Abstract
BAK1 and BKK1 are two functionally redundant leucine-rich repeat receptor-like protein kinases (LRR-RLKs) involved in brassinosteroid signal transduction by their direct interactions with the BR receptor, BRI1. Recent studies from our group and others indicated that the two RLKs also play critical roles in regulating pathogen-related and pathogen-unrelated cell-death controls. Genetic data suggest that the two kinases are essential for plant survival because the double mutants show spontaneous cell-death and seedling lethality phenotypes. Physiological analyses further suggest that the cell-death of the double mutant is triggered by the light, as dark-grown seedlings do not show any cell-death symptoms. These observations indicate that BAK1 and BKK1 regulate a novel signaling pathway to detoxify or to limit the production of a yet unknown toxin/toxins produced by plants under light conditions.
Addendum to: He K, Gou X, Yuan T, Lin H, Asami T, Yoshida S, Russell SD, Li J. BAK1 and BKK1 regulates brassinosteroid-dependent growth and brassinosteroid-independent cell-death pathways. Curr Biol; 1109-15.
Plant receptor-like protein kinases (RLKs) are transmembrane proteins essential for cell-to-cell communications. A typical RLK is composed of a cell-surface receptor domain which can sense and perceive diversified signaling molecules within the extracellular space, a transmembrane domain anchoring the protein to the plasma membrane, and a cytoplasmic kinase domain whose activity can often be regulated by the conformation change in the receptor domain upon the binding of the signaling molecules to the receptor. The unique structure of RLKs suggests that these kinases may act as key switches in triggering many signal transduction cascades which greatly influence plant growth and development. Recent studies support this notion, as the functions of more and more RLKs have been revealed, and these RLKs indeed play critical roles in mediating many physiological processes such as steroidal hormone action, pathogenesis responses, and recognition of various peptide signals.Citation1–Citation3 There are more than 600 RLKs in the Arabidopsis genome.Citation4,Citation5 Based on the structure of the receptor domains, RLKs can be divided into more than 10 subfamilies. Among them, LRR-RLKs belong to the largest group consisting of at least 220 members. The functions of only a small fraction of RLKs have been revealed.
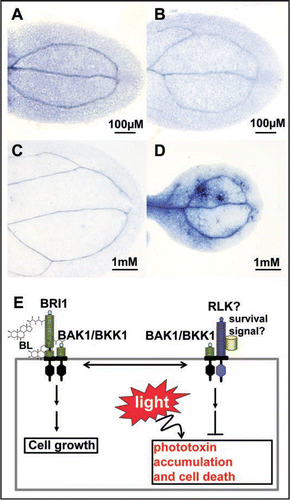
BAK1 is a typical LRR-RLK, identified via an activation tagging genetic screen for suppressors of a weak BR receptor (BRI1) mutant called bri1–5, and via a yeast two-hybrid screen for BRI1 kinase domain physical interactors.Citation6,Citation7 Although the detailed molecular mechanisms of BAK1 in activating the BR signaling pathway is still mysterious, the in vivo interaction between BAK1 and BRI1 is clearly ligand (BR)-dependent.Citation8 The mutual phosphorylation of the two kinases is also BR-dependent.Citation8 BKK1, the closest homolog of BAK1, was identified as a functionally redundant protein of BAK1.Citation9 Interestingly, the double null mutant of BAK1 and BKK1, bak1–4 bkk1-1, did not show a typical bri1 phenotype but showed a spontaneous cell-death phenotype under a normal growth condition. This unexpected result suggests that BAK1 and BKK1 may have more roles than their functions in BR signal transduction. This hypothesis is supported by the recent discovery of BAK1 in mediating pathogen-related signaling pathways in order to regulate innate immunity and cell-death control.Citation10–Citation12 The spontaneous cell-death seen in the bak1–4 bkk1-1 double mutant, however, is not caused by the challenges from pathogens;Citation10 it is also unlikely to be the result from the disruption of the FLS2-dependent innate immunity pathway,Citation11,Citation12 as overexpression or T-DNA disruption of the RLK gene, FLS2, does not show a phenotype similar to that of the bak1–4 bkk1-1 double mutant. In addition, the cell-death phenotype of the double mutant occurs even in a sterile growth condition, suggesting that the pathogens are not the key triggers of cell-death in the bak1–4 bkk1-1 double mutant. Early results indicated that the double mutant seedlings are indistinguishable