Abstract
Lobelia dortmanna leads a group of small, highly-valued rosette species that grow on coarse, nutrient-poor soils in temperate softwater lakes. They acquire most CO2 for photosynthesis by root uptake and efficient gas transport in large air channels to the leaves. Lobelia is the only species that releases virtually all photosynthetic oxygen from the roots and generates profound day-night changes in oxygen and CO2 in the sediment pore-water. While oxygen release from roots stimulates decomposition and supports VA-mycorrhiza fungi, the ready gas exchange presents a risk of insufficient oxygen supply to the distal root meristems as sediments accumulate organic matter from lake pollution. So the plant with the greatest oxygen release from roots is also the most sensitive to oxygen depletion in sediments and it dies or losses anchorage by shortening the roots from 10 to 2 cm at even modest contents (2.4%) of degradable organic matter. Coatings of oxidized iron on roots in organically enriched sediments reduce radial oxygen loss and, thereby, increase internal concentrations and supply of oxygen to root tips. Oxidized iron is also a redox buffers which may prevent the ingress of sulfides and other reduced toxic solutes during nights. Controlled experiments are under way to test if iron enrichment can help survival of rosette species threatened by lake pollution or whether removal of organic surface sediments is required.
Addendum to: Møller CL, Sand-Jensen K. Iron plaques improve the oxygen supply to root meristems of the freshwater plant, Lobelia dortmanna. New Phytol 2008; In press.
Inhospitable Sediments
Aquatic sediments and waterlogged soils are usually anoxic a few mm below the surface.Citation1,Citation2 Aquatic and wetland plants, therefore, need to supply oxygen to the roots by rapid intra-plant gas transport through large air channels running from the green shoot in contact with atmospheric air or aerated water to the roots deeply buried in anoxic environments.Citation3,Citation4 To ensure oxygen transport to the root tip in competition with radial oxygen loss to the anoxic hydrosoil, it should be advantageous for roots to be thick, short and impermeable to radial oxygen loss.Citation5,Citation6 Thick roots, however, are costly to produce relative to their capacity to take up nutrients so this morphology is not a general solution to cope with anoxic hydrosoils. Short roots, likewise, have a reduced nutrient uptake capacity and also a poor anchorage. Thus, the third option—having roots relatively impermeable to radial oxygen loss—appears to be a suitable solution. Indeed, this property is common among wetland plants,Citation6 it is present in the marine seagrass, Zostera marina and probably widespread among marine and freshwater plants ().
Small freshwater rosette species, however, have highly gas permeable root surfaces as an adaptation to take up free CO2 for photosynthesis from the CO2-rich hydrosoil in the nutrient-poor sandy sediments of softwater lakes.Citation1,Citation7 These rosette species have thick entire leaves from a short stem, many roots and well-developed air channels facilitating transport of oxygen and CO2 between leaves and roots. Utilizing CO2 from the hydrosoil for photosynthesis, the roots are also highly permeable to radial oxygen transportCitation8 and this property has recently been confirmed by quantification of oxygen loss and root wall permeability for every 5 mm along the roots of Lobelia dortmanna.Citation9 The question therefore arises: how do these species supply oxygen to root tips in sediments undergoing organic enrichment and oxygen depletion?
Coping with Sediment Oxygen Depletion
Lobelia dortmanna is the only known species that releases virtually all photosynthetic oxygen from the roots to the sediments thanks to high root surface permeability and low leaf surface permeability and it, therefore, generates profound day-night fluctuations in pools and penetration depth of oxygen in sandy sediments of low oxygen consumption.Citation1,Citation2 Sediment CO2 varies opposite to oxygen due to their complementary roles in photosynthesis and respiration.Citation1 Other rosette species also exchange much CO2 and oxygen via the roots,Citation8 but leaf exchange is more important than in Lobelia.
