Abstract
Receptor-like proteins (RLPs) are cell surface receptors that play important roles in various processes. In several plant species RLPs have been found to play a role in disease resistance, including the tomato Cf and Ve proteins and the apple HcrVf proteins that mediate resistance against the fungal pathogens Cladosporium fulvum, Verticillium spp. and Venturia inaequalis, respectively. The Arabidopsis genome contains 57 AtRLP genes. Two of these, CLV2 (AtRLP10) and TMM (AtRLP17), have well-characterized functions in meristem and stomatal development, respectively, while AtRLP52 is required for defense against powdery mildew. We recently reported the assembly of a genome-wide collection of T-DNA insertion lines for the Arabidopsis AtRLP genes. This collection was functionally analyzed with respect to plant growth, development and sensitivity to various stress responses including pathogen susceptibility. Only few new phenotypes were discovered; while AtRLP41 was found to mediate abscisic acid sensitivity, AtRLP30 (and possibly AtRLP18) was found to be required for full non-host resistance to a bacterial pathogen. Possibly, identification of novel phenotypes is obscured by functional redundancy. Therefore, RNA interference (RNAi) to target the expression of multiple AtRLP genes simultaneously was employed followed by functional analysis of the RNAi lines.
Addendum to: Wang G, Ellendorff U, Kemp B, Mansfield JW, Forsyth A, Mitchell K, Bastas K, Liu CM, Woods-Tör A, Zipfel C, de Wit PJGM, Jones JDG, Tör M, Thomma BPHJ. A genome-wide functional investigation into the roles of receptor-like proteins in Arabidopsis. Plant Physiol 2008; 147:503-17.
Receptor-like proteins (RLPs) are cell surface receptors that typically consist of an extracellular leucine-rich repeat (eLRR) domain, a single-pass transmembrane domain and a short cytoplasmatic tail that lacks obvious motifs for intracellular signaling except for the putative endocytosis motif found in some members.Citation1–Citation3 In several plant species RLPs play important roles in development and pathogen defense. Arabidopsis CLAVATA2 (CLV2; AtRLP10) and its maize ortholog FASCINATED EAR2 are required for maintaining the meristematic stem cell population in shoot apical meristems, while Arabidopsis TOO MANY MOUTHS (TMM; AtRLP17) controls the initiation of stomatal precursor cells.Citation4–Citation7 The RLP disease resistance gene family comprises the tomato Cf and Ve genes that provide resistance against Cladosporium fulvum and Verticillium spp., respectively,Citation8–Citation10 LeEIX genes that encode receptors for the ethylene inducible xylanase produced by Trichoderma biocontrol fungi,Citation11 apple HcrVf genes that confer resistance to the scab fungus Venturia inaequalis,Citation12 and an Arabidopsis RLP gene (AtRLP52) that provides resistance against the powdery mildew pathogen Erysiphe cichoracearum.Citation13 We recently reported the assembly of a genome-wide collection of T-DNA insertion lines for the 57 Arabidopsis RLP genes (AtRLP) in the Arabidopsis genome.Citation14 This collection was functionally analyzed with respect to plant growth, development and sensitivity to various stress responses including pathogen susceptibility. Only few novel phenotypes were discovered; while AtRLP41 was found to mediate abscisic acid sensitivity, AtRLP30 (and possibly AtRLP18) was found to influence non-host resistance towards Pseudomonas syringae pv. phaseolicola.Citation14
The lack of identification of biological functions for the majority of the AtRLP genes may be caused by functional redundancy. Here, we describe a reverse genetics strategy by employing RNA interference (RNAi) to target the expression of multiple AtRLP genes simultaneously, and thus possibly overcome functional redundancy among AtRLP genes. To select suitable fragments for RNAi silencing, the AtRLP genes were aligned and sequence stretches of a few hundred base pairs (bp) containing minimum one 21 bp stretch with 100% identity to minimum one other AtRLP gene were identified. Specificity of the selected fragments was verified with BLAST searches against the Arabidopsis genome.Citation15 Seven AtRLP gene fragments, varying in length between 238 and 407 bp, were PCR-amplified such that the PCR products contained a 5' BamHI or HindIII site and a 3′ EcoRI and NotI site (; ) and cloned into the pGEM-T Easy vector (Promega, Leiden, The Netherlands). The resulting plasmids were digested in two separate reactions with HindIII (or BamHI for RNAi constructs 2 and 5) in combination with NotI and in combination with EcoRI. Both inserts were cleaned from gel using the QIAquick gel extraction kit (Qiagen, Venlo, NL) and subsequently ligated with a NotI- and EcoRI-digested 129 bp spacer segment from the Pichia pastoris Aox-1 gene into the HindIII-digested (or BamHI for RNAi constructs 2 and 5) pGreen plasmidCitation16 to obtain inverted repeat constructs driven by the CaMV 35S promoter that target expression of multiple AtRLP genes (). The resulting seven plasmids were introduced into Agrobacterium tumefaciens strain GV3101 by electroporation, transformed to Arabidopsis,Citation17 and multiple homozygous single-insertion T3 lines were selected on MS plates supplemented with 100 µg/mL kanamycin that were used for functional analysis.
