Abstract
The signal transduction pathway of the plant stress and defense hormone, ethylene, has been extensively elucidated using the plant genetic model Arabidopsis over the last two decades. Among others, a MAPKKK CTR1 was identified as a negative regulator that has led to the speculation of MAPK involvement in ethylene signaling. However, it remained unclear how the MAPK modules acting downstream of the receptors to mediate ethylene signaling We have recently presented new evidence that the MKK9-MPK3/6 modules identified by combined functional genomic and genetic screens mediate ethylene signaling, which is negatively regulated by the genetically identified CTR1-dependent cascades. Our genetic studies show consistently that the MKK9-MPK3/MPK6 modules act downstream of the ethylene receptors. Biochemical and transgenic analyses further demonstrated that the positive-acting and negative-acting MAPK activities are integrated and act simultaneously to control the key transcription factor EIN3 through dual phosphorylations to regulate the EIN3 protein stability and downstream transcription cascades. This study has revealed a novel molecular mechanism that defines the specificity of complex MAPK signaling. Comprehensive elucidation of MAPK cascades and the underlying molecular mechanisms would provide more precise explanations for how plant cells utilize MAPK cascades to control specific downstream outputs in response to distinct stimuli.
Addendum to: Yoo SD, Cho YH, Tena G, Xiong Y, Sheen J. Dual Control of nuclear EIN3 by bifurcate MAPK cascades in C2H4 signalling. Nature 2008; 451:789.
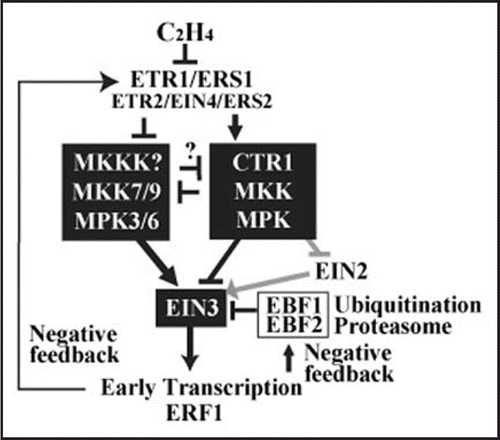
Ethylene (C2H4) regulates stress and defense responses and many key events of plant growth and development.Citation1,Citation2 In the plant model system Arabidopsis thaliana, several signaling components in the ethylene signal transduction pathway have been genetically identified from membrane receptors to nuclear activators as well as feedback regulators ().Citation2,Citation3 Multiple membrane proteins, ETR1, ETR2, ERS1, ERS2 and EIN4, play partially redundant roles as ethylene receptors.Citation4–Citation9 In the absence of ethylene, ETR1 and other receptors suppress hormone signaling by activating a negative regulator, CTR1Citation10,Citation11 (a putative Raf-like MAPKKK), in ER complexes.Citation12–Citation15 Subsequently, a key transcription factor EIN3 is degraded by the 26S proteasome through the recognition by specific F-box proteins EBF1/2 in the E3 ligase complexes.Citation16–Citation20 Such action eventually blocks the downstream signal responses.
In contrast, upon binding of ethylene to the receptors, CTR1 is dissociated from receptor complexes and inactivated to initiate the downstream signaling.Citation15 The immediately following principal nuclear event is to stabilize EIN3 proteins in the nucleus to activate the primary transcription.Citation16–Citation21 In ethylene signaling, a metal transporter-like membrane protein EIN2, that is genetically placed downstream of the receptors but upstream of EIN3, is to be important for the EIN3 accumulation.Citation18,Citation22 One of the primary EIN3 response genes with EIN3 binding sites in its promoter region, ERF1, is involved in the secondary transcription activation through its GCC element binding activity.Citation23 There are several negative feedback mechanisms. For instance, ERS1 and ETR2 are transcriptionally elevated as a primary response to ethylene, and then the newly synthesized ethylene receptors attenuate the signaling effect of ethylene.Citation6,Citation8 EBF2 is also transcriptionally activated in early ethylene signaling and destabilizes EIN3 in the nucleus and diminish its accumulation.Citation18
Despite the well-defined genetic pathway, cellular, molecular and biochemical connections among individual components remain to be elucidated in ethylene signaling. For example, an elevated level of MAPK-like activities have been reported in the loss-of-function ctr1 mutant, strongly implicating positively acting MAPKs are involved in ethylene signaling ().Citation24,Citation25 However, the MAPK cascade components have not been unequivocally identified.Citation26
Here we have elucidated MAPK cascades in ethylene signaling using an integrative approach combining molecular, cellular, computational and genetic tools via exploring genomic information available in Arabidopsis.Citation27,Citation28 First, cell-based MAPK activity screen and ethylene-specific reporter assay facilitated by constitutively ethylene responsive ctr1 cells indicated the activity of antagonistic MAPK cascades in ethylene signaling: MKK9-MPK3/6 comprises positive-acting MAPK cascades, whereas CTR1 initiates negative-acting ones. Consistently, loss-of-function mkk9 shows a broad spectrum of ethylene insensitivity for the typical triple response, primary gene activation, ethylene-dependent growth inhibition and senescence promotion, as well as hypersensitivity to glucose and salt. The epistatic analysis using a transgenic approach indicates that MKK9 act downstream of the receptor complexes, but upstream or independent of EIN2. The MKK9 localization in the nucleus upon signaling as well as the MAPK cascade dependent EIN3 regulation in the nucleus have provided compelling evidence that two antagonistic MAPK cascades activities are integrated into regulating EIN3 through alternative phosphorylation, and modulating the protein stability and downstream transcription cascades. Significantly, this study establishes a new paradigm for linking complex MAPK cascades in controlling quantitative hormonal responses.