from the wild-type seedlings during the first 4–5 days after germination but quickly show terminating growth and cotyledon necrosis phenotypes a week after germination.Citation9 This observation prompted a test of whether light is a true trigger for cell-death seen in the double mutant. Both wild-type and the double mutant were planted in the dark and long-day lighting conditions. Cotyledons from eight-day-old seedlings were stained with Trypan blue to examine cell-death symptoms of the seedlings grown under different illumination conditions.Citation13 Both the dark-grown wild-type and the double mutant seedlings showed no cell-death symptoms on their cotyledons at any time during a three-week experimental period ( and B). Under a long-day lighting condition, on the other hand, cotyledons from the double mutant, but not from the wild-type, exhibited severe cell-death symptoms ( and D). Three weeks after germination, the double mutant seedlings growing under a long-day lighting condition was completely dead, the ones under the dark condition were still healthy and showed no cell-death symptoms (data not shown).
Based on our current results, it is apparent that the double mutant is more vulnerable to light. It is probable that the mutant lost its capability to detoxify or to restrict the production of an unknown toxin/toxins naturally generated by plants under a light condition. The wild-type plants may also produce the toxin/toxins, but BAK1 and BKK1 can direct a signal transduction pathway to constantly check and eliminate extra amount of the toxin/toxins (). Under a sterile growth condition, BAK1 and BKK1 are likely involved in both the BR signaling pathway to positively regulate cell growth and in a novel pathway to negatively control cell-death. Under normal environmental conditions (not sterile condition), BAK1 might also be recruited to participate in the innate immunity pathway via its interaction with FLS2 and other RLKs. Based on the model from the BRI1/BAK1 signal transduction, there might be another RLK which can pair with BAK1 or BKK1 in controlling the light-dependent cell-death process. An unknown “survival signal” could be an unknown metabolite or the toxin/toxins causing the cell-death. Under the current model, the “survival signal” may activate the BAK1- and BKK1-associated stress defense pathway and constantly check the levels of the light-dependent toxin/toxins in the plants. The homeostasis of the toxin/toxins is therefore strictly under control. If both BAK1 and BKK1 are removed, as in the case of the double mutant, the plant loses its capability to check the levels of the toxin/toxins. The uncontrolled accumulation of the toxin/toxins is likely the ultimate cause of the spontaneous cell-death observed in the double mutant.
Perspective
Elucidating the molecular mechanisms of the BAK1 and BKK1 related cell-death control pathway will rely on future identification of the light-dependent toxin/toxins. It is possible that reactive oxygen species (ROS) are the toxins which caused the cell-death in the double mutant. ROS are highly reactive and toxic compounds which are principally generated by plants during light-dependent photosynthesis and photorespiration.Citation14 ROS are composed of hydrogen peroxide (H2O2), singlet oxygen (1O2), superoxide anion (O2−) and hydroxyl radical (HO). Although ROS are crucial for maintaining the redox homeostasis of cells, extra amount of ROS can cause irreversible cell-death. In addition, cell-death seen in the bak1–4 bkk1-1 double mutant is always accompanied by uncontrolled accumulation of ROS.Citation9 Future studies should be concentrated on demonstrating whether ROS are the authentic triggers of cell-death in the double mutant, or identifying the true toxin/toxins causing cell-death of the double mutant. Genetic analysis will be crucial for further defining this novel stress-related cell-death control pathway. A weak double mutant recently identified by our group will be an ideal tool to identify cell-death genetic suppressors (unpublished). Further studies on this subject should shed light into our understanding hormonal actions, plant stress tolerance and detailed molecular mechanisms of the interplay among several BAK1- and BKK1-regulated signaling pathways.
