Abstract
Our recent work used novel methods to localize and track discrete vesicle populations in pollen tubes undergoing oscillatory growth. The results show that clathrin-dependent endocytosis occurs along the shank of the pollen tube, smooth vesicle endocytosis occurs at the tip, and exocytosis occurs in the subapical region. Here, growth of tobacco and lily pollen tubes is examined in greater temporal resolution using refraction-free high-resolution time-lapse differential interference contrast microscopy. Images were collected at 0.21 s intervals for 10 min, sequentially examined for millisecond details, compressed into video format and then examined for details of growth dynamics. The subapical growth zone is structurally fluid, with vesicle insertion into the plasma membrane, construction of new cell surface and cellular expansion. Incorporation of new membrane and wall materials causes localized disruption at the cell surface that precedes the start of the growth cycle by 3.44 ± 0.39 s in tobacco, and 1.02 ± 0.01 s in lily pollen tubes. Vesicle deposition increases after the start of the growth cycle and supports expansion of the growth zone. Growth reorientation involves a shift in the position and angle of the growth zone. In summary, these results support a new model of pollen tube growth.
Addendum to: Zonia L, Munnik T. Vesicle trafficking dynamics and visualization of zones of exocytosis and endocytosis in tobacco pollen tubes. J Exp Bot 2008; In press.
Introduction
It is crucial to understand how a cell grows before assigning functions to signals, proteins and effectors that regulate growth. The long-standing model of pollen tube growth considers that exocytosis occurs at the tip and that cell elongation is regulated primarily by cell wall yielding.Citation1 However, exocytosis at the apex had never been observed, and the role of osmotic pressure during growth had not been fully explored. Additionally, several reports were inconsistent with the model of tip-growth.Citation2–Citation4 Recent work has revealed that exocytosis occurs in the subapical region and that hydrodynamics integrates the rates of endo/exocytosis and growth.Citation5–Citation8 Exocytosis was observed using novel methods of FM pulse-chase labeling and refraction-free DIC microscopy.Citation8 In this Addendum, close examination of high-resolution time-lapse DIC images provides further evidence in support of the new model.
From Nascent Bulge to Organized Pollen Tube
In early stages of pollen germination, the nascent tube appears as a bulge extruding through one of the germination apertures (). Vesicles traffic from the pollen grain into the expanding bulge. The growth rate is slow and randomly fluctuating. The bulge lacks the cytological organization that characterizes longer tubes, particularly the thin tubular cell shape and the clear zone in the apical dome. During these early stages, vesicles are observed to incorporate along the entire apical membrane (, arrow).
As the pollen tube elongates, vesicle incorporation into the apical dome decreases, the cell organizes into the characteristic tube shape and the clear zone appears in the apical dome (). At the boundary between the hemispherical apical dome and the cylindrical tube, called the subapical region, there is a periodic appearance of a narrow annulus that is differentiated from the surrounding area by distinct surface properties (, arrow). Close examination of the time-lapse images and compressed videos reveals that this subapical zone undergoes elongation during growth.
Structure of the Growth Zone During Oscillatory Growth
As pollen tubes elongate, they become organized into a nonlinear dynamic system with sinusoidal oscillations of growth rates. Structural details of the subapical growth zone were closely examined in tobacco and lily pollen tubes undergoing oscillatory growth. In all pollen tubes examined, this subapical zone was clearly the region that undergoes elongation during growth (n = 40 for tobacco pollen, n = 18 for lily pollen). Vesicle incorporation into the plasma membrane in this zone is observed as highly active membrane flux followed by expansion of the cell perimeter (see also and 5, Movie 1 and 2).Citation8 Three successive growth cycles of a tobacco pollen tube are shown in . Deposition of new materials, including membrane and cell wall, at the cell surface in the growth zone causes structural disturbances that are extremely rapid and transient, and that appear as transecting features within the growth zone annulus (, arrowheads). During the growth pulse, vesicle incorporation in the growth zone supports expansion of the annulus as osmotic pressure drives cell elongation ( arrows) (see also Fig. 5 and Movie 2).Citation8
Significantly, the subapical growth zone is located at the boundary between esterified and deesterified pectins.Citation9–Citation10 Newly deposited pectins are methylesters and are enriched in the apical dome. They are deesterified by pectin methylesterase in a process that promotes cell wall rigidification. Deesterified pectins are enriched along the shank of the pollen tube. Fine-mapping of plasma membrane dynamics in growing tobacco pollen tubes showed that membrane undergoes anterograde flow from the subapical growth zone toward the apex, where membrane internalization via smooth vesicle endocytosis occurred.Citation8 There was only a low level of retrograde membrane flow from the subapical growth zone along the shank of the tube. This indicates that, at least in part, the flow of membrane and materials from the site of exocytosis controls sequestration of methylesterified pectins at the apical dome.
Cell surface disturbance at the growth zone in tobacco pollen tubes starts before the start of the growth cycle with an average of 3.44 ± 0.39 s (n = 21 cycles from 4 pollen tubes; min = 0.90 s, max = 8.40 s; growth cycle period 50–100 s) (). Similar results were observed in the structural details of lily pollen tube growth, although the cell surface features were more difficult to discern than in tobacco pollen (). Disturbance of the cell surface in lily pollen tubes precedes the start of the growth cycle by 1.02 ± 0.01 s (n = 22 cycles from 4 pollen tubes; min = 0.20 s, max = 2.20 s; growth cycle period 15–30 s) ().
