Abstract
The interaction between tomato plants and Clavibacter michiganensis subsp. michiganensis (Cmm) represents a model pathosystem to study the interplay between the virulence determinants of a Gram-positive bacterium and the attempt of a crop plant to counteract pathogen invasion. To investigate plant responses activated during this compatible interaction, we recently analyzed gene expression profiles of tomato stems infected with Cmm. This analysis revealed activation of basal defense responses that are typically observed upon plant perception of pathogen-associated molecular patterns. In addition, Cmm infection upregulated the expression of host genes related to ethylene synthesis and response. Further analysis of tomato plants impaired in ethylene perception and production demonstrated an important role for ethylene in the development of disease symptoms. Here we discuss possible molecular strategies used by the plant to recognize Cmm infection and possible mechanisms employed by the pathogen to interfere with the activation of plant defense responses and promote disease.
Addendum to: Balaji V, Mayrose M, Sherf O, Jacob-Hirsch J, Eichenlaub R, Iraki N, Manulis-Sasson S, Rechavi G, Barash I, Sessa G. Tomato transcriptional changes in response to Clavibacter michiganensis subsp. michiganensis reveal a role for ethylene in disease development. Plant Physiol 2008; In press.
Little is known on the strategies employed by Gram-positive phytopathogenic bacteria to sense the presence of the host plant, penetrate and colonize tissue, and counteract plant defense responses. Also largely unexplored are the molecular mechanisms associated with detection of Gram-positive bacteria by the host plant and with the activation of attempted defense responses.
Among the most devastating Gram-positive disease agents are actinobacteria of the genus Clavibacter whose subspecies cause systemic infections of the xylem in different plant species.Citation1 The subspecies Clavibacter michiganensis subsp. michiganensis (Cmm) causes bacterial wilt and canker of tomato (Solanum lycopersicum), an economically important disease causing yield losses worldwide.Citation1 In recent years important insight into the molecular mechanism of Cmm pathogenicity has been achieved,Citation1 and genome sequence of a Cmm strain has been established.Citation2 Major Cmm pathogenicity determinants are plasmid borne and include the β-1,4-endocellulase CelA,Citation3 and the putative serine protease Pat-1.Citation4 Additional genes important for virulence are located in a pathogenicity island of about 129 kb on the Cmm chromosome which has a relatively low G + C content and is required for effective Cmm colonization of tomato plants.Citation2
Tomato is an economically important crop amenable to genetic analysis and transformations. Many resources are available for this plant species, including germplasm collections, natural and induced mutants, an extensive expressed sequence tag database and an ongoing genome sequencing project.Citation5 In addition, because of its experimental tractability, tomato plants have been widely used to study plant disease resistance and susceptibility. As genetic and molecular tools for both Cmm and tomato are in place, the tomato-Cmm pathosystem represents an excellent model to study the interplay between virulence determinants of a Gram-positive phytopathogenic bacterium and defense responses of a crop plant.
To get insight into host responses occurring during the tomato-Cmm compatible interaction and molecular mechanisms associated with the development of wilt and canker disease symptoms, we recently analyzed gene expression profiles of tomato stems infected with Cmm.Citation6 This analysis revealed a clear activation of basal defense responses, which are typically observed upon plant perception of pathogen-associated molecular patterns (PAMPs).Citation7 These include production and scavenging of free oxygen radicals, induction of defense-related genes, enhanced protein turnover, and hormone biosynthesis. Interestingly, several tomato genes encoding proteins with characteristics of cell-surface receptors were differentially expressed in response to Cmm infection.Citation6 These proteins can be considered as candidate receptors for Cmm PAMPs and include two receptor-like kinases, a homolog of the receptor for the fungal PAMP ethylene-inducing xylanase from Trichoderma viride,Citation8 and the Ve1 resistance protein, which confers resistance in tomato to the vascular disease Verticillium wilt.Citation9