The ready oxygen exchange across Lobelia roots and the oxic conditions in nutrient-poor sediments have been regarded as an advantage for the nutrient supply due to stimulation of organic decomposition by aerobic sediment bacteria and mycorrhiza fungi.Citation1 This situation is reversed, however, in sediments undergoing organic enrichment by input from the catchment or from phytoplankton-rich lake water. Organically enriched sediments of higher oxygen demand offer a threat to survival of rosette species due to higher radial oxygen loss and insufficient oxygen supply to root tips during the night when photosynthetic oxygen production is switched off and the lake water is the only oxygen source to plant and sediment respiration.Citation7,Citation9
Organic enrichment of sandy Lobelia sediments leads to a decline in root length from about 10 cm to 2 cm (). While growth and tissue concentrations of N and P are stimulated by a small organic enrichment (e.g., from 0.3 to 0.6% of dry weight) due to higher mineralization, a stronger organic enrichment (2.4%) stresses the plants and leads to loss of chlorophyll, poor growth and die offCitation4. Organic enrichment is accompanied by deprivation of oxygen pools and lower oxygen penetration depths (e.g., from 35 to 5 mm) in the sediments signaling that insufficient oxygen supply to the root tips is a main reason for the reduction of root length and plant performance.Citation4 Quantity and degradability of the organic matter input will obviously determine the severity of plant stress.Citation6
Other rosette species such as Littorella uniflora and Isoetes lacustris respond similarly but are less sensitive to sediment anoxia presumably due to lower radial oxygen loss from the roots. Littorella produces a mixture of thin, short roots and thick, longer and probably less susceptible roots. Both species tolerate organic sediments and in deep calm water you find Isoetes lacustris anchored with just the stem in very soft organic sediments, but with no roots and an obvious risk of vegetation loss during storms or release of methane bubbles from the sediments.
Iron Plaques as a Protection?
Wetland plants and probably most aquatic plants have continuous layers of suberin or lignin just below the root surface as barriers to radial oxygen loss to the sediment.Citation6,Citation11 In Phragmites australis the barrier becomes stronger when the roots are exposed to anoxia and small fatty acids from anaerobic fermentation.Citation5 There is no sign that this diffusion barrier is present or inducible in Lobelia roots, while the response of other rosette species is unknown.Citation9 If the diffusion barrier turns out to be inducible, it will impede root uptake of CO2, but because sediment CO2 concentrations are elevated by organic enrichmentCitation4 it could match a certain reduction of root permeability and still maintain the CO2 influx for photosynthesis.
For Lobelia, alteration of bio-geochemical processes accompanying organic enrichment of the sediment can offer additional protection to radial oxygen loss from the roots and this mechanism has hitherto been overlooked. As Lobelia's roots become shorter by oxygen depletion of sediments, 1–2 mm thick coatings of oxidized iron precipitate on the root surfacesCitation4,Citation10 (). Mobile dissolved Fe2+ is produced from insoluble Fe3+ by degradation of organic matter in the anoxic sediment and Fe2+ is re-precipitated as Fe3+-oxyhydroxides at the root surface when it meets the outward oxygen flux. Iron coatings on aquatic plant roots in organically rich sediments are common and the consequences should be general.
Iron coatings on Lobelia roots (30 mmol m−2) reduce 10-fold oxygen permeability of the root wall and two-fold radial oxygen loss (). Therefore, oxygen concentrations rise within the roots and can better sustain root respiration including the distal meristem.Citation9 While iron coatings only lead to c. 15% higher internal oxygen concentrations in the root tips at low respiration rates at low temperatures,Citation9 the influence will be stronger at higher respiration rates and at reduced oxygen supply from the leaves in the dark.
Therefore, we anticipate that iron coatings should offer the strongest protection to root anoxia and other toxic effects during warm summer nights when oxygen consumption rates in the plants and the sediments peak, hence, no oxygen is produced by photosynthesis and oxygen supply from the bottom waters is reduced by falling oxygen concentrations.
Oxidized iron may offer additional protection by increasing the diffusive resistance to toxic solutes across the root surfaces and providing an oxidation capacity slowing the ingress of sulfide and reduced metals from the sediment.Citation5,Citation9 Iron coatings formed by oxygen release from the roots during the day can, thereby, offer protection during the night when oxygen release diminishes or is reversed.Citation1 Likewise, iron coatings formed during sunny days can offer protection during dark days.
Studies of marine sediments have clearly demonstrated how surface pools of oxidized iron formed in winter and spring when bottom waters are rich in oxygen can protect the anoxic bottom layer during summer against the release of toxic sulfides from deeper sediments.Citation12
To test the protective effect of oxidized iron on or close to root surfaces independent of the influence of sediment anoxia and oxygen supply from the water, we shall expose Lobelia and other freshwater species to controlled crossed gradients of degradable organic matter and iron in the sediments and set levels of dissolved oxygen in the water and evaluate the resulting morphological and functional plant responses.
