; 126:993 - 1000; http://dx.doi.org/10.1104/pp.126.3.993; PMID: 11457950
- Affek HP, Yakir D. Protection by isoprene against singlet oxygen in leaves. Plant Physiol 2002; 129:269 - 77; http://dx.doi.org/10.1104/pp.010909; PMID: 12011357
- Vickers CE, Possell M, Cojocariu CI, Velikova VB, Laothawornkitkul J, Ryan A, Mullineaux PM, Nicholas Hewitt C. Isoprene synthesis protects transgenic tobacco plants from oxidative stress. Plant Cell Environ 2009; 32:520 - 31; http://dx.doi.org/10.1111/j.1365-3040.2009.01946.x; PMID: 19183288
- Calfapietra C, Mugnozza GS, Karnosky DF, Loreto F, Sharkey TD. Isoprene emission rates under elevated CO2 and O3 in two field-grown aspen clones differing in their sensitivity to O3. New Phytol 2008; 179:55 - 61; http://dx.doi.org/10.1111/j.1469-8137.2008.02493.x; PMID: 18557875
- Sharkey TD, Wiberley AE, Donohue AR. Isoprene emission from plants: why and how. Ann Bot 2008; 101:5 - 18; http://dx.doi.org/10.1093/aob/mcm240; PMID: 17921528
- Singsaas EL, Lerdau M, Winter K, Sharkey TD. Isoprene increases thermotolerance of isoprene-emitting species. Plant Physiol 1997; 115:1413 - 20; PMID: 12223874
- Loreto F, Delfine S. Emission of isoprene from salt-stressed Eucalyptus globulus leaves. Plant Physiol 2000; 123:1605 - 10; http://dx.doi.org/10.1104/pp.123.4.1605; PMID: 10938376
- Feussner I, Wasternack C. The lipoxygenase pathway. Annu Rev Plant Biol 2002; 53:275 - 97; http://dx.doi.org/10.1146/annurev.arplant.53.100301.135248; PMID: 12221977
- Behnke K, Kleist E, Uerlings R, Wildt J, Rennenberg H, Schnitzler JP. RNAi-mediated suppression of isoprene biosynthesis in hybrid poplar impacts ozone tolerance. Tree Physiol 2009; 29:725 - 36; http://dx.doi.org/10.1093/treephys/tpp009; PMID: 19324699
- Capitani D, Brilli F, Mannina L, Proietti N, Loreto F. In situ investigation of leaf water status by portable unilateral nuclear magnetic resonance. Plant Physiol 2009; 149:1638 - 47; http://dx.doi.org/10.1104/pp.108.128884; PMID: 19193862
- Körner E, von Dahl CC, Bonaventure G, Baldwin IT. Pectin methylesterase NaPME1 contributes to the emission of methanol during insect herbivory and to the elicitation of defence responses in Nicotiana attenuata.. J Exp Bot 2009; 60:2631 - 40; http://dx.doi.org/10.1093/jxb/erp106; PMID: 19380422
- Messenger DJ, McLeod AR, Fry SC. The role of ultraviolet radiation, photosensitizers, reactive oxygen species and ester groups in mechanisms of methane formation from pectin. Plant Cell Environ 2009; 32:1 - 9; http://dx.doi.org/10.1111/j.1365-3040.2008.01892.x; PMID: 18811731
- Bleecker AB, Kende H. Ethylene: a gaseous signal molecule in plants. Annu Rev Cell Dev Biol 2000; 16:1 - 18; http://dx.doi.org/10.1146/annurev.cellbio.16.1.1; PMID: 11031228
- Morgan PW, Drew MC. Ethylene and plant responses to stress. Physiol Plant 1997; 100:620 - 30; http://dx.doi.org/10.1111/j.1399-3054.1997.tb03068.x
- Pan Y, Seymour GB, Lu C, Hu Z, Chen X, Chen G. An ethylene response factor (ERF5) promoting adaptation to drought and salt tolerance in tomato. Plant Cell Rep 2012; 31:349 - 60; http://dx.doi.org/10.1007/s00299-011-1170-3; PMID: 22038370
- Zhang Z, Li F, Li D, Zhang H, Huang R. Expression of ethylene response factor JERF1 in rice improves tolerance to drought. Planta 2010; 232:765 - 74; http://dx.doi.org/10.1007/s00425-010-1208-8; PMID: 20574667