Abstract
Several different D-type cyclins can be found in plants and in maize, four of these have been characterized: CycD2;1, CycD4;1, CycD5;1 and CycD5;2. These cyclins appear to form complexes with Cdks, with PCNA and also with KRP proteins and in these kinase activity can be measured. The expression of the corresponding genes during maize germination is highly stimulated by phytohormones like auxin and cytokinin, however this is not followed by an equivalent increase in the amount of the corresponding proteins; nonetheless, auxins do stimulate the associated kinase activity, particularly at early germination times. Thus, auxins appear to stimulate the cell cycle during germination at two levels, transcription and kinase activation. Both auxins and cytokinins appear to shorten the G1 phase during germination and stimulate DNA synthesis, but apparently they do it in different ways as the simultaneous addition of both to germinating maize axes eliminates DNA synthesis stimulation. Therefore, similar actions may be achieved by different paths.
Cyclins D and Association to G1 Cell Cycle Proteins
During the G1 phase of the cell cycle in mammalian cells, cyclin D (D1–D3) proteins associate to Cdk proteins (either Cdk 4 or 6) forming a complex with kinase activity; this complex is often found associated with so called kinase inhibitors, p21Cip1 or p27 Kip1,Citation1 which seem also to serve as vehicles to carry cyclin-kinases into the nucleus. The protein PCNA (Proliferating Cell Nuclear Antigen) is also part of these complexes. The latter, originally described as a sliding clamp for replicative DNA polymerase δ, appears to function as a meeting point for multiple DNA metabolism enzymes (replication, repair, recombination) and regulatory factors such as cyclins/Cdks.Citation2 Similar proteins appear to be present in plant cells and our studies have shown that PCNA forms complexes with D cyclins and Cdks in an analogous manner to animal cells.Citation3 p27kip1-type proteins have been described in plant cells, known as KRPs for Kip1 related proteins and they have been shown associating to Cdks or to D-type cyclins.Citation4–Citation6 Association of maize cyclins D5 and D4;1 with a maize KRP1 type protein has also been observed (). Thus, a similar quaternary complex formed by D cyclins, Cdks, PCNA and KRP proteins might function in plant cells.
Of interest, and differently to animal cells, multiple D cyclins are found in plant cells, the physiological meaning of which is not yet understood. In the case of maize cells, at least 4 D-cyclins have been described (there are 12 in riceCitation7) and our studies indicate that each one of them is expressed at different times during germination; moreover, they are differentially regulated by phytohormones, particularly auxins in germinating maize seeds.Citation8 Although similar results were described for cyclins D in Arabidopsis cells in culture, a distinction is observed in the way cyclins D belonging to the same classification group respond to phytohormones and/or to sugars: maize cyclin D2;1 responds differently to stimuli to Arabidopsis cycD2;1, but similarly to Arabidopsis D3;1.Citation9 It is necessary to be cautious though since the models of study are different.
Cyclins D, Phytoregulators and Maize Germination
Seed germination would represent an excellent model to study reactivation of proliferation as most if not all cells are halted in a G1 DNA state (particularly those cells in meristems) in the dry seed. Water imbibition gradually activates metabolism and the cell cycle; however, cells in maize dry seed axes already contain visible levels of every cell cycle protein measured: cyclins, Cdks, PCNA, DNA polymerases, DNA ligase and others. For some of them, their levels do not change during the germination time (DNA polymerases and Cdks), although changes in their activities are observed.Citation10 Others like D cyclins show significant, and cyclin-specific, fluctuations during germination, with only a partial correlation with the associated kinase activity.Citation8,Citation11 This behaviour does not match with the typical onset of a cell cycle in which protein factors and cell cycle events should gradually start appearing. In fact, activation of pre-existing enzymes during germination appears to be the way the cell cycle is started. In this context, progression of the cell cycle during germination of a seed may be atypical.
The onset of the S phase correlates well with an important increase in the activity of DNA polymerases α and δCitation12,Citation13 and DNA synthesis during germination is greatly enhanced if seeds are imbibed in the presence of auxins (IAA) or cytokinins (BA). Thus, there must be a signaling cascade that is activated at some point during the germination process that has existing replicative DNA polymerases as targets. Perhaps the endogenous phytohormones participate, either directly or indirectly, in the mechanism that regulates the S phase onset. Thus, it is relevant that both cytokinins and auxins, added exogenously, accelerate entry into the S phase during the germination process and stimulate an early increase in the kinase activity associated to Cyclins D5 and D4;1, even though neither phytoregulator increases the amount of cyclins proteins present in germinating seeds.Citation11 This shortening of the cell cycle timing might be part of the mechanism by which these phytohormones stimulate the S phase.Citation10
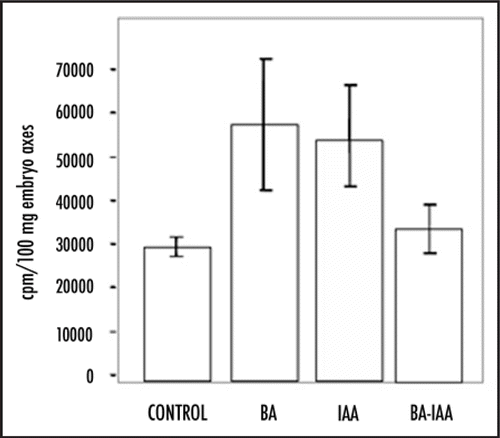
Although the mechanism by which BA and IAA stimulate DNA synthesis is not known, it may not be exactly the same. Addition of both phytohormones together to germinating seed axes reduces or eliminates the individual effect, i.e., there is almost no stimulation of DNA synthesis (). The reported effect that one phytohormone has against the accumulation of the otherCitation14 may not be the only reason for this behaviour as both IAA and BA are added exogenously at similar concentrations. Therefore, the biochemical perception of every phytoregulator by cells, leading to cell cycle stimulation, may follow different pathways. Examples are stimulation of PCNA protein accumulation by cytokinins but not auxins (our unpublished results), and strong stimulation of cyclin D5;2 expression by auxins, but only slightly by cytokinins, during early germination.Citation15,Citation8 Similar physiological responses may be achieved by different paths.