Since several hormone, stress and defense signals can activate MPK3 and MPK6 through upstream MAPK cascades in plants, it has long been questioned how converged MAPK signaling can secure their specificity. In this study, we have demonstrated that ethylene signaling specifically activates the MKK9-MPK3/6 modules that phosphorylate T174 of EIN3 and stabilize the EIN3 protein, but suppresses the CTR1-dependent cascades phosphorylating T592 of EIN3 that enhances the protein degradation. Only when both MAPK modules are regulated simultaneously, ethylene signaling can be appropriately executed in plants. This explains the broad but relatively weak ethylene insensitivity of mkk9 lacking only one part of two MAPK cascades involved in ethylene signaling. Likewise, mkk9 ctr1 double mutants displayed a partial but clear ethylene insensitivity in light-grown seedlings. Moreover, ctr1 displays a stronger constitutive ethylene signaling phenotype most likely due to the activation or derepression of the MKK9-MPK3/6 cascade in addition to the complete loss of CTR1-dependent MAPK cascade activity.
How CTR1 regulates the MKK9-MPK3/6 modules and which MKKs and MPKs are involved in the CTR1 modules remain to be determined. In our preliminary studies, there appears to be multiple MKKs sharing the activity of CTR1 in suppressing ethylene-specific reporter expression and promoting EIN3 degradation. Some loss- of-function mkk mutants exhibit ethylene hypersensitivity. The new studies have established essential cellular and genetics tools and assays as well as a novel conceptual foundation for more detailed molecular understanding of ethylene signaling. Future efforts will elucidate the complete and complex MAPK cascades in ethylene signaling as well as in other stress, defense and hormone signaling pathways.
Abbreviations
| MAPK | = | Mitogen activated protein kinase |
| EIN3 | = | ethylene insensitive3 |
| CTR1 | = | constitutive triple response1 |
| MKK | = | MAPK kinase |
| MKKK | = | MAPKK kinase |
Figures and Tables
Figure 1 Model of antagonistic MAPK cascades in ethylene signaling. A hypothetical MKKK is placed upstream of MKK7/9.
Addendum to:
References
- Abeles FB, Morgan PW, Saltveit JME. Ethylene in Plant Biology 1992; 2nd edn San Diego Academic Press
- Schaller GE, Kieber JJ. Somerville C, Meyerowitz EM. Ethylene. The Arabidopsis Book 2002; Rockville, MD American Society of Plant Biologists Vol http://dx.doi.org/10.1199/tab.0071
- Alonso JM, Stepanova AN. The ethylene signalling pathway. Science 2004; 306:1513 - 1515
- Bleecker AB, Estelle MA, Somerville C, Kende H. Insensitive to ethylene conferred by a dominant mutation in Arabidopsis thaliana. Science 1988; 241:1086 - 1089
- Chang C, Kwok SF, Bleecker AB, Meyerowitz EM. Arabidopsis ethylene response gene ETR1: Similarity of product to two component regulators. Science 1993; 262:539 - 544
- Hua J, Chang C, Sun Q, Meyerowitz EM. Ethylene sensitivity conferred by Arabidopsis ERS gene. Science 1995; 269:1712 - 1714
- Hua J, Sakai H, Nourizadeh S, Chen QG, Bleecker AB, Ecker JR, Meyerowitz EM. EIN4 and ERS2 are members of the putative ethylene receptor family in Arabidopsis. Plant Cell 1998; 10:1321 - 1332
- Sakai H, Hua J, Chen QG, Chang C, Medrano LJ, Bleecker AB, Meyerowitz EM. ETR2 is an ETR1-like gene involved in ethylene signaling in Arabidopsis. Proc Natl Acad Sci USA 1998; 95:5812 - 5817
- Schaller GE, Bleecker AB. Ethylene-binding sites generated in yeast expressing the Arabidopsis ETR1 gene. Science 1995; 270:1809 - 1811
