
Abstract
Previously we described that different priming stimuli trigger common metabolomic responses against P. cucumerina. Furthermore we showed that several primed metabolites were present following independent priming inducers such as natural constitutive priming promoted by gene mutations and chemical priming induced by the β-aminobutyric acid (BABA). Despite we found a common metabolomic fingerprint, in the present research we focus our attention in specific metabolites that are primed differentially by a mutation in the NRT2.1 gene (lin1 mutant) and BABA treatments against P. cucumerina. Around eight hundred compounds were overaccumulated in the resistant mutant lin1 and in BABA treated plants upon infection. Among them 404 and 412 were specific of each priming condition while 103 compounds were shared by both. Flavonoids and lignans were specifically accumulated in lin1 in response to the fungal attack, while tyrosine, purine metabolism, and aromatic carbon degradation compounds were only accumulated in BABA primed plants upon infection. However, most metabolites differentially accumulated by the two priming conditions belonged to the same metabolic pathways, suggesting that different priming stimuli, upon a given biotic stress, may stimulate similar pathways but activate specific differences depending on the priming stimulus.
Priming is a physiological state of the plant that regulates several defense mechanisms to cope with pathogens of different life style.Citation1 Priming state can be achieved by exogenous application of chemical compounds or by gene alterations that makes the plant react faster and stronger to abiotic and biotic challenge. As a chemical priming inducer the β-aminobutyric acid (BABA) has become in one of the most efficient priming compounds effective against broad range of pathogen diseases stimulating the natural defense of the plant depending on the life style of the pathogen. For instance, it was demonstrated that BABA induces resistance against the hemibiotrophic bacteria Pseudomonas syringae (Pst) and the downy mildew Hyaloperonospora arabidopsidis activating PR1 gene expression against the bacteria and callose depositions at the sites of infection against the biotrophic oomycete.Citation2 BABA-induced resistance (BABA-IR) was also effective in Arabodopsis against the pathogenic fungi P. cucumerina and Alternaria brassicicola by increasing callose deposition and stimulating Reactive oxygen species (ROS).Citation3,Citation4,Citation5 An augmented H2O2 accumulation at the sites of infection followed by callose deposition indicates that BABA-IR phenomenon involves several defense layers of resistance.
Constitutive priming also requires induction of defense barriers. The Arabidopsis mutant ocp3 was demonstrated to show an induction in callose accumulation and increased expression of defense marker genes such as PDF1.2 against P. cucumerina.Citation6 The Arabidopsis mutant lin1 which is blocked in a High Affinity nitrate Transporter (HATs) displayed constitutive primed expression of pathogenesis related genes PR1 and PR5 in response to Pst.Citation7 Besides, a recent publication demonstrated that lin1 acquire an inherent induced resistance against necrotrophic fungi P. cucumerina with no related fitness costs in terms of development and seed production, indicating that lin1 behaves as a priming mutant effective against a broad range of pathogens.Citation8
Horizontal primed responses involve modifications not only in ROS homeostasis, callose deposition and gene expression but also in hormone and metabolite levels. Attention in hormonal homeostasis has gained interest since hormonal responses play an important role in constitutive- and chemical-induced priming. It was previously reported that the indole derivative Indole-3-carboxylic acid (I3CA) behaves as a primed metabolite in BABA-IR since its accumulation in infected plants was higher in BABA- than in water-treated plants.Citation9 Furthermore, I3CA was detected to be not only over-accumulated in BABA but also in other two constitutive priming Arabidopsis mutants indicating that this molecule, among others, is part of a common profile of primed metabolites against P. cucumerina.Citation8
However, different priming stimuli do not always stimulate the same defense mechanisms. A good example is the induction of jasmonic acid/ethylene defense marker gene PDF1.2. This gene was not related to BABA-IR since it does not presents a priming behavior in the Arabidopsis-P. cucumerina interaction.Citation3 Recently, this gene was involved in ocp3 resistance against this pathogen since ocp3 inoculated plants displayed higher expression compared with control plants,Citation6 suggesting that, despite there is a common defense core in primed plants stimulated by different stimuli, there are also specific responses depending on the way the plants achieve the priming state.
Therefore in the present research we focus our attention in those specific responses observed following constitutive priming in lin1 compared with BABA-induced priming against P. cucumerina.