Figures and Tables
Figure 1 BAK1 and BKK1 regulate a light-dependent cell-death control pathway. (A) A representative Trypan blue stained wild-type cotyledon from a dark grown seedling on a ½ MS plate; (B) A representative Trypan blue stained bak1–4 bkk1-1 cotyledon from a dark grown seedling on a ½ MS plate; (C) A representative Trypan blue stained wild-type cotyledon from a long-day light-grown seedling on a ½ MS plate; (D) A representative Trypan blue stained bak1–4 bkk1-1 cotyledon from a long-day light-grown seedling on a ½ MS plate; (E) A hypothetical model of BAK1 and BKK1 in regulating both the BR signaling pathway to promote cell growth, and a novel light-dependent cell-death control pathway to prevent plants from unnecessary cell-death. Under a light condition, plants naturally produce unknown toxins (phototoxins), whose accumulation can lead to the cell-death. BAK1 and BKK1 likely mediate a signaling pathway to constantly check and limit the levels of these toxins.
Addendum to:
References
- Becraft PW. Receptor kinase signaling in plant development. Annu Rev Cell Dev Biol 2002; 18:163 - 192
- Dievart A, Clark SE. LRR-containing receptors regulating plant development and defense. Development 2004; 131:251 - 261
- Morillo SA, Tax FE. Functional analysis of receptor-like kinases in monocots and dicots. Curr Opin Plant Biol 2006; 9:460 - 469
- Arabidopsis Genome Initiative. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 2000; 408:796 - 815
- Shiu SH, Bleecker AB. Receptor-like kinases from Arabidopsis form a monophyletic gene family related to animal receptor kinases. Proc Natl Acad Sci USA 2001; 98:10763 - 10768
- Li J, Wen J, Lease KA, Doke JT, Tax FE, Walker JC. BAK1, an Arabidopsis LRR receptor-like protein kinase, interacts with BRI1 and modulates brassinosteroid signaling. Cell 2002; 110:213 - 222
- Nam KH, Li J. BRI1/BAK1, a receptor kinase pair mediating brassinosteroid signaling. Cell 2002; 110:203 - 212
- Wang X, Goshe MB, Soderblom EJ, Phinney BS, Kuchar JA, Li J, Asami T, Yoshida S, Huber SC, Clouse SD. Identification and functional analysis of in vivo phosphorylation sites of the Arabidopsis BRASSINOSTEROID-INSENSITIVE1 receptor kinase. Plant Cell 2005; 17:1685 - 1703
- He K, Gou X, Yuan T, Lin H, Asami T, Yoshida S, Russell SD, Li J. BAK1 and BKK1 regulate brassinosteroid-dependent growth and brassinosteroid-independent cell-death pathways. Curr Biol 2007; 17:1109 - 1115
- Kemmerling B, Schwedt A, Rodriguez P, Mazzotta S, Frank M, Qamar SA, Mengiste T, Betsuyaku S, Parker JE, Mussig C, Thomma BP, Albrecht C, de Vries SC, Hirt H, Nurnberger T. The BRI1-associated kinase 1, BAK1, has a brassinolide-independent role in plant cell-death control. Curr Biol 2007; 17:1116 - 1122
- Chinchilla D, Zipfel C, Robatzek S, Kemmerling B, Nurnberger T, Jones JD, Felix G, Boller T. A flagellin-induced complex of the receptor FLS2 and BAK1 initiates plant defence. Nature 2007; 448:497 - 500
- Heese A, Hann DR, Gimenez Ibanez S, Jones AM, He K, Li J, Schroeder JI, Peck SC, Rathjen JP. The receptor-like kinase SERK3/BAK1 is a central regulator of innate immunity in plants. Proc Natl Acad Sci U S A 2007; 104:12217 - 12222
- Shirasu K, Lahaye T, Tan MW, Zhou F, Azevedo C, Schulze-Lefert P. A novel class of eukaryotic zinc-binding proteins is required for disease resistance signaling in barley and development in C. elegans. Cell 1999; 99:355 - 366
- Mittler R, Vanderauwera S, Gollery M, Van Breusegem F. Reactive oxygen gene network of plants. Trends Plant Sci 2004; 9:490 - 498