Similar as the individual AtRLP insertion lines,Citation14 also the RNAi lines were analyzed with respect to plant development and sensitivity to various abiotic and biotic stress factors. Development of roots, rosettes, leaf cuticle and flowers as well as stomatal patterning were examined, but no developmental anomalies were observed. In addition, the RNAi lines were assayed for altered sensitivity to plant hormones and abiotic stress factors. The only consistently altered phenotype was observed for lines containing RNAi construct 5 upon exogenous application of the plant hormone abscisic acid (ABA), as leaves of the RNAi lines bleached while wild-type leaves remained green (). Since RNAi construct 5 is predicted to target AtRLP41 of which a knock-out has been shown to result in enhanced ABA susceptibilityCitation14 this phenotype was expected. Moreover, this observation confirms that RNAi-mediated gene silencing can be used as a mechanism to investigate the function of RLP receptors. To determine whether AtRLP genes play a role in recognition of plant pathogens, similar as the individual AtRLP insertion linesCitation14 the collection of AtRLP RNAi lines was assessed for altered phenotypic responses upon challenge with a range of diverse host-adapted and non-adapted necrotrophic and biotrophic pathogens.Citation14,Citation18 In addition to the previously used pathogens,Citation14 we included Fusarium oxysporum f. sp. raphani strain 815, the Verticillium dahliae strains St12.01, St17.01 and JR2, as well as the two oomycete strains Phytophthora brassicae HH/CBS782.97 and CBS686.95 in our analysis. Interestingly, no significant differences were identified when the responses of the RNAi lines were compared to those of the parental Col-0 line upon inoculation with any of the pathogens used.
The Arabidopsis genome harbors 24 loci containing a single AtRLP gene and 13 loci comprising multiple, between two and five, AtRLP genes.Citation14,Citation19 Often, the most homologous AtRLP genes reside at the same locus,Citation14,Citation19 and therefore crossing individual T-DNA insertion lines to obtain knock-out lines for multiple AtRLP genes is nearly impossible. RNAi-mediated gene silencing currently is the most suitable strategy to target expression of several highly homologous genes simultaneously. Based on the sequence comparison between Arabidopsis and rice RLP genes, and building on the hypothesis that developmental genes are less likely to be duplicated and undergo diversifying selection than are disease resistance genes,Citation20 most AtRLP genes were proposed to be candidate disease resistance genes.Citation19 Remarkably, despite an extensive list of pathogens tested, including adapted and non-adapted pathogens of Arabidopsis, we have been able to identify only one AtRLP gene with a role in basal non-host resistance against the non-adapted bacterial pathogen Pseudomonas syringae pv phaseolicola when screening a genome-wide collection of T-DNA insertion lines in the AtRLP genes.Citation14 It was hypothesized that the lack of identification of biological functions for AtRLP genes may be explained by functional redundancy.Citation14 In the experiments presented in this manuscript we employed RNA interference to interfere