Iron and other important redox constituents vary extensively in amount and input by groundwater seepage through sediments from different lakes and wetlands as do the presence and well-being of the vegetation. Thus, if oxidized iron is generally ecological significant for plant survival it will also be important for plant distribution and historical changes of the vegetation. Work is under way to evaluate the importance of light availability and sediment contents of organic matter and iron for the survival of the Red-listed rosette vegetation in the diminishing numbers of oligotrophic lakes worldwide.Citation13
Figures and Tables
Figure 1 Radial oxygen loss measured with platinum sleeve electrodes along the length of roots in an anoxic medium. The basal part of the roots was in contact with atmospheric air. Lobelia dortmanna had either none or thick iron coatings of the root surface (0.09 ± 0.05 and 30 ± 3 mmol Fe m−2 root surface respectively),Citation9 while Phragmites australisCitation5 and the seagrass, Zostera marina (Unpublished) were without iron coatings.
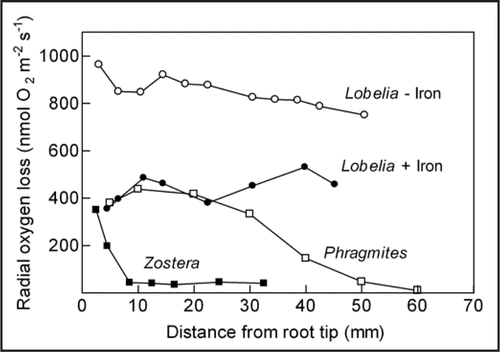
Figure 2 Roots become shorter and iron coatings thicker on Lobelia roots exposed from left to right to increasing levels of organic matter (from 0.2 to 3.4 % of dry weight) and potential oxygen consumption rates (from 1.1 to 24.0 µg O2 ml−1 sediment h−1) for 16 weeks.Citation10

Addendum to:
References
- Pedersen O, Sand-Jensen K, Revsbech NP. Diel pulses of O2 and CO2 in sandy lake sediments inhabited by Lobelia dortmanna. Ecology 1995; 76:1536 - 1545
- Sand-Jensen K, Pedersen O, Binzer T, Borum J. Contrasting oxygen dynamics in the freshwater isoetid Lobelia dortmanna and the marine seagrass Zostera marina. Ann Bot 2005; 96:613 - 623
- Justin SHFW, Armstrong W. The anatomical characteristics of roots and plant-response to soil flooding. New Phytol 1987; 106:465 - 495
- Sand-Jensen K, Borum J, Binzer T. Oxygen stress and reduced growth of Lobelia dortmanna in sandy lake sediments subject to organic enrichment. Freshwater Biol 2005; 50:1034 - 1048
- Armstrong J, Armstrong W. Rice and Phragmites: effects of organic acids on growth, root permeability and radial oxygen loss to the rhizosphere. Am J Bot 2001; 88:1359 - 1370
- Colmer TD. Long-distance transport of gases in plants: a perspective on internal aeration and radial oxygen loss from roots. Plant Cell Environm 2003; 26:17 - 36
- Sand-Jensen K, Prahl C. Oxygen exchange with the lacunae and across leaves and roots of the submerged vascular macrophyte, Lobelia dortmanna L. New Phytol 1982; 91:103 - 120
- Sand-Jensen K, Prahl C, Stokholm H. Oxygen release from roots of submerged aquatic macrophytes. Oikos 1982; 50:1034 - 1048
- Møller CL, Sand-Jensen K. Iron plaques improve the oxygen supply to root meristems of the freshwater plant, Lobelia dortmanna. New Phytol 2008; 179:848 - 856
- Raun AL. Sediment organic matter influences growth and survival of submerged plants 2008; University of Copenhagen MS. Thesis, Freshwater Biological Laboratory
- Soukup A, Armstrong W, Schreiber L, Franke R, Votrubova O. Apoplastic barriers to radial oxygen loss and solute penetration: a chemical and functional comparison of the exodermis of two wetland species, Phragmites australis and Glyceria maxima. New Phytol 2007; 173:264 - 267
- Azzoni R, Giordani G, Viaroli P. Iron-sulphur-phosphorus interactions: implications for sediment buffering capacity in a Mediterranean eutrophic lagoon (Sacca di Goro, Italy). Hydrobiologia 2005; 550:131 - 148
- Sand-Jensen K, Riis T, Vestergaard O, Larsen S. Macrophyte decline in Danish lakes and streams over the past 100 years. J Ecol 2000; 88:1030 - 1040