It remains to be elucidated what are the Cmm PAMPs perceived by tomato plants. Cold-shock protein from Gram-positive bacteria and different microbial patterns of Gram-negative bacteria, including lipopolysaccharides, flagellin, and the translational elongation factor EF-TU, were shown to act as PAMPs in plants.Citation10 Similarly, Cmm cold shock protein or cell wall components, such as peptidoglycan, lipoteichoic acid, and lipopeptides, which function as Gram positive-derived PAMPs in animal systemsCitation11, may act as PAMPs during the tomato-Cmm interaction. Additional possible Cmm PAMPs are exopolysaccharides, which are produced in large amounts by the bacterium and may interact directly with surface-exposed plant proteins.Citation1 The numerous extracellular cell wall degrading enzymes secreted by Cmm may also function as PAMPs, as observed for the fungal ethylene-inducing xylanase.Citation2,Citation12 Alternatively, by virtue of their hydrolytic activity, these enzymes may release plant cell wall fragments that are recognized by PAMP receptors. Indeed, different β-glucan fragments released from plant cell walls were shown to elicit plant basal defense responses.Citation13,Citation14
How Cmm copes with the activation of basal defense responses is largely unknown. Many potential virulence determinants that might interfere with the plant defense reaction are clustered in the Cmm pathogenicity island, which is essential for effective plant colonization.Citation2 Several extracellular serine proteases are encoded in this region and inactivation of part of them by gene replacement drastically reduced Cmm colonization of tomato plants.Citation2 Although their targets are still unknown, these proteins might interfere with plant signaling pathways as it was described for certain cysteine proteases that serves in Gram-negative bacteria as suppressors of plant defenses.Citation15 An additional candidate for interference with plant signaling may be a tomatinase, also encoded in the Cmm pathogenicity island, because hydrolysis products of α-tomatine were shown to suppress plant defense responses in a fungal system.Citation16
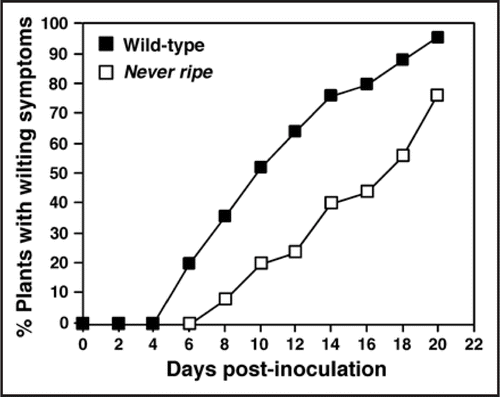
In addition to detecting the activation of basal defense responses, host gene expression profiling during the tomato-Cmm interaction unraveled the involvement of ethylene in disease development.Citation6 In fact, Cmm infection of tomato stems was found to induce expression of host genes related to ethylene biosynthesis and response ().Citation6 Further analysis of ethylene-insensitive Never ripe mutants and transgenic plants with reduced ethylene synthesis indicated that ethylene is required for normal development of wilting symptoms (), but not for the activation of defense-related genes or bacterial colonization.Citation6 We hypothesize that during infection ethylene synthesis and response are manipulated by Cmm virulence determinants to promote disease. Alternatively, ethylene is released as part of the host responses activated by bacterial recognition, or as a result of tissue maceration. In line with our first hypothesis, the type III effectors AvrPto and AvrPtoB from Pseudomonas syringae pv. tomato were shown to promote enhanced disease symptoms in tomato leaves, in part, by upregulating genes involved in ethylene production.Citation17 Interestingly, expression in tomato plants of AvrPto or AvrPtoB, and infection with Cmm resulted in the upregulation of the SlACO1 gene encoding the key enzyme of ethylene biosynthesis ACC oxidase.Citation6,Citation17
In conclusion, future research challenges for understanding how host responses are regulated by the plant and manipulated by a Gram-positive bacterium will be the isolation of Cmm PAMPs and their plant receptors, the identification of Cmm virulence determinants and the elucidation of their mode of action.
Abbreviations
| Cmm | = | Clavibacter michiganensis subsp. michiganensis |
| PAMP | = | pathogen-associated molecular pattern |
| ACC | = | 1-aminocyclopropane-1-carboxylic acid |
| ACO | = | ACC oxidase |
Figures and Tables
Figure 1 Kinetics of ACC oxidase (ACO) gene expression in tomato plants inoculated with Cmm. Six-week-old tomato plants were infected with a Cmm suspension (108 cfu/ml) or mock-inoculated. Total RNA was extracted from stem samples harvested at the indicated day post-inoculation (dpi) and subjected to Northern blot analysis using as probe a 550 bp fragment of the SlACO1 gene, which shares high homology with other ACO family members (upper). Ethidium bromide staining shows the amount of RNA loaded in each lane (lower).
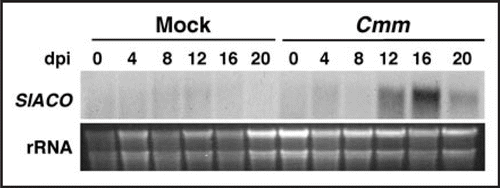
Figure 2 Effect of impaired ethylene sensitivity on development of wilt symptoms in tomato plants infected with Cmm. Six-week-old plants were infected with a Cmm suspension (108 cfu/ml) and examined for development of wilt symptoms during a 20-day period. The percentage of plants showing wilt symptoms was calculated in a group of at least 30 plants for the ethylene-insensitive mutant Never ripe and wild-type Pearson plants. Data are representative of two independent experiments.