Priming Promotes Metabolic Reprocessing Driving to Specific and Common Over-Accumulated Metabolites
The PCA analysis of metabolites in Col-0 treated either with water or BABA and lin1 showed that metabolic changes imposed by BABA treatments in Arabidopsis in the absence of challenge overlap with the metabolome of water treatments. However, metabolomic changes in lin1 are significantly separated from Col-0 (). This means that, rather than treatments, the principal components of the metabolomic differences in the absence of infection are due to the genotype. A similar result is observed comparing natural and chemical priming with water treated plants following infection. After P. cucumerina infection, the metabolome of BABA and water treated plants overlap while the metabolome of infected lin1 separates significantly (). Thus it seems clear that BABA pretreatment induces small changes that affect the way the plant acquires an alarm state, making the response to the infection more effective ().
Figure 1. Non-supervised PCA representation of signals obtained from a non targeted analysis by HPLC-QTOFMS to monitor metabolomic changes during fungal invasion. (A) PCA1 vs PCA2 and (B) PCA1 vs PCA3. Three week old seedlings were sprayed inoculated with 10E3 spores.ml−1 of P. cucumerina and samples for analysis were collected 48hpi. Leaf material from 15 individual plants were pooled together for each treatment x genotype combination. Data points represent two technical replicates from three independent experiments injected randomly into the HPLC-QTOFMS. The signals corresponding to different treatments were compared using the non-parametric Kruskal-Wallis test, and only data with a P value lower than 0.01 between groups was used for subsequent processing.
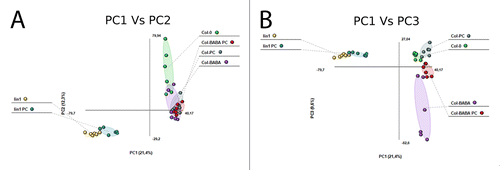
Analysis of the principal components 1 and 3 revealed that, in addition to the strong differences are still visible in the lin1 metabolome, other significant differences due to BABA treatments are also detected. The comparative analysis of the PC1 vs PC3 shows that the chemical and constitutive natural priming and the infection contribute to the differences in the metabolome since there is no overlap between the groups of metabolites in any of the treatments ().
We showed before that constitutive and chemical priming stimulus can equally protect Arabidopsis against P. cucumerina. Among these primed metabolites, a set of compounds that applied exogenously can protect Arabidopsis such as Indole-3-carboxylic acid and hypoxanthine was found. These were identified following different priming stimuli.Citation8 Despite these experimental evidences, it cannot be discarded that specific changes observed in lin1 and Col-0-BABA treated plants can contribute to priming defense.
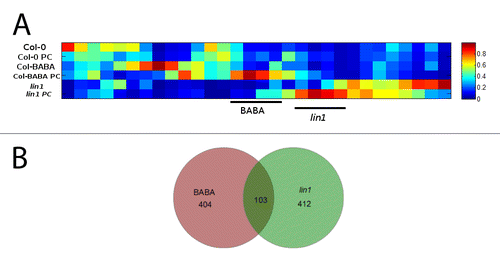
A dedicated heat map and Venn diagram analysis () showed few clusters of signals shared by both priming stimuli, around 103 compounds (). On the other hand, several clusters of compounds are specific to each priming stimulus. Among these differential clusters, 404 metabolites are specifically accumulated in BABA-treated Col-0 plants upon infection, while 412 metabolites are specifically accumulated in lin1 infected plants ().
Figure 2. (A) Heat map analysis of the signals extracted from the Kruskal-Wallis test with a P value lower than 0.01 between groups performed with Marvis (Filter and Cluster packages) and (B) Venn diagrams of detected primed signals.
Different Priming Stimuli Activate Similar Metabolic Pathways Promoting Specific Metabolite Accumulation
Despite 816 metabolites are specifically primed either by BABA or in lin1, many of these compounds belong to the same metabolic pathways. This reinforces the hypothesis that priming is a horizontal phenomenon that activates multiple signaling pathways.Citation5 In fact, phenylpropanoid-, shikimate-, Trp-derivatives and glucosinolate biosynthetic pathways are primed by both stimuli, while a set of flavonoids and lignans were specifically accumulated in lin1 in response to the P. cucumerina and tyrosine derivatives, purine metabolites and aromatic carbon degradation compounds were only accumulated in BABA primed plants upon infection (). This highlights that both specific and common responses can be triggered by different priming stimuli. Remarkably, all these common pathways triggered by both stimuli have been obtained by selecting clusters of differentially accumulated compounds (). Therefore, different priming stimuli may trigger the plant to achieve similar metabolic goals by activating specific compounds among shared metabolic pathways.
