Abstract
Semagenesis, the process of signal generation, is a novel signaling strategy first uncovered within the parasitic plants. Recent evidence suggests that the parasite's production of reactive oxygen species (ROS) has been focused externally to exploit the host's innate immunity. Here we use the inducer identified from decoding semagenesis, as well as other signaling strategies of the parasitic plants, to synchronize host commitments of Striga asiatica and reveal the molecular events that control plant development.
“Tell me what you eat and I will tell you who you are.”
J.A. Brillat-Savarin
All organisms maintain regulatory processes that enable appropriate responses to Nature's constant barrage of biotic and abiotic stimuli. These responses are particularly notable in plants where a sessile life strategy has necessitated developmental programs and regulatory processes intimately coupled with the external environment. Indeed, the successful defense against pathogens, reproductive cycles, cell growth, tissue development and many other processes are under the complex and often overlapping control of both exogenous and endogenous signals. Included are a diverse array of unique secondary metabolites, fragments of structural biopolymers, hormones and even osmotic pressure, temperature, light and pH. Notable among the plants are the obligate parasitic angiosperms where host derived xenognosins direct much of their developmental programs ().Citation1–Citation4 As these parasites are responsible for billions in lost agricultural yields worldwide and a major source of malnutrition and poverty throughout the developing world, there has been considerable interest in developing new strategies to interpret and disrupt signaling. The recent elucidation of a new role for reactive oxygen species (ROS)—molecules traditionally associated with defense—in host selection by parasitic angiosperms underscores the complex interplay between signaling events, and may open new strategies for intervention.Citation5
This new process, known as semagenesis for signal generation, was discovered in the parasitic Scrophulariaceae Striga asiatica. Previous results had suggested that the accumulation of the p-benzoquinone xenognosins, which regulate the transition between vegetative growth and development of the host attachment organ known as the haustorium, were dependant on parasite-derived H2O2.Citation1–Citation4 In the recent study, fluorescence and transmission electron microscopies successfully defines both the cellular site of oxidant production as well as the sensitive feedback regulation by the xenognosin.Citation5 Indeed, ROS production by the parasite appeared insufficient to cross-link cell wall components or activate apoptosis in either plant, but was sufficient to generate the necessary p-benzoquinone xenognosins from host cell walls. Such precise control over ROS accumulation helps explain both the earlier reports of xenognosin signal integration by the parasite,Citation6 as well as how complications associated with using ROS as a secondary messenger in defense responses can be avoided during pathogen attack. It is this last event, where Striga effectively uses the first line of the eukaryotic cell's defense against pathogens—the production of ROS—to effectively “taste” the surface of host cells that may provide the most interesting insight into the evolutionary origins of semagenesis. The activity of such simple cell wall oxidation products as haustoria inducers generally among the parasitic plants suggests that semagenesis is a broadly distributed process.Citation4,Citation7
In addition to broadening the roles for ROS in biology and contributing to our understanding of the mechanisms for host detection, or xenognosis, the observed regulation of oxidant production and xenognosin generation opens the door to a number of research questions uniquely accessible through parasitic plants. For example, with the better mechanistic understanding of signal generation and the clear time dependence for commitment has emerged the very real possibility of constructing differential expression libraries during the developmental transitions. As shown in , the entire seedling population can be synchronized by their dependence on a single xenognosin, thus eliminating the inherent random ‘noise’ of most developmental commitments. Indeed, initial libraries have now been constructed for haustorial development and used in identifying the Striga NADPH oxidase 1 (SaNOX1) as the oxidant source in semagenesis.Citation8
In fact the initial developmental transitions shown in , specifically germination, haustorial initiation, and shoot apical meristem development can now be synchronized. Libraries for each of the xenognosin-mediated transitions would provide spatial and temporal maps of the necessary genes. Indeed, building on the success of the initial library for haustorial development, a new library for the initiation of shoot apical meristem development was constructed. Initial results have identified additional components implicated in ROS regulation, unexpected genes for cell wall synthesis and expansion, and novel regulators of cellular development. Such a molecular map offers a unique window onto the elements regulating the transitions into and out of meristem quiescence. Not only is this information of foundational importance to plant development and evolution, but specific targets for pathogenic inhibition may emerge from the study of these agriculturally significant parasites.