Growth Reorientation Involves a Shift of the Growth Zone Axis
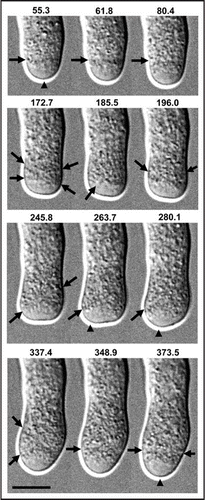
Pollen tubes can undergo abrupt interruptions of normal growth dynamics. This results in a decreased growth rate and a transient swelling of the apical region. Close examination of such an event reveals that the subapical growth zone shifts position and angle during growth reorientation (). Before the event, the subapical growth zone is organized along a transecting axis (, 55.3, 61.8, 80.4 s, arrows). During onset of the event, vesicle incorporation briefly continues but cell elongation ceases, so the tip region undergoes expansion. As the growth zone expands radially, tangential axes with different orientations emerge rapidly and transiently (, 172.7, 185.5 s, arrows). One of these axes eventually becomes stabilized and enables vesicle incorporation leading to continued cell elongation (, 245.8, 263.7, 280.1 s, arrows). As growth continues, the subapical growth zone axis is re-established (, 373.5 s, arrows).
Conclusions and Perspectives
Pollen tube growth was visualized with unprecedented clarity using refraction-free imaging and high-resolution DIC microscopy with millisecond capture.Citation8 Together with pulse-chase FM dye labeling, the results have revealed that exocytosis occurs in the subapical region. Deposition of new membrane and immature wall materials in the growth zone disturbs the cell surface and occurs before the start of the growth cycle (), indicating that other factors in addition to cell wall loosening mediate cell elongation. Recent work has demonstrated that transcellular hydrodynamic flow has an integral role in regulating rates of exocytosis and endocytosis, and driving cell elongation during growth.Citation6–Citation8
Figures and Tables
Figure 1 High-resolution refraction-free DIC microscopy of tobacco pollen tube growth. (A) Pollen germination with exocytosis along the apical perimeter (arrow). (B) Pollen tube ca. 60 µm length with exocytosis in the subapical growth zone annulus (arrow). (C) Three successive growth cycles in a pollen tube undergoing oscillatory growth. Each row represents 1 cycle with elapsed time in s given above. Arrowheads mark surface features that appear before the start of the cycle. Exocytosis and cell elongation occur in the subapical growth zone marked with arrows. (D) Oscillatory growth in a lily pollen tube, elapsed time in s given above. Growth occurs in the structurally fluid subapical zone marked with arrow. (E and F) Correlation between growth rate oscillations and disruption of cell surface features resulting from deposition of new materials. Emergence of the growth zone annulus can occur immediately before (triple arrow) or just before (double arrow) the start of the growth cycle or at the end of the previous cycle (single arrow). (E) Tobacco pollen tube, shown in (C). (F) Lily pollen tube, shown in (D). Bars = 10 µm.
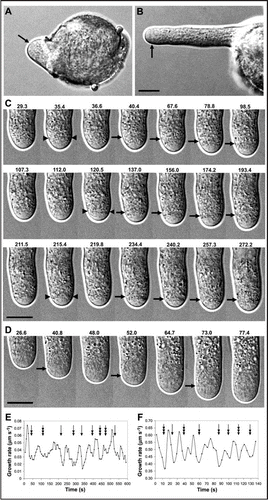
Figure 2 High-resolution refraction-free DIC microscopy of tobacco pollen tube undergoing growth reorientation. Elapsed time in s given above. The growth zone axis is reorganized from transverse (55.3–80.4 s) to tangential (245.8–289.1 s) and back to transverse (348.9–373.5 s). Arrows mark growth axis orientation. Arrowheads mark location of the tip. Bar = 10 µm.
Acknowledgements
This work is supported by the Netherlands Organization for Scientific Research (NWO-ECHO 700.56.00 and NWO-VIDI 864.05.001).
Addendum to:
References
- Holdaway Clarke TL, Hepler PK. Control of pollen tube growth: role of ion gradients and fluxes. New Phytol 2003; 159:539 - 563
- Shaw SL, Dumais J, Long SR. Cell surface expansion in polarly growing root hairs of Medicago truncatula. Plant Phys 2000; 124:959 - 969
- Parton RM, Fischer Parton S, Watahiki MK, Trewavas AJ. Dynamics of the apical vesicle accumulation and the rate of growth are related in individual pollen tubes. J Cell Sci 2001; 114:2685 - 2695
- Zonia L, Cordeiro S, Tupy J, Feijo J. Oscillatory chloride efflux at the pollen tube apex has a role in growth and cell volume regulation and is targeted by inositol 3,4,5,6-tetrakisphosphate. Plant Cell 2002; 14:2233 - 2249
- Zonia Munnik T. Osmotically induced cell swelling versus cell shrinking elicits specific changes in phospholipid signals in tobacco pollen tubes. Plant Phys 2004; 134:813 - 823
- Zonia L, Müller M, Munnik T. Hydrodynamics and cell volume oscillations in the pollen tube apical region are integral components of the biomechanics of Nicotiana tabacum pollen tube growth. Cell Biochem Biophys 2006; 46:209 - 232
- Zonia L, Munnik T. Life under pressure: hydrostatic pressure in cell growth and function. Trends Plant Sci 2007; 12:90 - 97
- Zonia L, Munnik T. Vesicle trafficking dynamics and visualization of zones of exocytosis and endocytosis in tobacco pollen tubes. J Exp Bot 2008; 59:861 - 873
- Bosch M, Cheung AY, Hepler PK. Pectin methylesterase, a regulator of pollen tube growth. Plant Physiol 2005; 138:1334 - 1346
- Bosch M, Hepler PK. Pectin methylesterase and pectin dynamics in pollen tubes. Plant Cell 2005; 17:3219 - 3226