Table 1. Tentatively identified pathways
| Abbreviations: | ||
| P. cucumerina | = | Plectosphaerella cucumerina |
| BABA | = | β-aminobutyric |
| Pst | = | Pseudomonas syringae |
| IR | = | Induced resistance |
| ROS | = | Reactive oxygen species |
| HATs | = | High Affinity nitrate Transporter |
| PR1 and PR5 | = | Pathogenesis related genes |
| I3CA | = | Indole-3-carboxylic acid |
| PCA | = | Principal component analysis |
| Trp | = | Triptophan |
| lin1 | = | Lateral root initation 1 |
| ocp3 | = | Overexpressor of cationic peroxidase |
| PDF1 | = | Promoter of the plant defensin gene |
| NRT2.1 | = | Nitrogen Transporter |
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
We thank the SCIC of the Universitat Jaume I and particularly Cristian Barrera for his technical support. This work was financially supported by the Plan de Promoción de la Investigación-Universitat Jaume I P1·1B2013-68.
References
- Pastor V, Luna E, Mauch-Mani B, Ton J, Flors V. Primed plants do not forget. Environ Exp Bot 2012; 94:46 - 56; http://dx.doi.org/10.1016/j.envexpbot.2012.02.013
- Zimmerli L, Métraux JP, Mauch-Mani B. β-Aminobutyric acid-induced protection of Arabidopsis against the necrotrophic fungus Botrytis cinerea. Plant Physiol 2001; 126:517 - 23; http://dx.doi.org/10.1104/pp.126.2.517; PMID: 11402183
- Ton J, Mauch-Mani B. β-amino-butyric acid-induced resistance against necrotrophic pathogens is based on ABA-dependent priming for callose. Plant J 2004; 38:119 - 30; http://dx.doi.org/10.1111/j.1365-313X.2004.02028.x; PMID: 15053765
- Flors V, Ton J, van Doorn R, Jakab G, García-Agustín P, Mauch-Mani B. Interplay between JA, SA and ABA signalling during basal and induced resistance against Pseudomonas syringae and Alternaria brassicicola.. Plant J 2008; 54:81 - 92; http://dx.doi.org/10.1111/j.1365-313X.2007.03397.x; PMID: 18088307
- Pastor V, Luna E, Ton J, Cerezo M, García-Agustín P, Flors V. Fine tuning of reactive oxygen species homeostasis regulates primed immune responses in Arabidopsis. Mol Plant Microbe Interact 2013; 26:1334 - 44; http://dx.doi.org/10.1094/MPMI-04-13-0117-R; PMID: 24088017
- García-Andrade J, Ramírez V, Flors V, Vera P. Arabidopsis ocp3 mutant reveals a mechanism linking ABA and JA to pathogen-induced callose deposition. Plant J 2011; 67:783 - 94; http://dx.doi.org/10.1111/j.1365-313X.2011.04633.x; PMID: 21564353
- Camañes G, Pastor V, Cerezo M, García-Andrade J, Vicedo B, García-Agustín P, Flors V. A deletion in NRT2.1 attenuates Pseudomonas syringae-induced hormonal perturbation, resulting in primed plant defenses. Plant Physiol 2012; 158:1054 - 66; http://dx.doi.org/10.1104/pp.111.184424; PMID: 22158760
- Gamir J, Pastor V, Kaever A, Cerezo M, Flors V. Targeting novel chemical and constitutive primed metabolites against Plectosphaerella cucumerina.. Plant J 2014; 78:227 - 40; http://dx.doi.org/10.1111/tpj.12465; PMID: 24506441
- Gamir J, Pastor V, Cerezo M, Flors V. Identification of indole-3-carboxylic acid as mediator of priming against Plectosphaerella cucumerina.. Plant Physiol Biochem 2012; 61:169 - 79; http://dx.doi.org/10.1016/j.plaphy.2012.10.004; PMID: 23116603