- Hua J, Meyerowitz EM. Ethylene responses are negatively regulated by a receptor gene family in Arabidopsis thaliana. Cell 1998; 94:261 - 271
- Wang W, Hall AE, O'Malley R, Bleecker AB. Canonical histidine kinase activity of the transmitter domain of the ETR1 ethylene receptor from Arabidopsis is not required for signal transmission. Proc Natl Acad Sci USA 2003; 100:352 - 357
- Kieber JJ, Rothenberg M, Roman G, Feldmann KA, Ecker JR. CTR1, a negative regulator of the ethylene response pathway in Arabidopsis, encodes a member of the raf family of protein kinases. Cell 1993; 72:427 - 441
- Clark KL, Larsen PB, Wang X, Chang C. Association of the Arabidopsis CTR1 raflike kinase with the ETR1 and ERS ethylene receptors. Proc Natl Acad Sci USA 1998; 95:5401 - 5406
- Huang Y, Li H, Hutchison CE, Laskey J, Kieber JJ. Biochemical and functional analysis of CTR1, a protein kinase that negatively regulates ethylene signaling in Arabidopsis. Plant J 2003; 33:221 - 233
- Gao Z, Chen YF, Randlett MD, Zhao XC, Findell JL, Kieber JJ, Schaller GE. Localization of the Raf-like kinase CTR1 to the endoplasmic reticulum of Arabidopsis through participation in ethylene receptor signaling complexes. J Biol Chem 2003; 278:34725 - 34732
- Chao Q, Rothenberg M, Solano R, Roman G, Terzaghi W, Ecker JR. Activation of the ethylene gas response pathway in Arabidopsis by the nuclear protein Ethylene-Insensitive3 and related proteins. Cell 1997; 89:1133 - 1144
- Yanagisawa S, Yoo SD, Sheen J. Differential regulation of EIN3 stability by glucose and ethylene signalling in plants. Nature 2003; 425:521 - 525
- Guo H, Ecker JR. Plant responses to ethylene gas are mediated by SCFEBF1/EBF2-dependent proteolysis of EIN3 transcription factor. Cell 2003; 115:667 - 677
- Potuschak T, Lechner E, Parmentier Y, Yanagisawa S, Grava S, Koncz C, Genschik P. EIN3-dependent regulation of plant ethylene hormone signaling by two Arabidopsis F box proteins: EBF1 and EBF2. Cell 2003; 115:679 - 689
- Gagne JM, Smalle J, Gingerich DJ, Walker JM, Yoo SD, Yanagisawa S, Vierstra R. Arabidopsis EIN3-binding F-box 1 and 2 form ubiquitin-protein ligases that repress ethylene action and promote growth by directing EIN3 degradation. Proc Natl Acad Sci USA 2004; 101:6803 - 6808
- Binder BM, Walker JM, Gagne JM, Emborg TJ, Hemmann G, Bleecker AB, Vierstra RD. The Arabidopsis EIN3 binding F-Box proteins EBF1 and EBF2 have distinct but overlapping roles in ethylene signaling. Plant Cell 2007; 19:509 - 523
- Alonso JM, Hirayama T, Roman G, Nourizadeh S, Ecker JR. EIN2, a bifunctional transducer of ethylene and stress responses in Arabidopsis. Science 1999; 284:2148 - 21452
- Solano R, Stepanova A, Chao Q, Ecker JR. Nuclear events in ethylene signaling: a transcriptional cascade mediated by ETHYLENE-INSENSITVE3 and ETHYLENE-ESPONSE-FACTOR1. Genes Dev 1998; 12:3703 - 3714
- Novikova GV, Moshkov IE, Smith AR, Hall MA. The effect of ethylene on MAP Kinase-like activity in Arabidopsis thaliana. FEBS 2000; 474:29 - 32
- Ouaked F, Rozhon W, Lecourieux D, Hirt H. A MAPK pathway mediates ethylene signaling in plants. EMBO J 2003; 22:1282 - 1288
- Ecker JR. Reentry of the ethylene MPK6 module. Plant Cell 2004; 16:3169 - 3173
- Yoo SD, Cho YH, Xiong Y, Tena G, Xiong Y, Sheen J. Dual control of nuclear EIN3 by bifurcated MAPK cascades in C2H4 signalling. Nature 2008; 451:789 - 795
- Yoo SD, Cho YH, Sheen J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nature Protocols 2007; 2:1565 - 1572