with the expression of multiple AtRLP genes at the same time to overcome functional redundancy among AtRLP genes. Nevertheless, no biological functions could be assigned to additional AtRLP genes. Obviously, the targeted AtRLP genes might function in defense against pathogens that have not yet been assayed. As suggested previously,Citation14 if AtRLP genes are active in non-host resistance or basal defense, the array of potential microbial targets may be significantly increased and the response to more microbes or even insects and nematodes should be tested.Citation21 Furthermore, it may be questioned whether the knock-down established by RNAi is sufficiently strong to compromise RLP receptor activity, although gene silencing has been successfully used to compromise the activity of RLP-type disease resistance genes in tomato.Citation22 Also, the observation that transformants expressing RNAi construct 5 phenocopies the AtRLP41 T-DNA insertion allele with respect to ABA responsiveness argues against this possibility. Possibly, however, the RNAi constructs do not silence all redundant AtRLP homologs as efficiently or target all the redundant AtRLP homologs. For instance, RNAi construct 4 that is derived from AtRLP15 is predicted to silence expression of AtRLP13, but not of AtRLP16 which is also close homologue of AtRLP15. Finally, redundant AtRLP genes are not necessarily those with the highest overall homology, since ligand specificity may be determined by only a small sequence stretch, making it difficult to design the most potent RNAi constructs. Therefore, a more extensive analysis using many more RNAi constructs is needed to exclude the possibility that the lack of phenotypes can be explained by a high degree of functional redundancy among the AtRLP genes. Overall, the RNAi lines developed in our studies provide a useful tool for further investigation into roles of the AtRLP genes.
Figures and Tables
Figure 1 Cloning strategy for RNAi constructs. (A) PCR fragments of specific AtRLP fragments are generated with 5′HindIII (or BamHI for RNAi construct 2 and 5) and 3′ NotI and EcoRI restriction sites. (B) Inverted repeat constructs are generated by ligating HindIII (or BamHI for RNAi constructs 2 and 5) and NotI digested PCR fragment and HindIII (or BamHI for RNAi constructs 2 and 5) and EcoRI digested PCR fragment together with a NotI- and EcoRI-digested 129 bp spacer segment from the Pichia pastoris Aox-1 gene into the HindIII-digested (or BamHI for RNAi constructs 2 and 5) pGreen backbone. The fragments are not drawn to scale.
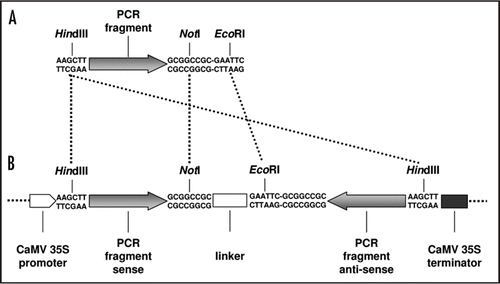
Figure 2 RNAi construct 5 triggers ABA-induced chlorosis. Comparison of the leaf phenotype of six independent transgenic lines containing RNAi construct 5 (a to f) with the parental line Col-0 three days after application of 100 mM abscisic acid (ABA).

Table 1 RNAi constructs to target homologous AtRLP genes
Acknowledgements
B.P.H.J.T. is supported by a Vidi grant of the Research Council for Earth and Life sciences (ALW) of the Netherlands Organization for Scientific Research (NWO). This project was cofinanced by the Centre for BioSystems Genomics (CBSG) which is part of the Netherlands Genomics Initiative/Netherlands Organisation for Scientific Research.