Figures and Tables
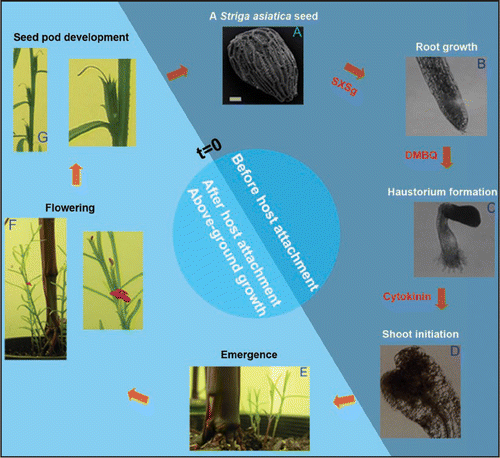
Figure 1 Life cycle of Striga asiatica. The life cycle of the parasitic angiosperm Striga asiatica is divided between early developmental transitions with known xenognosins (dark blue) and latter stages where the necessity of such signals is less clear (light blue). (A) Striga asiatica seed. The tiny seed, shown here by SEM, contains sufficient nutrients to sustain growth for only 5 days post-germination (Bar = 31.2 µm). To enhance survival, germination is coupled to the presence of xenognosins specific for germination, like the hydroquinone SXSg derived from the root exudate of Sorghum spp.Citation9 (B) Vegetative growth. Upon germination, the growing seedling generates H2O2 to oxidize host cell wall phenols and release haustorial-inducing p-benzoquinones.Citation10 These xenognostic signals diffuse back to the parasite and initiate development of the haustorium. (C) Haustorium development. Host attachment occurs via a specialized organ known as the haustorium. Once formed, the organ penetrates the host tissues, ultimately forming a vascular connection for parasitism. (D) Shoot apical meristem (SAM) development. Penetration of the host root precedes the initiation of shoot growth. In vitro, SAM initiation can be induced by cytokinins. (E–G). Emergence/Flowering/Seeds development on Zea mays. Striga asiatica seedlings emerge 8 weeks after inoculation on host roots and the mature plant develops closed flowers that give rise to seed pods containing approximately 500 seeds each.
Addendum to:
References
- Kuijt J. The Biology of Parasitic Flowering Plants 1969; Berkeley University of California Press
- Press MC, Graves JD. Haustorial initiation and differentiation in Parasitic plants 1995; London Chapman & Hall 39 - 79
- Kim D, et al. On becoming a parasite: evaluating the role of wall oxidases in parasitic plant development. Chem Biol 1998; 5:103 - 117
- Palmer A, et al. Chemical biology of multi-host/pathogen interactions:chemical perception and metabolic complementation. Annu Rev Phytopath 2004; 42:439 - 464
- Keyes WJ, et al. Semagenesis and the parasitic angiosperm Striga asiatica. Plant J 2007; 51:707 - 716
- Smith C, Dudley M, Lynn D. Vegetative/parasitic transition: control and plasticity in striga development. Plant Physiol 1990; 93:208 - 215
- Yoder J. Host-plant recognition by parasitic Scrophulariaceae. Curr Opin Plant Biol 2001; 4:359 - 365
- Liang Lizhi, et al. Feed-back regulation of NADPH oxidases and the control of semagenesis in Striga asiatica. Submitted to Plant Physiol 2008;
- Chang M, et al. Chemical regulation of distance. Characterization of the first natural host germination stimulant for Striga asiatica. J Am Chem Soc 1986; 108:7858 - 7860
- Chang M, Lynn DG. The haustorium and the chemistry of host recognition in parasitic angiosperms. J Chem Ecol 1986; 12:561 - 579