Acknowledgements
This work was supported by the German Research Foundation program for trilateral cooperation among Israel, Palestinian Authority, and Germany (grant no. EI 535/12-1), and by the US-Israel Binational Agricultural Research and Development Fund (BARD; grant no. IS-4047-07).
Addendum to:
References
- Gartemann KH, Kirchner O, Engemann J, Grafen I, Eichenlaub R, Burger A. Clavibacter michiganensis subsp. michiganensis: first steps in the understanding of virulence of a Gram-positive phytopathogenic bacterium. J Biotechnol 2003; 106:179 - 191
- Gartemann KH, Abt B, Bekel T, Burger A, Engemann J, Flugel M, Gaigalat L, Goesmann A, Grafen I, Kalinowski J, Kaup O, Kirchner O, Krause L, Linke B, McHardy A, Meyer F, Pohle S, Ruckert C, Schneiker S, Zellermann EM, Puhler A, Eichenlaub R, Kaiser O, Bartels D. The genome sequence of the tomato-pathogenic actinomycete Clavibacter michiganensis subsp. michiganensis NCPPB382 reveals a large island involved in pathogenicity. J Bacteriol 2008; 190:2138 - 2149
- Jahr H, Dreier J, Meletzus D, Bahro R, Eichenlaub R. The endo-β-1,4-glucanase CelA of Clavibacter michiganensis subsp. michiganensis is a pathogenicity determinant required for induction of bacterial wilt of tomato. Mol Plant Microbe Interact 2000; 13:703 - 714
- Dreier J, Meletzus D, Eichenlaub R. Characterization of the plasmid encoded virulence region pat-1 of phytopathogenic Clavibacter michiganensis subsp. michiganensis. Mol Plant Microbe Interact 1997; 10:195 - 206
- Van der Hoeven R, Ronning C, Giovannoni J, Martin G, Tanksley S. Deductions about the number, organization, and evolution of genes in the tomato genome based on analysis of a large expressed sequence tag collection and selective genomic sequencing. Plant Cell 2002; 14:1441 - 1456
- Balaji V, Mayrose M, Sherf O, Jacob-Hirsch J, Eichenlaub R, Iraki N, Manulis-Sasson S, Rechavi G, Barash I, Sessa G. Tomato transcriptional changes in response to Clavibacter michiganensis subsp. michiganensis reveal a role for ethylene in disease development. Plant Physiol 2008; 146:1797 - 1809
- Nurnberger T, Brunner F, Kemmerling B, Piater L. Innate immunity in plants and animals: striking similarities and obvious differences. Immunol Rev 2004; 198:249 - 266
- Ron M, Avni A. The receptor for the fungal elicitor ethylene-inducing xylanase is a member of a resistance-like gene family in tomato. Plant Cell 2004; 16:1604 - 1615
- Kawchuk LM, Hachey J, Lynch DR, Kulcsar F, van Rooijen G, Waterer DR, Robertson A, Kokko E, Byers R, Howard RJ, Fischer R, Prufer D. Tomato Ve disease resistance genes encode cell surface-like receptors. Proc Natl Acad Sci USA 2001; 98:6511 - 6515
- Bittel P, Robatzek S. Microbe-associated molecular patterns (MAMPs) probe plant immunity. Curr Opin Plant Biol 2007; 10:335 - 341
- Aderem A, Ulevitch RJ. Toll-like receptors in the induction of the innate immune response. Nature 2000; 406:782 - 787
- Belien T, Van Campenhout S, Robben J, Volckaert G. Microbial endoxylanases: effective weapons to breach the plant cell-wall barrier or, rather, triggers of plant defense systems?. Mol Plant Microbe Interact 2006; 19:1072 - 1081
- Klarzynski O, Plesse B, Joubert JM, Yvin JC, Kopp M, Kloareg B, Fritig B. Linear beta-1,3 glucans are elicitors of defense responses in tobacco. Plant Physiol 2000; 124:1027 - 1038
- Yamaguchi T, Yamada A, Hong N, Ogawa T, Ishii T, Shibuya N. Differences in the recognition of glucan elicitor signals between rice and soybean: beta-glucan fragments from the rice blast disease fungus Pyricularia oryzae that elicit phytoalexin biosynthesis in suspension-cultured rice cells. Plant Cell 2000; 12:817 - 826
- Mudgett MB. New insights to the function of phytopathogenic bacterial type III effectors in plants. Annu Rev Plant Biol 2005; 56:509 - 531
- Bouarab K, Melton R, Peart J, Baulcombe D, Osbourn A. A saponin-detoxifying enzyme mediates suppression of plant defences. Nature 2002; 418:889 - 892
- Cohn JR, Martin GB. Pseudomonas syringae pv. tomato type III effectors AvrPto and AvrPtoB promote ethylene-dependent cell death in tomato. Plant J 2005; 44:139 - 154