Addendum to:
References
- Jones DA, Jones JDG. The role of leucine-rich repeat proteins in plant defences. Adv Bot Res 1997; 24:89 - 167
- Joosten MHAJ, de Wit PJGM. The tomato-Cladosporium fulvum interaction: a versatile experimental system to study plant-pathogen interactions. Annu Rev Phytopathol 1999; 37:335 - 367
- Kruijt M, de Kock MJD, de Wit PJGM. Receptor-like proteins involved in plant disease resistance. Mol Plant Pathol 2005; 6:85 - 97
- Geisler M, Nadeau J, Sack FD. Oriented asymmetric divisions that generate the stomatal spacing pattern in Arabidopsis are disrupted by the too many mouths mutation. Plant Cell 2000; 12:2075 - 2086
- Jeong S, Trotochaud AE, Clark SE. The Arabidopsis CLAVATA2 gene encodes a receptor-like protein required for the stability of the CLAVATA1 receptor-like kinase. Plant Cell 1999; 11:1925 - 1933
- Nadeau JA, Sack FD. Control of stomatal distribution on the Arabidopsis leaf surface. Science 2002; 296:1697 - 1700
- Taguchi-Shiobara F, Yuan Z, Hake S, Jackson D. The fasciated ear2 gene encodes a leucine- rich repeat receptor-like protein that regulates shoot meristem proliferation in maize. Genes Dev 2001; 15:2755 - 2766
- Fradin EF, Thomma BPHJ. Physiology and molecular aspects of Verticillium wilt diseases caused by V. dahliae and V. albo-atrum. Mol Plant Pathol 2006; 7:71 - 86
- Kawchuk LM, Hachey J, Lynch DR, Kulcsar F, van Rooijen G, Waterer DR, Robertson A, Kokko E, Byers R, Howard RJ, Fischer R, Prüfer D. Tomato Ve disease resistance genes encode cell surface-like receptors. Proc Natl Acad Sci USA 2001; 98:6511 - 6515
- Thomma BPHJ, van Esse HP, Crous PW, de Wit PJGM. Cladosporium fulvum (syn. Passalora fulva), a highly specialized plant pathogen as a model for functional studies on plant pathogenic Mycosphaerellaceae. Mol Plant Pathol 2005; 6:379 - 393
- Ron M, Avni A. The receptor for the fungal elicitor ethylene-inducing xylanase is a member of a resistance-like gene family in tomato. Plant Cell 2004; 16:1604 - 1615
- Malnoy M, Xu M, Borejsza-Wysocka E, Korban SS, Aldwinckle HS. Two receptor-like genes, Vfa1 and Vfa2, confer resistance to the fungal pathogen Venturia inaequalis inciting apple scab disease. Mol Plant Microbe Interact 2008; 21:448 - 458
- Ramonell K, Berrocal-Lobo M, Koh S, Wan JR, Edwards H, Stacey G, Somerville S. Loss-of-function mutations in chitin responsive genes show increased susceptibility to the powdery mildew pathogen Erysiphe cichoracearum. Plant Physiol 2005; 138:1027 - 1036
- Wang G, Ellendorff U, Kemp B, Mansfield JW, Forsyth A, Mitchell K, Bastas K, Liu CM, Woods-Tör A, Zipfel C, de Wit PJGM, Jones JDG, Tör M, Thomma BPHJ. A genome-wide functional investigation into the roles of receptor-like proteins in Arabidopsis. Plant Physiol 2008; 147:503 - 517
- Altschul SF, Madden TL, Schaffer AA, Zhang JH, Zhang Z, Miller W, Lipman DJ. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 1997; 25:3389 - 3402
- Hellens RP, Edwards EA, Leyland NR, Bean S, Mullineaux PM. pGreen: a versatile and flexible binary Ti vector for Agrobacterium-mediated plant transformation. Plant Mol Biol 2000; 42:819 - 832
- Clough SJ, Bent AF. Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J 1998; 16:735 - 743
- Thomma BPHJ, Penninckx IAMA, Broekaert WF, Cammue BPA. The complexity of disease signaling in Arabidopsis. Curr Opin Immunol 2001; 13:63 - 68
- Fritz-Laylin LK, Krishnamurthy N, Tör M, Sjölander KV, Jones JDG. Phylogenomic analysis of the receptor-like proteins of rice and Arabidopsis. Plant Physiol 2005; 138:611 - 623
- Leister D. Tandem and segmental gene duplication and recombination in the evolution of plant disease resistance genes. Trends Genet 2004; 20:116 - 122
- Stout MJ, Thaler JS, Thomma BPHJ. Plant-mediated interactions between pathogenic microorganisms and herbivorous arthropods. Annu Rev Entomol 2006; 51:663 - 689
- Gabriels SHEJ, Takken FLW, Vossen JH, de Jong CF, Liu Q, Turk SCHJ, Wachowski LK, Peters J, Witsenboer HMA, de Wit PJGM, Joosten MHAJ. cDNA-AFLP combined with functional analysis reveals novel genes involved in the hypersensitive response. Mol Plant Microbe Interact 2006; 19:567 - 576