Figures and Tables
Figure 1 Association of maize D cyclins with maize KRP1. Protein extracts from 12 h imbibed maize embryo axes were immunoprecipitated using anticyclin D4;1 antibody (A) or anti-Cyclin D5 antibody (B) and immunoprecipitates were prepared for western blotting, using an anti-maize KRP1 antibody. Lane 1, KRP1 antibody alone; lane 2, antibody + maize protein extract; lane 3, maize protein extract.
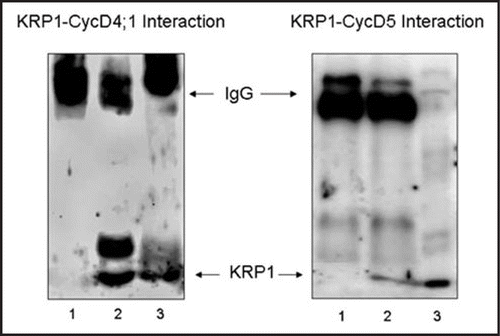
Figure 2 DNA synthesis stimulation by BA and IAA in germinating seeds. Maize embryo axes were incubated in imbibition buffer containing (methyl-3H) thymidine for 15 h in the presence of BA (10−6 M), IAA (10−6 M) or both and incorporation of the radioactive precursor was taken as the capacity of cells in embryo axes to synthesize DNA. DNA replication starts by 12 h of imbibition during maize germination.Citation16
Acknowledgements
We acknowledge receiving grants PAPIIT IN-206799 and IN-200706 and Conacyt grant 42896-Z.
Addendum to:
References
- Xiong Y, Zhang H, Beach D. D-type cyclins associate with multiple protein kinases and the replication and repair factor PCNA. Cell 1992; 71:505 - 514
- Kelman Z. PCNA: structure, functions and interactions. Oncogene 1997; 14:629 - 640
- Sánchez MP, Torres A, Boniotti AB, Gutierrez C, Vázquez Ramos JM. PCNA protein associates to Cdk-A type protein kinases in germinating maize. Plant Mol Biol 2002; 50:167 - 175
- De Veylder L, Beeckman T, Beemster GT, Krols L, Terras F, Landrieu I, Van der Schueren E, Maes S, Naudts M, Inzé D. Functional analysis of cyclin-dependent kinase inhibitors of Arabidopsis. Plant Cell 2001; 13:1653 - 1667
- Weinl C, Marquart S, Kuijt SJH, Nowack MK, Jakoby MJ, Hülskamp M, Schnittger A. Novel funtions of plant cyclin-dependet kinase inhibitors, ICK1/KRP1, can act non-cell-autonomously and inhibit entry into mitosis. Plant Cell 2005; 17:1704 - 1722
- Verkest A, Weinl C, Inzé D, De Veylde L, Schnittger A. Switching the cell cycle. Kip-Related Proteins in plant cell cycle control. Plant Physiol 2005; 139:1099 - 1106
- Guo J, Song J, Wang F, Zhang XS. Genome-wide identification and expression analysis of rice cell cycle genes. Plant Mol Biol 2007; 64:349 - 360
- Quiroz Figueroa F, Vázquez Ramos JM. Expression of maize D-type cyclins: comparison, regulation by phytohormones during seed germination and description of a new D cyclin. Physiol Plant 2006; 128:556 - 568
- Gutiérrez R, Quiroz Figueroa F, Vázquez Ramos JM. Maize cyclin D2 expression, associated kinase activity and effect of phytohormones during germination. Plant Cell Physiol 2005; 46:166 - 173
- Vázquez Ramos JM, Sánchez MP. The cell cycle and seed germination. Seed Sci Res 2003; 13:113 - 130
- Lara Núñez A, de Jesús N, Vázquez Ramos JM. Maize D4;1 and D5 cyclin proteins in germinating maize. Associated kinase activity and regulation by phytohormones. Physiol Plant 2008; 132:79 - 88
- Coello P, Vázquez Ramos JM. Maize DNA polymerase 2 is a phosphoprotein with increasing activity during germination. Eur J Biochem 1995; 231:99 - 103
- García E, Quiroz F, Uchiyama Y, Sakaguchi K, Vázquez Ramos JM. Expression of a maize δ-type DNA polymerase during seed germination. Physiol Plant 2006; 127:268 - 276
- Crozier A, Kamiya Y, Bishop G, Yokota T. Buchanan, Gruissem, Jones. Biosynthesis of hormones and elicitor molecules. Biochemistry and Molecular Biology of plants 2000; First edition 850 - 929
- Herrera I, Sánchez MP, Molina J, Plasencia J, Vázquez Ramos JM. Proliferating Cell Nuclear Antigen expression in maize seed development and germination. Regulation by phytohormones and its association with cell cycle proteins. Physiol Plant 2000; 110:127 - 134
- Baiza AM, Vázquez Ramos JM, Sánchez De Jiménez E. DNA synthesis and cell division in embryonic maize tissues during germination. J Plant Physiol 1989; 135:416 - 421